
Allergy-Protective Arabinogalactan Modulates Human Dendritic ...
11
of February 20, 2018. This information is current as B κ C-Type Lectins and Inhibition of NF- Modulates Human Dendritic Cells via Allergy-Protective Arabinogalactan and Albrecht Bufe A. Megger, Barbara Sitek, Birgit Classen, Esther M. Heise Marcus Peters, Patrick M. Guidato, Karin Peters, Dominik http://www.jimmunol.org/content/196/4/1626 doi: 10.4049/jimmunol.1502178 January 2016; 2016; 196:1626-1635; Prepublished online 8 J Immunol Material Supplementary 8.DCSupplemental http://www.jimmunol.org/content/suppl/2016/01/07/jimmunol.150217 References http://www.jimmunol.org/content/196/4/1626.full#ref-list-1 , 16 of which you can access for free at: cites 46 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2016 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on February 20, 2018 http://www.jimmunol.org/ Downloaded from by guest on February 20, 2018 http://www.jimmunol.org/ Downloaded from
Transcript of Allergy-Protective Arabinogalactan Modulates Human Dendritic ...

of February 20, 2018.This information is current as
BκC-Type Lectins and Inhibition of NF-Modulates Human Dendritic Cells via Allergy-Protective Arabinogalactan
and Albrecht BufeA. Megger, Barbara Sitek, Birgit Classen, Esther M. Heise Marcus Peters, Patrick M. Guidato, Karin Peters, Dominik
http://www.jimmunol.org/content/196/4/1626doi: 10.4049/jimmunol.1502178January 2016;
2016; 196:1626-1635; Prepublished online 8J Immunol
MaterialSupplementary
8.DCSupplementalhttp://www.jimmunol.org/content/suppl/2016/01/07/jimmunol.150217
Referenceshttp://www.jimmunol.org/content/196/4/1626.full#ref-list-1
, 16 of which you can access for free at: cites 46 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2016 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on February 20, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on February 20, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Allergy-Protective Arabinogalactan Modulates HumanDendritic Cells via C-Type Lectins and Inhibition of NF-kB
Marcus Peters,*,1 Patrick M. Guidato,*,1 Karin Peters,* Dominik A. Megger,†
Barbara Sitek,† Birgit Classen,‡ Esther M. Heise,‡ and Albrecht Bufe*
Arabinogalactan (AG) isolated from dust of a traditional farm prevents disease in murine models of allergy. However, it is unclear
whether this polysaccharide has immune regulatory properties in humans. The aim of this study was to test the influence of AG on
the immune-stimulating properties of human dendritic cells (DCs). Moreover, we sought to identify the receptor to which AG binds.
AG was produced from plant callus tissue under sterile conditions to avoid the influence of pathogen-associated molecular patterns
in subsequent experiments. The influence of AG on the human immune system was investigated by analyzing its impact on
monocyte-derived DCs. To analyze whether the T cell stimulatory capacity of AG-stimulated DCs is altered, an MLR with naive
Th cells was performed. We revealed that AG reduced T cell proliferation in a human MLR. In the search for a molecular mech-
anism, we found that AG binds to the immune modulatory receptors DC-specific ICAM-3–grabbing non integrin (DC-SIGN) and
macrophage mannose receptor 1 (MMR-1). Stimulation of these receptors with AG simultaneously with TLR4 stimulation with
LPS increased the expression of the E3 ubiquitin-protein ligase tripartite motif–containing protein 21 and decreased the phos-
phorylation of NF-kB p65 in DCs. This led to a reduced activation profile with reduced costimulatory molecules and proinflam-
matory cytokine production. Blocking of MMR-1 or DC-SIGN with neutralizing Abs partially inhibits this effect. We conclude
that AG dampens the activation of human DCs by LPS via binding to DC-SIGN and MMR-1, leading to attenuated TLR signaling.
This results in a reduced T cell activation capacity of DCs. The Journal of Immunology, 2016, 196: 1626–1635.
The impact of allergic diseases is rising unimpeded in in-dustrial countries. The hygiene hypothesis proposes acausal explanation for this phenomenon, namely a re-
duction in infectious disease in early childhood (1). Although atempting explanation, epidemiological studies deliver inconsistentresults (2–4). In the search for other conditions that are protectiveagainst allergic disease, recent studies have shown that growing upand living in rural areas decrease the risk of developing allergiessignificantly compared with living in cities (5). In particular,children, when they were exposed to a traditional farming envi-ronment in the rural areas of the Alps, were protected in this way.Further studies showed that contact with barns in the earliestchildhood resulted in allergy-protective effects (6–9).
These allergy-protective effects have been traced back to dif-ferent factors, such as the inhalation of cowshed dust extracts(CDE) or single components isolated from CDE (10–13).One allergy-protective molecule that makes up 13% of CDE total
mass was identified as arabinogalactan (AG) (14). It was shown ina mouse asthma model that AG isolated from Alopecurus pra-tensis can prevent allergic airway inflammation and sensitization.AGs are polysaccharides that are ubiquitously present in mostplants and even in mycobacteria, but not in animals. They consistpredominantly of galactose and arabinose. Further sugars, such asrhamnose, fucose, and mannose, occur to a lesser extent. DifferentAG preparations are already in use in several traditional andnatural medical products worldwide and are subject to studies dueto their immune-modulating properties (15–17).Polysaccharides mediate their immune-modulatory properties in
mammals predominantly via binding to carbohydrate receptors.Dendritic cells (DCs) express a wide pattern of these receptors witha variety of different functions. Among these receptors, the group ofC-type lectin receptors (CLRs) is the largest one. The DC-specificICAM-3–grabbing nonintegrin (DC-SIGN) and the macrophagemannose receptor 1 (MMR-1) are two commonly expressed CLRson DCs. Both receptors are known to interact with mannose andfucose, but they differ in their number of carbohydrate recognitiondomains and cellular signaling. Several studies have identifiedexogenous ligands binding to DC-SIGN, such as the HIV, measlesvirus, Mycobacterium tuberculosis, and other pathogens. Fur-thermore, many pathogens use DC-SIGN not only to enter the DC,but also to inhibit DC function (18, 19). Unlike the members ofthe group of TLRs, these CLRs induce cellular activation, only toa much lesser extent. However, stimulation of these receptors,such as DC-SIGN, may modulate TLR-induced cytokine secretionand DC maturation.We have previously shown that AG isolated from CDE, hay, or
different grasses is able to protect from allergic disease in a mouse
*Department of Experimental Pneumology, Ruhr University Bochum, 44801 Bochum,Germany; †Medical Proteome Center, Ruhr University Bochum, 44801 Bochum,Germany; and ‡Department of Pharmaceutical Biology, Christian Albrechts Univer-sity, 24118 Kiel, Germany
1M.P. and P.M.G. contributed equally to this work.
ORCIDs: 0000-0003-1850-2845 (M.P.); 0000-0003-4737-2293 (P.M.G.); 0000-0002-1058-7744 (D.A.M.); 0000-0003-1452-801X (B.C.).
Received for publication October 7, 2015. Accepted for publication December 8,2015.
This work was supported by Deutsche Forschungsgemeinschaft Grant PE 1813/2-1,intramural research funding from Ruhr University Bochum Grants F659-2009 andF797-2013, and by funds from Protectimmun GmbH.
Address correspondence and reprint requests to Dr. Marcus Peters, Zentrum f€urKlinische Forschung II, Universitatsstraße 150, 44801 Bochum, Germany. E-mailaddress: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: AG, arabinogalactan; CDE, cowshed dust extract;CLR, C-type lectin receptor; DC, dendritic cell; DC-SIGN, DC–specific ICAM-3–grabbing nonintegrin; IL-1RA, IL-1R antagonist; MMR-1, macrophage mannosereceptor 1; moDC, monocyte-derived DC; PAMP, pathogen-associated molecularpattern; rh, recombinant human; TRIM21, tripartite motif–containing protein 21.
Copyright� 2016 by TheAmericanAssociation of Immunologists, Inc. 0022-1767/16/$30.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1502178
by guest on February 20, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

model of asthma. Therefore, it is of great interest whether AG isalso able to modulate the human immune response. Additionally,mice, compared with humans, have five different DC-SIGN ho-mologs named SIGNR1–SIGNR5, but there are contrary findingsas to which of them is the functional homolog to human DC-SIGN(20–22). In fact, these homologs have neither the same ligandspecificity nor the same signaling pathways compared with humanDC-SIGN. Besides DC-SIGN, there are even more structural andfunctional differences in CLRs between mice and humans (23).Therefore, we chose to investigate the effect of AG in a humanmodel of T cell activation to get direct insight into the modulationof the human immune system by AG.One major problem concerning in vitro studies with sub-
stances isolated from plants is the contamination with pathogen-associated molecular patterns (PAMPs). In particular, whenworking with cells of the innate immune system, tiny amounts ofPAMPs can have a major influence on the results of the ex-periments. Therefore, we have focused in the present study onthe production of contamination-free AG from plant cell tissuecultures. We aimed to investigate whether this PAMP-free AGmodifies the immune-stimulatory capacity of human monocyte-derived DCs (moDCs). Moreover, we sought to identify re-ceptors to which AG bind and to get insights into the molecularmechanisms behind immune modulation.
Materials and MethodsReagents and Abs
For flow cytometric analysis, anti–CD1a-PE (clone HI149; Immunotools,Friesoythe, Germany), anti-CD14 (clone TLK4; Miltenyi Biotec, BergischGladbach, Germany), anti-CD209 (clone DCN47.5; Miltenyi Biotec), andanti-CD83 (clone HB15a; Beckman Coulter, Krefeld, Germany) wereused. Anti-human phospho–NF-kB p65 (S529) PE (clone B33B4WP) andCFSE were purchased from eBioscience (San Diego, CA). Recombinanthuman (rh)DC-SIGN–Fc chimera and rhMMR-1 were purchased fromR&D Systems (Minneapolis, MN). Anti-human IgG1-biotin Ab andsandwich ELISA for human IFN-g, IL-6, IL-12p70, IL-10, and TNF-awere purchased from BD Biosciences (Erembodegem, Belgium).Blocking Abs against DC-SIGN (clone DCN47.5) and human MMR(clone 15-2) as well as an murine IgG1 isotype control Ab (clone T8E5)were purchased from InvivoGen (Toulouse, France). The concentrationof Ab used for blocking experiments was 10 mg/ml. These Abs werecarefully selected to not take influence on the activation of cells by LPS(Supplemental Fig. 1). Anti–His-biotin Ab was purchased from Qiagen(Hilden, Germany). The b-D-glucosyl Yariv reagent [1,3,5-tris-(4-b-glucopyranosyl-oxyphenylazo)-2,4,6-trihydroxybenzene] was preparedaccording to Yariv et al. (24). p-Aminophenyl-b-D-glucoside (1.8 mM) wasdiazotized with sodium nitrite (1.8 mM) in 10.8 ml 0.5 M HCl at 0˚C andadded dropwise to 0.45 mM phloroglucin dissolved in 22.5 ml water, withthe pH being kept at 9 by addition of 0.5 M NaOH. After 2 h the pH becamestable and an equal volume of methanol was added. The precipitate wascollected by centrifugation (4000 3 g) and dissolved in 40 ml water.The precipitation step was done again and the residue was dried at 40˚C.The Yariv reagent was stored frozen at 220˚C as a 1 mg/ml solutionin 0.9% sodium chloride. rhGM-CSF and rhIL-4 were purchased fromImmunotools.
Generation, cultivation, and purification of AG from Phleumpratense callus tissue suspension culture
The surface of P. pratense (Poaceae) seeds were sterilized by treatment for10 min in 5% thymol solubilized in 96% ethanol, followed by 10 min in2% sodium hypochloride. After washing with sterile water, the seeds weregrown on plates prepared with 1% agarose in Murashige–Skoog mediumcontaining sucrose and the following phytohormones: 2 mg/ml 2,4-dichlorophenoxyacetic acid, 4 mg/ml a-naphthaleneacetic acid, and 0.3mg/ml kinetin (all purchased from Sigma-Aldrich, St. Louis, MO). Aftergermination at 27˚C in darkness, parts of the shoot were cut and transferredto fresh agar plates to generate callus tissues. After 2 wk of growing, callustissue was transferred to Murashige–Skoog liquid medium containingthe same phytohormone concentrations as described above for the agar
plates. Suspension culture was performed in the absence of antibiotics.Suspension-cultured cells were then incubated at 27˚C at 120 U/min in ahorizontal shaker. Supernatants were collected after centrifugation (10min, 3000 3 g).
Supernatants of suspension cultures were then pooled and autoclaved. Inthis step proteins and peptides were denaturized so that the mucus-likestructure was resolved. To remove insoluble components, three filtrationsteps were included. At first, pooled and autoclaved callus supernatantswere filtered with a paper filter, afterward a glass fiber filter, and at last a0.22-mm cellulose filter was used. The raw AG-containing solution wasconcentrated by using a 30-kDa filtration unit (Merck Millipore, Billerica,MA). AG concentration was determined by b-D-glucosyl Yariv radialdiffusion gel as described previously (14). For precipitation of AG, anappropriate amount of Yariv’s reagent was added to the raw AG-containingsolution. After centrifugation for 1 h at 21,000 3 g, sediment was washedonce with sterile 0.9% NaCl solution. Afterward, 1 ml saturated sodiumdithionite solution was added to the sediment and incubated at 60˚C untilthe red color disappeared completely. AG was then purified by size ex-clusion chromatography on a PD-10 column (GE Healthcare, Fairfield,CT) and eluted in 1-ml fractions. AG-containing fractions were identifiedby radial diffusion assay and pooled. Concentration was then determinedby a resorcin assay and radial diffusion gel with AG from gum arabic(Biosupplies Australia, Melbourne, VIC, Australia) as a standard. Thesubstance was proved to be LPS free by TLR4 assay as described previ-ously (25). Molecular mass of the preparation was determined by sizeexclusion chromatography as described by Gollner et al. with followingmodifications: column set (two PL aquagel-OH 40, 8 mm, 300 3 7.5 mmand one PL aquagel-OH mixed, 8 mm, 300 3 7.5 mm) and flow rate(0.7 ml/min) (26).
Membrane receptor precipitation assay
Membranes of human moDCs were prepared as follows. Briefly, DCs werewashed with ice-cold PBS and resuspended in 20 mM HEPES buffer with250 mM sucrose, 1% digitonin, and protease inhibitor mixture (Roche,Basel, Switzerland). Cells (2.4 3 107) were mechanically homogenized andcentrifuged for 7 min at 700 3 g and 4˚C. Membrane-containing super-natant was collected and membranes were sedimented by centrifugationfor 60 min at 16,5003 g and 4˚C and washed twice. Membranes were thenresuspended in 1 ml cold HBSS containing protease inhibitor mixture(Roche, Basel, Switzerland). Resuspended membranes (100 ml) with aprotein concentration of 1.6 mg/ml were then incubated with 100 mg AGfor 1 h at 4˚C in HBSS buffer to allow receptors to bind AG. Afterward,AG-receptor complexes were precipitated by adding b-D-glucosyl Yariv’sreagent. Precipitates were washed twice with ice-cold HBSS, and after-ward reducing SDS-PAGE loading buffer (27) was added. Sample bandsfrom the SDS-PAGE were further analyzed by mass spectrometry.
Proteome analysis
For proteomics experiments, liquid chromatography–tandem mass spec-trometry analyses of tryptically digested proteins were carried out on anUltiMate 3000R SLC nano system online coupled to an Orbitrap Eliteinstrument (both Thermo Scientific, Bremen, Germany). Separated gelbands obtained after membrane receptor precipitation assay were digestedin gel with trypsin. Samples of human moDCs were digested in solutionafter lysis and protein solubilization with RapiGest (Waters, Eschborn,Germany) following the manufacturer’s instructions.
For protein identification, Proteome Discoverer software (ver. 1.3,Thermo Scientific, Waltham, MA) was used to search against UnipPotKB/Swiss-Prot database with Mascot search algorithm. Taxonomy was re-stricted to humans. Peptide modifications were considered at methionine(oxidation [variable]) and cysteine (propionamide for in-gel digestion[variable] and carbamidomethyl for in-solution digestion [fixed]). Precursormass tolerance was set to 5 ppm, fragment mass tolerance to 0.4 Da, andmaximum number of missed cleavage sites to 1. Peptide identificationswith .1% false discovery rate were discarded. Quantitative analyses wereperformed with Progenesis liquid chromatography–tandem mass spec-trometry software (ver. 4.0.4265.42984, Nonlinear Dynamics, Newcastleupon Tyne, U.K.). A detailed description of the single steps of quantifi-cation has been recently published (28, 29).
Inhibitory ELISA
To confirm the binding between AG and the identified receptors, an in-hibitory ELISA was used. Briefly mannan from Saccharomyces cerevisiaewas coated to 96-well ELISA plates (Thermo Fisher Scientific, Waltham,MA). The receptor of interest (DC-SIGN or MMR-1) was preincubatedeither with dilutions of mannan or AG for 1 h in HBSS buffer at 37˚C.
The Journal of Immunology 1627
by guest on February 20, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Afterward, the preincubation mixtures were transferred to mannan-coatedplates and incubated for 1 h at room temperature while shaking. Boundreceptors were detected by anti-human IgG1 secondary Ab (DC-SIGN) oranti-His Ab (MMR-1). Detection of biotinylated secondary Ab was carriedout by streptavidin-HRP (Sigma-Aldrich) and OptEIA tetramethylbenzi-dine substrate (BD Biosciences). Absorbance was measured at 450 nm.
Isolation and generation of human moDCs
Monocytes were isolated from a total of 18 different donors (61% female).Voluntary, healthy donors were selected independently of their sensitizationstatus to allergens. DCs were generated from human blood monocytes asdescribed elsewhere (30, 31). On day 6, DCs were positive for the fol-lowing surface molecules CD1a (93 6 1%), DC-SIGN (86 6 3%), CD83(15 6 3%), and CD86 (17 6 1%). All experimental procedures wereapproved by the Ethics Committee at Ruhr University Bochum (Bochum,Germany).
Human moDC costimulation experiments
moDCs were harvested and stimulated with 1 mM AG. After 4 h ofstimulation, 1 ng/ml LPS was added. After 24 h, supernatants were col-lected and cytokines were measured by ELISA. DCs were harvested,stained for surface markers, and analyzed by flow cytometry.
To detect NF-kB subunit p65 and its phosphorylation, an instant ELISAkit of eBioscience (Affymetrix eBioscience, Frankfurt, Germany) was usedaccording to the manufacturer’s protocol. DCs were stimulated as de-scribed above with the difference that activation with LPS was shortened to1 h before lysis of cells. Intracellular staining of phosphorylated p65 wasperformed by staining cells that were fixed with paraformaldehyde fol-lowed by permeabilization with methanol. Cells were stained with 0.03 mganti-human phospho–NF-kB p65-PE. Stained cells were analyzed by flowcytometry (CyFlow SL; Partec, M€unster, Germany).
Quantitative proteome analysis of human moDCs
Human moDCs were harvested and stimulated with AG, LPS, or both for24 h. Afterward, cells were washed, collected, and used for comparativequantitative proteomics analysis. In this study, five different monocytedonors were used to obtain statistically relevant data. Afterward, data wereanalyzed by a “protein information resource” database search for processesassociated to the significantly regulated proteins.
For visualization of the proteomic data, proteomaps were generated asdescribed previously (32).
Human moDC and naive CD4 T cell cocultures
Human naive CD4+ T cells were isolated with a human naive CD4+ iso-lation kit II (Miltenyi Biotec). T cells were stained with CFSE, as de-scribed elsewhere for proliferation assay (10). moDCs and naive CD4+
cells were cocultivated for 6 d at a ratio of 10,000 DCs to 20,000 naiveCD4+ T cells in 96-well round bottom plates. Cocultured cells werestimulated with AG alone or in combination with LPS. After 6 d of co-cultivation, CFSE-stained cocultures were used to determine T cell pro-liferation by flow cytometry, and supernatants were used to measure T cellcytokines (IFN-g, IL-5, IL-13, and IL-17A).
Statistical analysis
All data were analyzed by a Mann–Whitney test, Wilcoxon test, or one-wayANOVA as indicated. GraphPad Prism software (ver. 5; GraphPad Soft-ware, La Jolla, CA) or Progenesis liquid chromatography–mass spec-trometry (Nonlinear USA, Durham, NC) were used for the analysis. Thep values , 0.05 were considered statistically significant. Results are pre-sented as medians or means as indicated.
ResultsAG reduces LPS-induced inflammatory cytokine secretion andcostimulatory surface molecule expression in human moDCs
We isolated AG from P. pratense cell suspension culture. Themolecular mass was determined by multiangle laser light scat-tering to be 127 kDa. AG was tested to be free of TLR2 and TLR4ligands via TLR-transfected HEK293 cells. After confirming theallergy-protective activity of AG in a mouse model of allergicairway inflammation (Supplemental Fig. 2) we aimed to testwhether this molecule may also modulate the behavior of humanDCs in vitro.
Therefore we generated moDCs and stimulated the immaturecells with 1 mM AG, 1 ng/ml LPS, or both for 24 h to investigatethe effects of AG on DC biology. Afterward, we investigated thesurface marker expression by flow cytometry, cytokine secretionby ELISA, and cell viability by MTT assay. We observed that AGdoes not have an influence on CD1a (Fig. 1A), CD14, and DC-SIGN (data not shown) expression, showing that AG does notimpair DC generation. Moreover, we found no influence on DCviability, as measured by the MTT test (data not shown). In con-trast, stimulation with AG significantly reduced the expression ofthe activation marker CD83 when it was given 4 h before LPSstimulation (Fig. 1B), showing that AG attenuates the activation ofDCs by LPS. CD86 expression seems to be suppressed in a similarmanner (data not shown).Stimulation of DCs with AG leads to a significant increase in IL-6
and IL-10, but not TNF-a or IL-12p70 secretion. LPS, in contrast,induces a much higher amount of cytokine secretion in this setting.The costimulation of DCs with AG and LPS shows that AG sig-nificantly reduces LPS-induced IL-6, IL12p70, IL-10, and TNF-a(Fig. 1C–F) secretion. Stimulation of DCs with AG or costimulationwith LPS did not show any influence on the IL-1b secretion (datanot shown).
DC-SIGN and MMR-1 serve as receptors for AG
In search of receptors to which AG binds on DCs, we used theability of b-D-glucosyl Yariv reagent to precipitate AG-receptorcomplexes. Precipitated receptor complexes were fractionated bySDS-PAGE (see Supplemental Fig. 3A) and protein bands wereidentified by mass spectrometry.MMR-1, DC-SIGN, and SIGLEC-1 were identified in three
independent experiments as receptors with a high probability ofbinding AG. We focused our further studies on DC-SIGN andMMR, which are both known to modulate the behavior of DCs (33,34). We used an inhibitory ELISA with AG as the inhibitor tofurther confirm the binding of AG to DC-SIGN and MMR-1. Thebinding of DC-SIGN (Fig. 2A) and MMR-1 (Fig. 2A) to coatedmannan was inhibited dose-dependently by preincubation withAG. Representative binding curves are shown in SupplementalFig. 3B for DC-SIGN and for MMR-1 in Supplemental Fig. 3C.We calculated IC50 values for mannan and AG. Both receptors
showed a comparable IC50 for mannan IC50(DC-SIGN) of 0.28 andIC50(MMR-1) of 0.12. DC-SIGN showed a significantly lower IC50
for AG IC50(DC-SIGN) of 0.01 compared with mannan.To test whether binding of AG to DC-SIGN or MMR-1 explains
the altered activation profile found after stimulation of cells withLPS, we repeated the stimulation experiment in the presence ofreceptor-blocking Abs. Interestingly, blocking of binding of AG toeither MMR-1 or DC-SIGN by neutralizing Abs reconstitutedproduction of IL-6 partially (Fig. 3), showing that both receptorsseem to be involved in immune modulation.
Costimulation of moDCs with AG and LPS leads to increasedtripartite motif–containing protein 21 and IL-1R antagonistexpression but reduced NF-kB activation
To get an overview of the expression of cellular proteins that maybe regulated by AG, a quantitative proteome analysis was per-formed. The stimulation of moDCs with AG, LPS, or both showeda specific pattern of differentially regulated proteins (Fig. 4A,Supplemental Table I) linked to distinct cellular processes (Fig.4B). When DCs were stimulated with AG, these cells showedreduced response to LPS, and therefore we focused our attentionon processes that are involved in the negative regulation of TLRsignaling. One of these proteins is the E3 ubiquitin-protein ligasetripartite motif–containing protein 21 (TRIM21) that is involved
1628 MODULATION OF DCs BY ARABINOGALACTAN
by guest on February 20, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

in negative regulation of the NF-kB pathway. TRIM21 was up-regulated 2.2-fold after costimulation of cells with LPS and AG(Fig. 5A).A second protein that is differentially regulated by stimulation
with AG, LPS, or the combination of both is the IL-1R antagonist(IL-1RA), which is a secreted and immune regulatory protein.Expression of IL-1RA was upregulated after stimulation with AG(2.7-fold) or LPS (9.5-fold) and the combination of both (15.2-fold)(Fig. 5B).
Owing to the influence of AG on TRIM21 and the reducedexpression of proinflammatory cytokines, we measured the ex-pression of NF-kB subunit p65 (Fig. 5C) and its phosphorylationby a semiquantitative ELISA (Fig. 5D). The results reveal that theamount of the p65 subunit in cells that were costimulated with AGand LPS is only half the amount in cells that were stimulated withthe different factors alone. Moreover, the amount of the phos-phorylated p65 is greatly reduced in cells that were costimulatedin comparison with cells that were stimulated with LPS only. The
FIGURE 1. Stimulation of moDCs with AG reduces LPS-induced costimulatory molecule expression and cytokine production. Measurement of the
expression of the surface markers CD1a (A) and CD83 (B) and the cytokines IL-6 (C), IL-12p70 (D), IL-10 (E), and TNF-a (F) of human moDCs 24 h after
stimulation with 1 mMAG, 1 ng/ml LPS, or 1 mMAG and 1 ng/ml LPS. n$ 6. Statistical analysis by two-tailed Wilcoxon signed rank test. The experiment
was repeated three times with similar results. *p , 0.05, **p , 0.01, ***p , 0.001.
The Journal of Immunology 1629
by guest on February 20, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

reduced amount of the phosphorylated protein was confirmed byintracellular staining of phosphorylated p65 and subsequent analysisof fluorescence by flow cytometry (Fig. 5E).
T cell proliferation
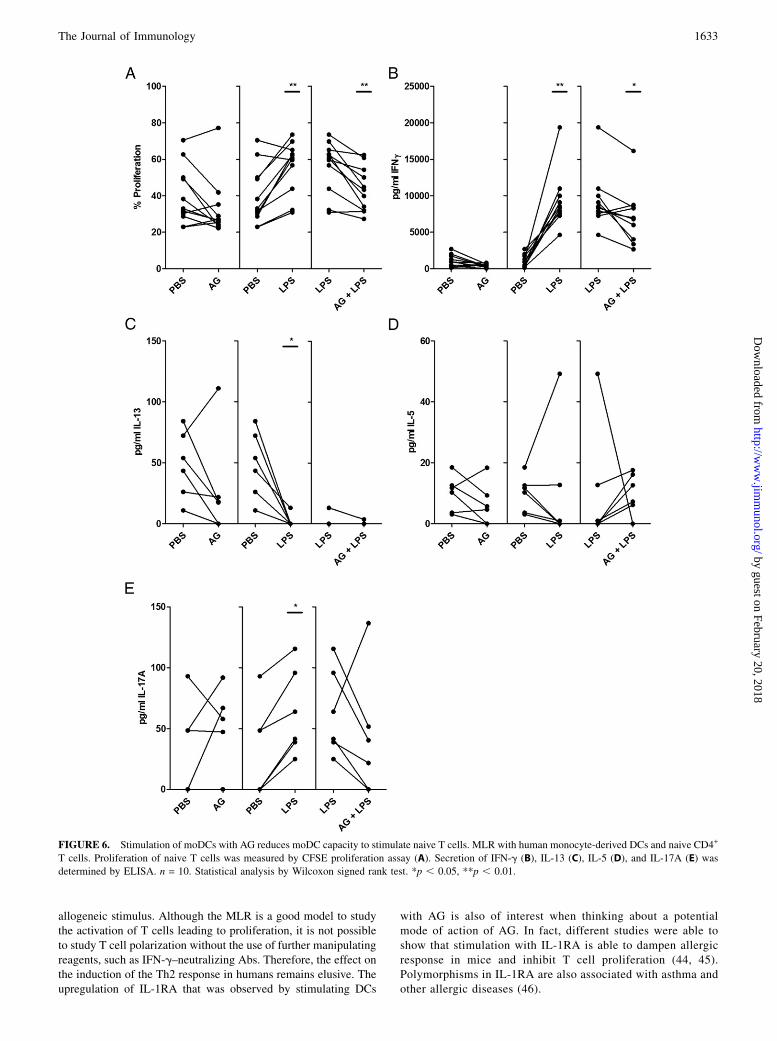
We set up an allogeneicMLR to test whether the influence of AG onthe biology of DCs has an impact on their T cell stimulatory ca-pacity. This was performed in the presence of AG, LPS, or both.Stimulation of DCs with LPS induced proliferation of naive
T cells in allogeneic MLRs. In combination with AG, however, theT cell proliferation was reduced to baseline level (Fig. 6A). Wefurther investigated cytokine production by the measurement ofcell culture supernatants by ELISA. We found that stimulationwith LPS induced IFN-g and IL-17A production, whereas it re-duced IL-13 production. Interestingly, simultaneous stimulationwith AG was able to reduce LPS-induced IFN-g and IL-17A se-cretion, whereas it had no influence on IL-13 (Fig. 6B, 6C, 6E).Production of IL-5 is not affected by stimulation with LPS or byAG (Fig. 6D).
DiscussionRecently, we showed that treatment of mice with AG isolated fromCDE prevents allergic airway inflammation, hyperreactivity, andsensitization (14). The allergy-protective role of AG seems to bemediated by attenuating the Th2-inducing properties of DCs, asshown by sensitization of mice with allergen-pulsed AG-treatedbone marrow–derived DCs. One major problem in all studiesusing substances isolated from the environment is contaminationwith PAMPs. In particular, cell culture models applying cells ofthe innate immune system are strongly affected by PAMPs, suchas LPS, influencing the results when present at concentrations nearthe detection limit. Furthermore, LPS itself was associated withallergy-protective activity by inducing LPS tolerance (8). There-fore, one aim of the present study was to produce contamination-free AG from P. pratense suspension culture. The use of P. pra-tense culture resulted from practical considerations, that is, it grewfaster than the tissue of A. pratensis and was therefore a moreefficient producer of AG. The efficient production of AG would beone major factor for a possible medical use of AG. We have shownpreviously that AG from these two related plants that belong to thesame family is structurally similar (14).The first step of allergic sensitization involves the uptake and
processing of allergens by DCs that, subsequently, induce a Th2response. DCs undoubtedly play an important role in the initiationof the allergic immune response in mice as well as in humans. Wehave chosen human moDCs to study the effects of AG on DCbiology because working with these cells as a model system iswell established and there are strong experimental data from
murine models that this cell type is crucial for orchestrating theasthmatic response (33). At first we sought to identify the receptorto which AG binds. For this purpose, we prepared membranesfrom human moDCs, incubated them with AG, and precipitatedAG-receptor complexes with Yariv’s reagent. Subsequently, weidentified the receptors precipitated by SDS-PAGE and massspectrometry. DC-SIGN and MMR-1 were identified as interestingreceptors with immune-modulating properties that were alreadydescribed in the literature (35). However, the method used here forthe identification of the receptors is limited to the fact thatmembranes are not completely solubilized, resulting in the releaseof small vesicles containing more than one sort of protein. Thisimplies that receptors identified in this way may be more in theclose vicinity of an AG-binding receptor, but do not bind to AGper se. It is possible that Siglec-1 was identified in this artificialway because it is unlikely that it is binding AG owing to the lackof any detectable sialic acid. Therefore, we confirmed binding ofAG to DC-SIGN and MMR-1 in vitro with recombinant receptors.A question that arises from this observation is: why do these plantpolysaccharides bind to mammal pattern recognition receptors?One possible explanation for this would be the similarity betweenAG from plants and microbial polysaccharides in the sense of“molecular mimicry.” For instance, mycobacteria do contain AGand lipoarabinomannan polysaccharides in their cell wall (36). Itis known that the cell walls of mycobacteria contain a variety ofligands of DC-SIGN (37); however, it needs to be defined whetherAG is one of them.
FIGURE 2. AG binds to MMR-1 and DC-SIGN in an inhibitory ELISA. An inhibitory ELISAwith AG as inhibitor was performed to determine the dose-
dependent inhibition of binding of DC-SIGN (A) and MMR-1 (B) to mannan by AG. IC50 values were calculated with one site-binding fit in GraphPad
Prism. Six independent experiments were compared for statistical analysis using the two-tailed Mann–Whitney U test. *p , 0.05.
FIGURE 3. Inhibition of DC-SIGN or MMR-1 partially blocks AG-
induced effects on cytokine secretion. Secretion of IL-6 of human moDCs
24 h after stimulation with 1 mM AG and 1 ng/ml LPS in the presence of
an Ab blocking binding to MMR-1 (A) or DC-SIGN (B). Both in direct
comparison with an appropriate isotype control Ab. n = 4. Statistical
analysis by one-tailed Wilcoxon signed rank test. *p , 0.05.
1630 MODULATION OF DCs BY ARABINOGALACTAN
by guest on February 20, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Activation of DC-SIGN is associated with immune regulation.There are several reports showing that DC-SIGN is targeted by adiversity of pathogens, such as M. tuberculosis, Helicobacterpylori, and HIV-1. This is discussed as an immune escapemechanism of these pathogens (38, 39). These reports describethat stimulation of DC-SIGN acts in a regulatory manner via en-
hancing IL-10 secretion of DCs by different intracellular signaltransduction events depending on the ligand. This may lead toDCs with a lower expression of costimulatory molecules exhib-iting a lower T cell stimulatory capacity in an allogeneic MLRwith CD4+ T cells (34). Ligation of MMR-1 is also linked toimmune regulation. In some reports, activation of MMR-1 on
FIGURE 4. Proteomaps of DCs that were either stimulated with LPS, AG, or LPS plus AG. Data were achieved by analysis of protein expression as
determined by quantitative comparative proteomics (n = 5). The upper panel shows proteins (depicted are the corresponding genes) that are upregulated
upon stimulation and the lower panel shows downregulated proteins in comparison with unstimulated control cells (A). Tiles are arranged and colored
according to the hierarchical Kyoto Encyclopedia of Genes and Genomes pathway maps. Tile size represents the change in expression compared with the
unstimulated sample. Color codes: blue, genetic information processing; brown, metabolism; red, cellular processes; turquoise and magenta, signaling.
Proteins that do not map to any category are shown in the gray area. A detailed assignment of the displayed proteins to cellular processes is shown in (B).
The Journal of Immunology 1631
by guest on February 20, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

human moDCs resulted in IL-10 secretion and reduced T cellstimulatory activity of DCs (40, 41). As expected from thesepublications, we found a significant increase in IL-10 productionof cells that were stimulated with AG alone. Besides IL-10, IL-6was also increased in the supernatant of cells that were stimulatedwith AG alone. IL-6 was previously found to be increased in DCsthat were stimulated with DC-SIGN via a pathway that involvedsignaling via RAF-1 and ERK (42).In contrast, we found decreased production of IL-10 in cells
that were costimulated with AG and LPS compared with cellsthat were stimulated with LPS only. Likewise, other cytokines,for example, IL-6, which is known to be involved in the in-duction of Th2 cells, are reduced due to costimulation of cellswith AG. To test whether this reduction in cytokine productionis due to binding of AG to MMR-1 or DC-SIGN, we have blockedbinding by using neutralizing Abs. Indeed LPS-induced IL-6production was partially restored after blocking either one ofthem, showing that stimulation of these CLRs is involved in AG-mediated downregulation of cytokine production. At first glance,the AG-induced IL-6 and IL-10 production contradicts its sup-pression upon LPS stimulation. However, this may reflect a stateof DC-SIGN–induced tolerance in DCs, which, for example, isalready known for stimulation of DCs with gp120 (19). Thisglycoprotein from HIV also leads to low-level production of cy-tokines such as IL-10 and IL-12 followed by a reduced respon-siveness of the cells to LPS.
Therefore, we studied alterations in the proteome of AG-stimulated DCs in search of mechanisms for the downregulationof DC function. We revealed that AG leads to a distinct pattern ofregulated processes in a comparative study utilizing a label-freequantitative proteomics approach. We show that costimulationwith AG and LPS, but not one substance alone, increases E3ubiquitin-protein ligase TRIM21 expression in moDCs. Moreover,we found that expression of the p65 subunit of NF-kB is reduced inmoDCs that were treated with AG and LPS simultaneously. Thesefindings strongly suggest that costimulation with both substancesleads to the increased expression of TRIM21, which then leads tothe degradation of NF-kB. It has already been shown that TRIM21(also known as RO52) leads to the monoubiquitination of IKKb,and in this manner it leads to decreased NF-kB phosphorylationand activity (43).Our findings suggest that one effect of AG is the activation of
DC-SIGN and MMR-1 resulting in attenuation of DC activation bydanger signals. The reduced expression of costimulatory moleculessupports this assumption. Reduced expression of costimulatorymolecules and proinflammatory cytokines may lead to a lowerT cell stimulatory capacity of DCs, thereby resulting in a reducedT cell response in vitro. Indeed, we observed a reduced T cellstimulatory capacity of AG-stimulated DCs in allogeneic MLRs,resulting in reduced proliferation of T cells and reduced productionof IFN-g. Importantly, note that the MLR used in this study islimited to the fact that a Th1 response is induced due to the strong
FIGURE 5. Stimulation of moDCs with AG and LPS leads to increased expression of TRIM21 and IL-1RA as well as a reduced NF-kB p65 expression
and its phosphorylation. DCs were either stimulated with AG, LPS, or a combination of both. Expression of TRIM21 (A) and IL-1RA (B) in moDCs was
determined by quantitative comparative proteomics (n = 5). Semiquantitative analysess of the expression of the p65 subunit of NF-kB (C) and its phos-
phorylation by ELISA (D) and flow cytometry (E) are shown. The histogram overlay shows the staining of phosphorylated p65 in cells treated with either
medium (light gray), LPS (black), and LPS plus AG (dark gray). The experiment was reproduced twice with similar results. Statistical analysis was done by
two-tailed Wilcoxon signed rank test. **p , 0.01.
1632 MODULATION OF DCs BY ARABINOGALACTAN
by guest on February 20, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
allogeneic stimulus. Although the MLR is a good model to studythe activation of T cells leading to proliferation, it is not possibleto study T cell polarization without the use of further manipulatingreagents, such as IFN-g–neutralizing Abs. Therefore, the effect onthe induction of the Th2 response in humans remains elusive. Theupregulation of IL-1RA that was observed by stimulating DCs
with AG is also of interest when thinking about a potentialmode of action of AG. In fact, different studies were able toshow that stimulation with IL-1RA is able to dampen allergicresponse in mice and inhibit T cell proliferation (44, 45).Polymorphisms in IL-1RA are also associated with asthma andother allergic diseases (46).
FIGURE 6. Stimulation of moDCs with AG reduces moDC capacity to stimulate naive T cells. MLR with human monocyte-derived DCs and naive CD4+
T cells. Proliferation of naive T cells was measured by CFSE proliferation assay (A). Secretion of IFN-g (B), IL-13 (C), IL-5 (D), and IL-17A (E) was
determined by ELISA. n = 10. Statistical analysis by Wilcoxon signed rank test. *p , 0.05, **p , 0.01.
The Journal of Immunology 1633
by guest on February 20, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

In conclusion, we found that AG binds to the human receptorsDC-SIGN and MMR-1 on the surface of DCs. Costimulation ofDCs with AG and LPS leads to decreased NF-kB p65 subunitexpression and phosphorylation, which is likely the reason forreduced DC activation. Furthermore, we showed that AG-treatedDCs have a reduced T cell stimulatory capacity. These observationsand the fact that AG lacks proinflammatory properties make AGan interesting candidate for the development of allergy-preventingcompounds.
AcknowledgmentsWe thank Petra Bonowitz and Sandra Busse at Experimental Pneumology,
Ruhr University Bochum, for excellent technical assistance. We also thank
Philip Saunders (Berlin, Germany) for proofreading the manuscript.
DisclosuresM.P. is a scientific consultant for Protectimmun GmbH. A.B. is an associate
of Protectimmun GmbH but has no other commercial relationship to this
company. The other authors have no financial conflicts of interest.
References1. Strachan, D. P. 1989. Hay fever, hygiene, and household size. BMJ 299: 1259–
1260.2. Strachan, D. P. 2000. Family size, infection and atopy: the first decade of the
“hygiene hypothesis.” Thorax 55(Suppl. 1): S2–S10.3. Sheikh, A., and D. P. Strachan. 2004. The hygiene theory: fact or fiction? Curr.
Opin. Otolaryngol. Head Neck Surg. 12: 232–236.4. Matricardi, P. M. 2010. 99th Dahlem conference on infection, inflammation and
chronic inflammatory disorders: controversial aspects of the “hygiene hypothe-sis.” Clin. Exp. Immunol. 160: 98–105.
5. Ma, Y., J. Zhao, Z. R. Han, Y. Chen, T. F. Leung, and G. W. Wong. 2009. Verylow prevalence of asthma and allergies in schoolchildren from rural Beijing,China. Pediatr. Pulmonol. 44: 793–799.
6. Von Ehrenstein, O. S., E. Von Mutius, S. Illi, L. Baumann, O. Bohm, and R. vonKries. 2000. Reduced risk of hay fever and asthma among children of farmers.Clin. Exp. Allergy 30: 187–193.
7. Riedler, J., C. Braun-Fahrlander, W. Eder, M. Schreuer, M. Waser, S. Maisch,D. Carr, R. Schierl, D. Nowak, and E. von Mutius, ALEX Study Team. 2001.Exposure to farming in early life and development of asthma and allergy: across-sectional survey. Lancet 358: 1129–1133.
8. Braun-Fahrlander, C., J. Riedler, U. Herz, W. Eder, M. Waser, L. Grize,S. Maisch, D. Carr, F. Gerlach, A. Bufe, et al; Allergy and Endotoxin StudyTeam. 2002. Environmental exposure to endotoxin and its relation to asthma inschool-age children. N. Engl. J. Med. 347: 869–877.
9. van Strien, R. T., R. Engel, O. Holst, A. Bufe, W. Eder, M. Waser, C. Braun-Fahrlander, J. Riedler, D. Nowak, and E. von Mutius, ALEX Study Team. 2004.Microbial exposure of rural school children, as assessed by levels of N-acetyl-muramic acid in mattress dust, and its association with respiratory health. J.Allergy Clin. Immunol. 113: 860–867.
10. Peters, M., M. Kauth, J. Schwarze, C. Korner-Rettberg, J. Riedler, D. Nowak,C. Braun-Fahrlander, E. von Mutius, A. Bufe, and O. Holst. 2006. Inhalation ofstable dust extract prevents allergen induced airway inflammation and hyper-responsiveness. Thorax 61: 134–139.
11. Debarry, J., H. Garn, A. Hanuszkiewicz, N. Dickgreber, N. Bl€umer, E. vonMutius, A. Bufe, S. Gatermann, H. Renz, O. Holst, and H. Heine. 2007. Aci-netobacter lwoffii and Lactococcus lactis strains isolated from farm cowshedspossess strong allergy-protective properties. J. Allergy Clin. Immunol. 119:1514–1521.
12. Stiehm, M., A. Bufe, and M. Peters. 2013. Proteolytic activity in cowshed dustextracts induces C5a release in murine bronchoalveolar lavage fluids which mayaccount for its protective properties in allergic airway inflammation. Thorax 68:31–38.
13. Stiehm, M., K. Peters, K.-H. Wiesm€uller, A. Bufe, and M. Peters. 2013. A novelsynthetic lipopeptide is allergy-protective by the induction of LPS-tolerance.Clin. Exp. Allergy 43: 785–797.
14. Peters, M., M. Kauth, O. Scherner, K. Gehlhar, I. Steffen, P. Wentker, E. vonMutius, O. Holst, and A. Bufe. 2010. Arabinogalactan isolated from cowsheddust extract protects mice from allergic airway inflammation and sensitization. J.Allergy Clin. Immunol. 126: 648–656.e1, 4.
15. Thude, S., B. Classen, W. Blaschek, D. Barz, and H. Thude. 2006. Bindingstudies of an arabinogalactan-protein from Echinacea purpurea to leucocytes.Phytomedicine 13: 425–427.
16. Grønhaug, T. E., P. Ghildyal, H. Barsett, T. E. Michaelsen, G. Morris, D. Diallo,M. Inngjerdingen, and B. S. Paulsen. 2010. Bioactive arabinogalactans from theleaves of Opilia celtidifolia Endl. ex Walp. (Opiliaceae). Glycobiology 20: 1654–1664.
17. Holderness, J., I. A. Schepetkin, B. Freedman, L. N. Kirpotina, M. T. Quinn,J. F. Hedges, M. A. Jutila, and J. Zimmer. 2011. Polysaccharides isolated fromAcaı fruit induce innate immune responses. PLoS One 6: e17301.
18. Geijtenbeek, T. B. H., G. C. van Duijnhoven, S. J. van Vliet, E. Krieger,G. Vriend, C. G. Figdor, and Y. van Kooyk. 2002. Identification of differentbinding sites in the dendritic cell-specific receptor DC-SIGN for intercellularadhesion molecule 3 and HIV-1. J. Biol. Chem. 277: 11314–11320.
19. Shan, M., P. J. Klasse, K. Banerjee, A. K. Dey, S. P. Iyer, R. Dionisio, D. Charles,L. Campbell-Gardener, W. C. Olson, R. W. Sanders, and J. P. Moore. 2007. HIV-1gp120 mannoses induce immunosuppressive responses from dendritic cells. PLoSPathog. 3: e169.
20. Park, C. G., K. Takahara, E. Umemoto, Y. Yashima, K. Matsubara, Y. Matsuda,B. E. Clausen, K. Inaba, and R. M. Steinman. 2001. Five mouse homologues ofthe human dendritic cell C-type lectin, DC-SIGN. Int. Immunol. 13: 1283–1290.
21. Takahara, K., Y. Yashima, Y. Omatsu, H. Yoshida, Y. Kimura, Y. S. Kang,R. M. Steinman, C. G. Park, and K. Inaba. 2004. Functional comparison of themouse DC-SIGN, SIGNR1, SIGNR3 and Langerin, C-type lectins. Int. Immunol.16: 819–829.
22. Tanne, A., B. Ma, F. Boudou, L. Tailleux, H. Botella, E. Badell, F. Levillain,M. E. Taylor, K. Drickamer, J. Nigou, et al. 2009. A murine DC-SIGN homo-logue contributes to early host defense against Mycobacterium tuberculosis. J.Exp. Med. 206: 2205–2220.
23. Lech, M., H. E. Susanti, C. Rommele, R. Grobmayr, R. G€unthner, andH.-J. Anders. 2012. Quantitative expression of C-type lectin receptors in hu-mans and mice. Int. J. Mol. Sci. 13: 10113–10131.
24. Yariv, J., M. M. Rapport, and L. Graf. 1962. The interaction of glycosides andsaccharides with antibody to the corresponding phenylazo glycosides. Biochem.J. 85: 383–388.
25. Peters, M., P. Fritz, and A. Bufe. 2012. A bioassay for determination of lipo-polysaccharide in environmental samples. Innate Immun. 18: 694–699.
26. Gollner, E. M., W. Blaschek, and B. Classen. 2010. Structural investigations onarabinogalactan-protein from wheat, isolated with Yariv reagent. J. Agric. FoodChem. 58: 3621–3626.
27. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of thehead of bacteriophage T4. Nature 227: 680–685.
28. Megger, D. A., T. Bracht, M. Kohl, M. Ahrens,W. Naboulsi, F.Weber, A.-C. Hoffmann,C. Stephan, K. Kuhlmann, M. Eisenacher, et al. 2013. Proteomic differences betweenhepatocellular carcinoma and nontumorous liver tissue investigated by a com-bined gel-based and label-free quantitative proteomics study. Mol. Cell. Pro-teomics 12: 2006–2020.
29. Padden, J., D. A. Megger, T. Bracht, H. Reis, M. Ahrens, M. Kohl,M. Eisenacher, J. F. Schlaak, A. E. Canbay, F. Weber, et al. 2014. Identificationof novel biomarker candidates for the immunohistochemical diagnosis of chol-angiocellular carcinoma. Mol. Cell. Proteomics 13: 2661–2672.
30. Sallusto, F., and A. Lanzavecchia. 1994. Efficient presentation of soluble antigenby cultured human dendritic cells is maintained by granulocyte/macrophagecolony-stimulating factor plus interleukin 4 and downregulated by tumor ne-crosis factor alpha. J. Exp. Med. 179: 1109–1118.
31. Grage-Griebenow, E., D. Lorenzen, R. Fetting, H. D. Flad, and M. Ernst. 1993.Phenotypical and functional characterization of Fcg receptor I (CD64)-negativemonocytes, a minor human monocyte subpopulation with high accessory andantiviral activity. Eur. J. Immunol. 23: 3126–3135.
32. Liebermeister, W., E. Noor, A. Flamholz, D. Davidi, J. Bernhardt, and R. Milo.2014. Visual account of protein investment in cellular functions. Proc. Natl.Acad. Sci. USA 111: 8488–8493.
33. Plantinga, M., M. Guilliams, M. Vanheerswynghels, K. Deswarte, F. Branco-Madeira, W. Toussaint, L. Vanhoutte, K. Neyt, N. Killeen, B. Malissen, et al.2013. Conventional and monocyte-derived CD11b+ dendritic cells initiate andmaintain T helper 2 cell-mediated immunity to house dust mite allergen. Im-munity 38: 322–335.
34. Svajger, U., N. Obermajer, M. Anderluh, J. Kos, and M. Jeras. 2011. DC-SIGNligation greatly affects dendritic cell differentiation from monocytes compro-mising their normal function. J. Leukoc. Biol. 89: 893–905.
35. Gringhuis, S. I., J. den Dunnen, M. Litjens, M. van der Vlist, and T. B.H. Geijtenbeek. 2009. Carbohydrate-specific signaling through the DC-SIGNsignalosome tailors immunity to Mycobacterium tuberculosis, HIV-1 and Heli-cobacter pylori. Nat. Immunol. 10: 1081–1088.
36. Jankute, M., J. A. G. Cox, J. Harrison, and G. S. Besra. 2015. Assembly of themycobacterial cell wall. Annu. Rev. Microbiol. 69: 405–423.
37. Wheat, W. H., R. Dhouib, S. K. Angala, G. Larrouy-Maumus, K. Dobos,J. Nigou, J. S. Spencer, and M. Jackson. 2015. The presence of a galactosaminesubstituent on the arabinogalactan of Mycobacterium tuberculosis abrogates fullmaturation of human peripheral blood monocyte-derived dendritic cells andincreases secretion of IL-10. Tuberculosis (Edinb.) 95: 476–489.
38. Gringhuis, S. I., J. den Dunnen, M. Litjens, M. van der Vlist, B. Wevers,S. C. Bruijns, and T. B. Geijtenbeek. 2009. Dectin-1 directs T helper cell dif-ferentiation by controlling noncanonical NF-kB activation through Raf-1 andSyk. Nat. Immunol. 10: 203–213.
39. Geijtenbeek, T. B. H., S. J. Van Vliet, E. A. Koppel, M. Sanchez-Hernandez,C. M. J. E. Vandenbroucke-Grauls, B. Appelmelk, and Y. Van Kooyk. 2003.Mycobacteria target DC-SIGN to suppress dendritic cell function. J. Exp. Med.197: 7–17.
40. Chieppa, M., G. Bianchi, A. Doni, A. Del Prete, M. Sironi, G. Laskarin,P. Monti, L. Piemonti, A. Biondi, A. Mantovani, et al. 2003. Cross-linking of themannose receptor on monocyte-derived dendritic cells activates an anti-inflammatory immunosuppressive program. J. Immunol. 171: 4552–4560.
41. Allavena, P., M. Chieppa, G. Bianchi, G. Solinas, M. Fabbri, G. Laskarin, andA. Mantovani. 2010. Engagement of the mannose receptor by tumoral mucinsactivates an immune suppressive phenotype in human tumor-associated macro-phages. Clin. Dev. Immunol. 2010: 1–10.
1634 MODULATION OF DCs BY ARABINOGALACTAN
by guest on February 20, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

42. Hsu, S.-C., C.-H. Chen, S.-H. Tsai, H. Kawasaki, C.-H. Hung, Y.-T. Chu,H.-W. Chang, Y. Zhou, J. Fu, B. Plunkett, et al. 2010. Functional interaction ofcommon allergens and a C-type lectin receptor, dendritic cell-specific ICAM3-grabbing non-integrin (DC-SIGN), on human dendritic cells. J. Biol. Chem. 285:7903–7910.
43. Wada, K., M. Niida, M. Tanaka, and T. Kamitani. 2009. Ro52-mediated mon-oubiquitination of IKKb down-regulates NF-kB signalling. J. Biochem. 146:821–832.
44. Ortiz, L. A., M. Dutreil, C. Fattman, A. C. Pandey, G. Torres, K. Go, andD. G. Phinney. 2007. Interleukin 1 receptor antagonist mediates the antiin-
flammatory and antifibrotic effect of mesenchymal stem cells during lung injury.Proc. Natl. Acad. Sci. USA 104: 11002–11007.
45. Hudock, K. M., Y. Liu, J. Mei, R. C. Marino, J. E. Hale, N. Dai, andG. S. Worthen. 2012. Delayed resolution of lung inflammation in Il-1rn2/2 micereflects elevated IL-17A/granulocyte colony-stimulating factor expression. Am.J. Respir. Cell Mol. Biol. 47: 436–444.
46. Ramadas, R. A., A. Sadeghnejad, W. Karmaus, S. H. Arshad, S. Matthews,M. Huebner, D.-Y. Kim, and S. L. Ewart. 2007. Interleukin-1R antagonist geneand pre-natal smoke exposure are associated with childhood asthma. Eur. Respir.J. 29: 502–508.
The Journal of Immunology 1635
by guest on February 20, 2018http://w
ww
.jimm
unol.org/D
ownloaded from


















