
Alfalfa Cyclins: Differential Expression during the Cell ... · 390 420 caa gag aca ctg tit cti aca...
8
The Plant Cell, Vol. 4, 1531-1538, December 1992 O 1992 American Society of Plant Physiologists Alfalfa Cyclins: Differential Expression during the Cell Cycle and in Plant Organs Heribert Hirt,' Mhtyhs Mink,* Martin Pfo~ser,~ Laszlo Bogre, Jdnos Gyorgyey,* Claudia Jonak, Anton Gartner,4 Dhes Dudits,* and Erwin Heberle-Bors lnstitute of Microbiology and Genetics, University of Vienna, Vienna Biocenter, Dr. Bohrgasse 9, 1030 Vienna, Austria Cell division in eukaryotes is mediated by the action of the mitosis promoting factor, which is composed of the CDC2 protein kinase and one of the various mitotic cyclins. We have recently isolated a cdc2 gene from alfalfa. Here, we report the isolation of two cyclin genes, cycMs7 and cycMs2, from alfalfa. The cycMs2 gene shows highest similarity to type 6 cyclins. In contrast, the predicted amino acid sequence of the cycMs7 gene shows similar homology scores to cyclins of all types (25 to 35%). 60th genes are expressed in dividing suspension cultured cells but cease to be expressed when the cells enter stationary phase. In synchronized alfalfa suspension cultured cells, the mRNAs of cycMs7 and cycMs2 show maximal expression in the 62 and M phases. Transcripts of cycMs2 are found only in late 6 2 and M phase cells, an expression pattern typical for cyclin B genes, whereas cycMs7 appears with the onset of 62. This pattern indicates that alfalfa cycMs7 and cycMs2 belong to different classes of cyclins. In young leaves, expression of both genes is high, whereas in mature leaves no transcripts can be detected, indicating that the two cyclin genes are true cell division mark- ers at the mRNA level. In other organs, a more complex expression pattern of the two cyclin genes was found. INTRODUCTION All eukaryotes have been shown to contain a 34-kD protein kinase that plays a central role in the regulation of cell divi- sion. The 34-kD protein kinase has been shown to be the product of the cdc2+/CDC28 genes in yeasts and animals (for review, see Nurse, 1990) and in various plant species (Colasanti et al., 1991; Ferreira et al., 1991; Hirt et al., 1991). The CDC2 kinase is one part of the mitosis promoting fac- tor: its activation involves physical association of ~ 3 4 ~ ~ ~ with cyclin proteins. In the absence of cyclins, cells cannot enter mitosis and CDC2 kinase activity is absent (Booher and Beach, 1987,1988; Minshullet al., 1989; Murray and Kirschner, 1989; Murray et al., 1989). At the end of metaphase, cyclins are de- stroyed and CDC2 kinaseactivity vanishes(Murray et al., 1989). Degradationof cyclins has been shown to be requiredfor com- pletion of mitosis and might be necessary for inactivation of the CDC2 kinase (Murray et al., 1989). Cyclins were originally identified in the eggs of marine in- vertebrates(Evanset al., 1983; Swenson et al., 1986; Standart et al., 1987). On the basis of amino acid homology, cyclins have subsequently been identified in fission yeast (Booher and To whom correspondenceshould be addressed. Current address: lnstitute of Plant Physiology, Biological Research Center, Hungarian Academy of Sciences, H-6701 Szeged, POB 521, Hungary. lnstitute of Botany, University of Agriculture and Forestry, Gregor- Mendelstr. 33, A-1190 Vienna, Austria. Current address: lnstitute of Molecular Pathology, Dr. Bohrgasse 3, A-1030 Vienna, Austria. Beach, 1988; Goebl and Byers, 1988; Hagan et al., 1988; Solomon et al., 1988), Xenopus (Minshull et al., 1989), Dro- sophila (Lehner and OFarrell, 1989; Whitfield et al., 1989), humans (Pines and Hunter, 1989), and recently also in carrot, soybean (Hata et al., 1991), and Arabidopsis (Hemerly et al., 1992). In this study, we describe the isolation of two partia1cyclin cDNA clones, cycMs7 and cycMs2, from alfalfa. Comparison of the predictedcyclin protein sequences with 18 other cyclins showed that cycMs2 can be classified as a type B cyclin. In contrast, cycMs7 appears to be equally distantly relatedto the type A, B, and G1 cyclins. RNA gel blot analysis with syn- chronized alfalfa cell suspension cultures showed cell cycle-dependent expression of both cyclin genes in the G2 and M phases. Duringthe growth cycle of a batch suspension culture, the two genes were expressed during exponential growth phase but not in stationary phase. In the plant, cycMs7 and cycMs2 were only found to be expressed in organs with dividing cells. RESULTS lsolation and Sequencing of Cyclin cDNAs from Alfalfa To isolate cyclins of type A and B, we screened 80,000 colo- nies of an alfalfa (Medicago sariva) cDNA library made from
Transcript of Alfalfa Cyclins: Differential Expression during the Cell ... · 390 420 caa gag aca ctg tit cti aca...

The Plant Cell, Vol. 4, 1531-1538, December 1992 O 1992 American Society of Plant Physiologists
Alfalfa Cyclins: Differential Expression during the Cell Cycle and in Plant Organs
Heribert Hirt,' Mhtyhs Mink,* Martin P f o ~ s e r , ~ Laszlo Bogre, Jdnos Gyorgyey,* Claudia Jonak, Anton Gartner,4 Dhes Dudits,* and Erwin Heberle-Bors
lnstitute of Microbiology and Genetics, University of Vienna, Vienna Biocenter, Dr. Bohrgasse 9, 1030 Vienna, Austria
Cell division in eukaryotes is mediated by the action of the mitosis promoting factor, which is composed of the CDC2 protein kinase and one of the various mitotic cyclins. We have recently isolated a cdc2 gene from alfalfa. Here, we report the isolation of two cyclin genes, cycMs7 and cycMs2, from alfalfa. The cycMs2 gene shows highest similarity to type 6 cyclins. In contrast, the predicted amino acid sequence of the cycMs7 gene shows similar homology scores to cyclins of all types (25 to 35%). 60th genes are expressed in dividing suspension cultured cells but cease to be expressed when the cells enter stationary phase. In synchronized alfalfa suspension cultured cells, the mRNAs of cycMs7 and cycMs2 show maximal expression in the 62 and M phases. Transcripts of cycMs2 are found only in late 6 2 and M phase cells, an expression pattern typical for cyclin B genes, whereas cycMs7 appears with the onset of 62. This pattern indicates that alfalfa cycMs7 and cycMs2 belong to different classes of cyclins. In young leaves, expression of both genes is high, whereas in mature leaves no transcripts can be detected, indicating that the two cyclin genes are true cell division mark- ers at the mRNA level. In other organs, a more complex expression pattern of the two cyclin genes was found.
INTRODUCTION
All eukaryotes have been shown to contain a 34-kD protein kinase that plays a central role in the regulation of cell divi- sion. The 34-kD protein kinase has been shown to be the product of the cdc2+/CDC28 genes in yeasts and animals (for review, see Nurse, 1990) and in various plant species (Colasanti et al., 1991; Ferreira et al., 1991; Hirt et al., 1991).
The CDC2 kinase is one part of the mitosis promoting fac- tor: its activation involves physical association of ~ 3 4 ~ ~ ~ with cyclin proteins. In the absence of cyclins, cells cannot enter mitosis and CDC2 kinase activity is absent (Booher and Beach, 1987,1988; Minshull et al., 1989; Murray and Kirschner, 1989; Murray et al., 1989). At the end of metaphase, cyclins are de- stroyed and CDC2 kinase activity vanishes (Murray et al., 1989). Degradation of cyclins has been shown to be required for com- pletion of mitosis and might be necessary for inactivation of the CDC2 kinase (Murray et al., 1989).
Cyclins were originally identified in the eggs of marine in- vertebrates(Evans et al., 1983; Swenson et al., 1986; Standart et al., 1987). On the basis of amino acid homology, cyclins have subsequently been identified in fission yeast (Booher and
To whom correspondence should be addressed. Current address: lnstitute of Plant Physiology, Biological Research
Center, Hungarian Academy of Sciences, H-6701 Szeged, POB 521, Hungary.
lnstitute of Botany, University of Agriculture and Forestry, Gregor- Mendelstr. 33, A-1190 Vienna, Austria. Current address: lnstitute of Molecular Pathology, Dr. Bohrgasse 3,
A-1030 Vienna, Austria.
Beach, 1988; Goebl and Byers, 1988; Hagan et al., 1988; Solomon et al., 1988), Xenopus (Minshull et al., 1989), Dro- sophila (Lehner and OFarrell, 1989; Whitfield et al., 1989), humans (Pines and Hunter, 1989), and recently also in carrot, soybean (Hata et al., 1991), and Arabidopsis (Hemerly et al., 1992).
In this study, we describe the isolation of two partia1 cyclin cDNA clones, cycMs7 and cycMs2, from alfalfa. Comparison of the predicted cyclin protein sequences with 18 other cyclins showed that cycMs2 can be classified as a type B cyclin. In contrast, cycMs7 appears to be equally distantly related to the type A, B, and G1 cyclins. RNA gel blot analysis with syn- chronized alfalfa cell suspension cultures showed cell cycle-dependent expression of both cyclin genes in the G2 and M phases. During the growth cycle of a batch suspension culture, the two genes were expressed during exponential growth phase but not in stationary phase. In the plant, cycMs7 and cycMs2 were only found to be expressed in organs with dividing cells.
RESULTS
lsolation and Sequencing of Cyclin cDNAs from Alfalfa
To isolate cyclins of type A and B, we screened 80,000 colo- nies of an alfalfa (Medicago sariva) cDNA library made from

1532 The Plant Cell
suspension cultured cells that were induced to form somatic embryos with redundant oligonucleotides encoding the con- served amino acid sequences K-Y-E-E-M(I)-Y-P for the cyclin A and B type and I-L-V(I)-D-W-L-V for the cyclin B class (see Methods for the nucleotide sequences). Positive colonies were isolated and rescreened according to the method of Hanks et al. (1988) at a temperature of 58OC in order to distinguish between false positives with a fit of less than 16 nucleotides. Only one colony was found to hybridize under these condi- tions to the I-L-V(I)-D-W-L-V oligonucleotide. DNA was prepared from this clone and digested with Pstl. Upon gel electropho- resis, a 750-bp insert was observed.
Because the insert contains GC tails at both ends, it was impossible to perform sequence analysis with this clone. A 400-bp BamHl fragment was subcloned into the vector pTz19U and sequenced from both sides. Sequencing was also possi- ble after removal of a 5'200-bp Xbal fragment from the original clone. The complete sequence is shown in Figure 1A. The se- quence at position 751 to 771 shows a fit of 18 out of the 20 nucleotides to the redundant oligonucleotide probe of the cy- clin B class (see above). However, at the amino acid levei, only five of the encoded seven amino acids fit to the highly con- served cyclin B box (the encoded sequence reads I-L-V-D-C-L-L instead of I-L-V-D-W-L-V). The clone contains a nontranslated region of 132 nucleotides before the first ATG, which is followed by a putative sequence of 211 amino acids. However, there is no stop codon or any indication of a poly(A) tail in the clone. This is surprising because the library was constructed by oligo(dT) priming of the poly(A)+ RNA. Rescreening of 250,000 colonies of the same library and 1,000,000 of another cDNA library, made from alfalfa somatic embryos, resulted in the isolation of two colonies that were found to be identical to the above described clone. Screening of a third cDNA li- brary made from suspension cultured alfalfa cells that were induced to form somatic embryos resulted in the isolation of one clone of 1.4 kb. The cDNA sequence is shown in Figure 18 and was found to be very different from that of CycMsl. A 5'truncated open reading frame of 984 nucleotides was iden- tified and appears to encode a different cyclin that is designated cycMs2. Interestingly, in contrast to CycMsl, the cycMs2 clone contains a cyclin B box (nucleotide position 313 to 333) which fits much better to the animal sequences. Rescreening the different libraries yielded no other hybridizing colony than the one indicated. These results indicate high instability andlor very low abundance of these types of mRNAs.
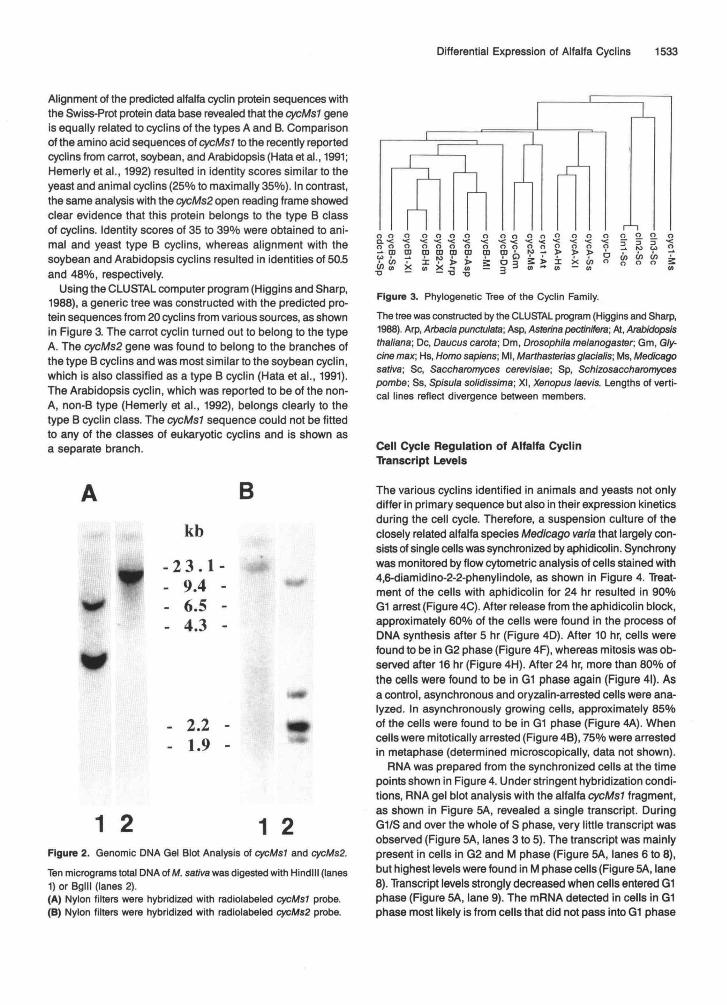
To determine the copy number of the CycMsl and cycM.92 genes in alfalfa, DNA gel blot analysis was performed with genomic DNAs from M. sativa. Hybridization with a 32P- labeled cycMs7 probe, as shown in Figure 2A, revealed the presence of two bands in Hindlll-digested total DNA (Figure 2A, lane 1) but only one band in Bglll-digested DNA (Figure 2A, lane 2). When the same DNAs were probed with a radio- labeled cycMs2 probe, as shown in Figure 26, the Hindlll digest revealed hybridization to one band (Figure 28, lane 1) but to four bands in Bglll-digested DNA (Figure 28, lane 2). Because alfalfa is an autotetraploid species, these data indicate the oc- currence of one copy per haploid genome for both cyclin genes.
A Zs"::C AAC TIlC TCT CAC TGT TTC TCT CTT CCA m TrT TTC AAG GRT CTT CAT TTT CRC CCC CTT CTT CTT T T A GGA TCT TCC A T T TTT TTI TGT uzj TTG TGT TTC A T T 'M"I TTT TTC A T T GAA GAA AAA ATG M G m TCT GAG GIG AAG AAT GTI TCT AAC AAC CCC ACA AAT m
210 2 4 0 G M GGA GGG TTA GAT TCT AGA AAG GTT UX: GAA AAC AGI AGA G C I TTG GGT GTG A T T AAT E G G L O S R K Y C E N R R A L G V I N
210 300 CAG AAT TTG GTT GTG CAA GGA CGT CCT TAT CCT TGT GTT GTI AAC AAG AGG GCA TTG TCA O N L Y V C G R P Y P C Y Y N X R A L S
330 360 GAG IGA AAT GAT GTT TGT GAG AAG A M C M GCG GAT CCG GTI CAT CGI CCC ATC ACT AGG
390 420 AGG TTT GCT GCA AAG ATT GCT AGE ACA A&A ACA TCT U T GCT GIG GGA ACT ACC AAA AGG
450 480 TCA AAC TTG GCA AAA TCA AGT TCA AAT GGA VIT CGA GAT Tpc ATA m GTT GAT GAT GAA S N L A K S S S N G F G O T I F Y D D E
510 540 CAC AAC CCA GTG GAC GAT CAG CCA GTG CCA ATG CCT TTA GAG CAA ACA GAA CCA ATC CAT
510 600 ACT GAA TCA GAT CAG ATG GAG OM GTI GAG ATG GAG GAT ATC ATG GAA GAG CCT CTI ATG S E S D Q H E C V E I E D I I E E P V I
630 660 GAC ATI GAC ACC CCT GAT GCA AAT CIC CCT CTI CCA GTT CCT OM TAT A T I G14 GAT CTI D I D T P D A N D P L A V A E Y I E D L
690 720 TAC TCT TAC TAC AGI AAA GTT GAG AGT ACT AGC TGT GTI TCA CCA AAC TAT li% GCA CAG
H K ~ S E E I N Y S N N P T N F
E R N D V C E K K Q A D P V H R P I T R
P ~ A A X I A S T X T S N A E G T T K R
H K P V E D O P V P I A L E Q T E P I H
Y S ~ Y R R V E S T S C Y S P N Y H A Q 750
C M TTT GAC ATT AAT GAA AGG ATG AGG GCT ATA CTG GTT GAC xc TTA Tn; Q F D l N E R l R A l L V D C L L
60 AAT TCG AAT GAG 'M"I GGA AAC Tpc ATC GCC ATP GAT GAT OM CTC M G TIA CCA GM GAC
90 120 C M CCA GAG CCT ATG ACT TTG OIG CAT ACT GAA CCA ATG CAT AGT GAT CCA CTT GAG ATG
150 180 GUL GAG CTT GAG ATG GAG GAT ATA GAG uill GAG ATG A m TX GAC ATT GAT AGC TGT GAT E E V f M E D I E G C M I L D l D S C D
210 240 GCA M C M T TCT CTI GCT GTI GTC GAG TAT ATT GAA GAT CTC CAT GCT TAC TAC AGA M G k W N S L A V V E Y I E D L H A Y I R K
210 300 ATT GAG TAT CTI GGC TGT GTI TCA C U ACC TAC ATG GAT GM CAA CTI GAC C M AAT GAG
330 360 A% ATG AGG GCT ATA CTA GlT GAC TGG CTG A m GAG GTT CAT GAC AAA TTC GAC CTC ATG R M R I I L V D Y L I E V H D K F D L M
390 420 CAA GAG ACA CTG TIT CTI ACA GTT M T CTC ATC GAC AGA TIC TX GCT AAG CAG AAT GTT
450 480 CTI AGA AAG AAA CTT CAG CTG 01T GGT CTT GTT GCC ATG CTI TX GCA TGC AAG TAT GAG V R R K L Q L V G L V I R L L A C X T E
510 540 G M GTT TCT GTG CCT GTC GlT W G GAT CTG ATC CAT ATA GCT GAC AGA GCT TAC ACA AGG
570 600 AAA GAC A T T CTC G M ATG GAG AAG TTG ATG CTC M C ACT CTC CAG TAT AAC ATG TCT CTT
630 660 CCA ACT GCT TIT GTT TTC ATG AGA AGG 2 7 7 CTG AAG GCT GCT CAA GCT CAC luu AAA CTI P T A Y Y F M R R F L K ~ A a I D r , K L
690 120 GAG CTG CTG GCT m Tpc TTG GTT GIC CTA TCT CTG GTG GM TAT GAG I\TG CTG AAC m E L V I F F L V D L S L Y E Y E M L K F
150 780
CCA CCA TCC TTG GTI GCT GCT GCT GCT CTC TAC ACA GCT C M TGC ACT GTC AGT GGT TTC
cyatI52 30
N S N E F G N F I A I D D E L K L P E O
~ P E P * T L E H T E P * H ~ D P L E u
I E Y L C C V S P T Y M D E Q L D L N E
~ E T L F L T ~ N L ~ D R F L A K Q N V
~ ~ S ~ P ~ V S D L I H I A D R ~ Y T R
K D I L E M ~ K L I L N T L O Y N M S L .
P P S L V A A A A V Y T I Q C T Y S G P 810 840
AAA CAC TGG AAC AAG ACA TGT GAG TCG CAC ACI UIC TAC TCA GAA GAT CAG CTG TIG GAG K H W N K T C E Y H T N I S E D Q L L E
810 900 TGC TCT ATG CTC ATG CTT GGA TIC CAC CAG AAA GCC GGA GCC ujc A&A CTG >.2k GGA GTG
930 960 CAT AGG AAG TAT GGC TCA GCA AAA TTI AGC Tpc ACCT GCT AAA TGT GAA CCA GCA TGT TlT
C S ~ L H V G F H Q K A G A G K L T G V
H R K I G S A K I S F T A K C E P A C F 990 1020
CTI CTG GAG AAC AAG AAC CAG CCA TAA GAA GAG AGC CAG CCA TGT ATA GCT TIT CTT ACA
AAC AAT ACC ATA AGT GTG CTG CTG ATT TTG UX: GGT T I A AGA AGA ACA AAA CAT AAA GAG C M CTT TGG TTG CTT TGT ACT TIT CTC AAG A M TIC TGT TAA GTI ACT m ATT CTI TGG TCA T T I T T A TlT TIG TTG TTC ATG Tpc CAT AGG TGA ACA TAA T M m CAG AAC m m ATT ATG TCT TCT GTA TTT TCT CTT TGA ACA AAA TAA TGG GAT ATA GAC ACA ATT m GTC CCT T T A AAA A M AAA AAA AAA AA
L L E N K W Q P
TAT crc ccc m v r TGA TGC m TAT AAC TTG mr =li TGC AAC TTC AAT TCA ATT TTA
Figure 1. Nucleotide Sequences of Alfalfa cycMs7 and cycMs2 cDNAs and Their Predicted Arnino Acid Sequences.
(A) cycMs7. The EMBL accession nurnber is X68740. (e) cycMs2. The EMBL accession number is X68741.
Therefore, we conclude that the cycMsl and cycMs2 genes are both present as single copy genes in alfalfa.
The cycMs2 Gene Encodes a Member of the Type B Cyclins, but cycMsl Cannot Be Classified into Any Cyclin Class
Alignment of the predicted protein sequences from cycMsl and cycMs2 revealed 65% identity over 111 amino acids.
Differential Expression of Alfalfa Cyclins 1533
Alignment of the predicted alfalfa cyclin protein sequences withthe Swiss-Prat protein data base revealed that the cycMsl geneis equally related to cyclins of the types A and B. Comparisonof the amino acid sequences of cycMsl to the recently reportedcyclins from carrot, soybean, and Arabidopsis (Hataet al., 1991;Hemerly et al., 1992) resulted in identity scores similar to theyeast and animal cyclins (25% to maximally 35%). In contrast,the same analysis with the cycMs2 open reading frame showedclear evidence that this protein belongs to the type B classof cyclins. Identity scores of 35 to 39% were obtained to ani-mal and yeast type B cyclins, whereas alignment with thesoybean and Arabidopsis cyclins resulted in identities of 50.5and 48%, respectively.
Using the CLUSTAL computer program (Higgins and Sharp,1988), a generic tree was constructed with the predicted pro-tein sequences from 20 cyclins from various sources, as shownin Figure 3. The carrot cyclin turned out to belong to the typeA. The cycMs2 gene was found to belong to the branches ofthe type B cyclins and was most similar to the soybean cyclin,which is also classified as a type B cyclin (Hata et al., 1991).The Arabidopsis cyclin, which was reported to be of the non-A, non-B type (Hemerly et al., 1992), belongs clearly to thetype B cyclin class. The cycMsl sequence could not be fittedto any of the classes of eukaryotic cyclins and is shown asa separate branch.
Bkb
2 3 . 1 -9.4 -6.5 -4.3 -
- 2.2 -- 1.9 -
1 2 1 2Figure 2. Genomic DMA Gel Blot Analysis of cycMsl and cycMs2.
Ten micrograms total DNA of M. sativa was digested with Hindlll (lanes1) or Bglll (lanes 2).(A) Nylon filters were hybridized with radiolabeled cycMsl probe.(B) Nylon filters were hybridized with radiolabeled cycMs2 probe.
t o r
1
>5
11
c •> o "o *f
1
r1
$ ->
~i
M r*- o*<
- L O 3 0 3 D O D D D D D D D DV in ? i V > > ±w « x » x ? M s of i i i— g o» ** « « 1
Figure 3. Phylogenetic Tree of the Cyclin Family.The tree was constructed by the CLUSTAL program (Higgins and Sharp,1988). Arp, Arbacia punctulata; Asp, Asterina pectinifera; At, Arabidopsisthaliana; DC, Daucus carota; Dm, Drosophila melanogaster; Gm, Gly-cine max; Hs, Homo sapiens; Ml, Marthasterias glacialis; Ms, Medicagosativa; Sc, Saccharomyces cerevisiae; Sp, Schizosaccharomycespomto; Ss, Spisula solidissima; XI, Xenopus laevis. Lengths of verti-cal lines reflect divergence between members.
Cell Cycle Regulation of Alfalfa CyclinTranscript Levels
The various cyclins identified in animals and yeasts not onlydiffer in primary sequence but also in their expression kineticsduring the cell cycle. Therefore, a suspension culture of theclosely related alfalfa species Medicago varia that largely con-sists of single cells was synchronized by aphidicolin. Synchronywas monitored by flow cytometric analysis of cells stained with4,6-diamidino-2-2-phenylindole, as shown in Figure 4. Treat-ment of the cells with aphidicolin for 24 hr resulted in 90%G1 arrest (Figure 4C). After release from the aphidicolin block,approximately 60% of the cells were found in the process ofDNA synthesis after 5 hr (Figure 4D). After 10 hr, cells werefound to be in G2 phase (Figure 4F), whereas mitosis was ob-served after 16 hr (Figure 4H). After 24 hr, more than 80% ofthe cells were found to be in G1 phase again (Figure 41). Asa control, asynchronous and oryzalin-arrested cells were ana-lyzed. In asynchronously growing cells, approximately 85%of the cells were found to be in G1 phase (Figure 4A). Whencells were mitotically arrested (Figure 4B), 75% were arrestedin metaphase (determined microscopically, data not shown).
RNA was prepared from the synchronized cells at the timepoints shown in Figure 4. Under stringent hybridization condi-tions, RNA gel blot analysis with the alfalfa cycMsl fragment,as shown in Figure 5A, revealed a single transcript. DuringG1/S and over the whole of S phase, very little transcript wasobserved (Figure 5A, lanes 3 to 5). The transcript was mainlypresent in cells in G2 and M phase (Figure 5A, lanes 6 to 8),but highest levels were found in M phase cells (Figure 5A, lane8). Transcript levels strongly decreased when cells entered G1phase (Figure 5A, lane 9). The mRNA detected in cells in G1phase most likely is from cells that did not pass into G1 phase

1534 The Plant Cell
B
D
iLlLlli
Figure 4. Synchronization of an M. varia Suspension Culture.Flow cytometric analysis of cell synchrony is as follows:(A) Asynchronous cells.(B) Mitotically arrested cells.(C) Cell distribution 24 hr after treatment with aphidicolin.(D) to (I) Cell distribution 5, 8,10,13,16, and 24 hr, respectively, afterrelease from the aphidicolin block.Samples were taken at the indicated time points and analyzed by flowcytometry. The profiles represent the distribution of the cells in differ-ent phases: left peak constitutes cells with a 2C value of DNA content,right peak is 4C cells, and the area between the 2C and 4C peaksrepresents cells with an intermediate DNA content.
but remained in G2 (20% as seen in Figure 41). When the sameset of RNAs was analyzed with a probe against the cycMs2gene, a different pattern was obtained (Figure 5B). Althoughmaximal expression occurred again in M phase (Figure 5B,lane 8), cycMs2 transcript levels appeared later in G2 thanthose for the cycMsl gene. The two cyclin genes attained aboutequal steady states in M phase (Figures 5A, lane 8, and 5B,lane 8). After rehybridization of these mRNAs with several othergenes, only the gene coding for an alfalfa protein phospha-tase 2A homolog (pp2aMs; M. Pirck, A. Pay, H. Hirt, and E.Heberle-Bors, unpublished data) showed real constitutive ex-pression during the cell cycle (Figure 5C). Therefore, thepp2aMs gene was used as a control in all subsequentexperiments.
Growth Phase-Dependent Expression of cycMsl andcycMs2 Transcripts
To test whether the two alfalfa cyclin genes behave differentlyduring the growth cycle of a batch suspension culture, loga-
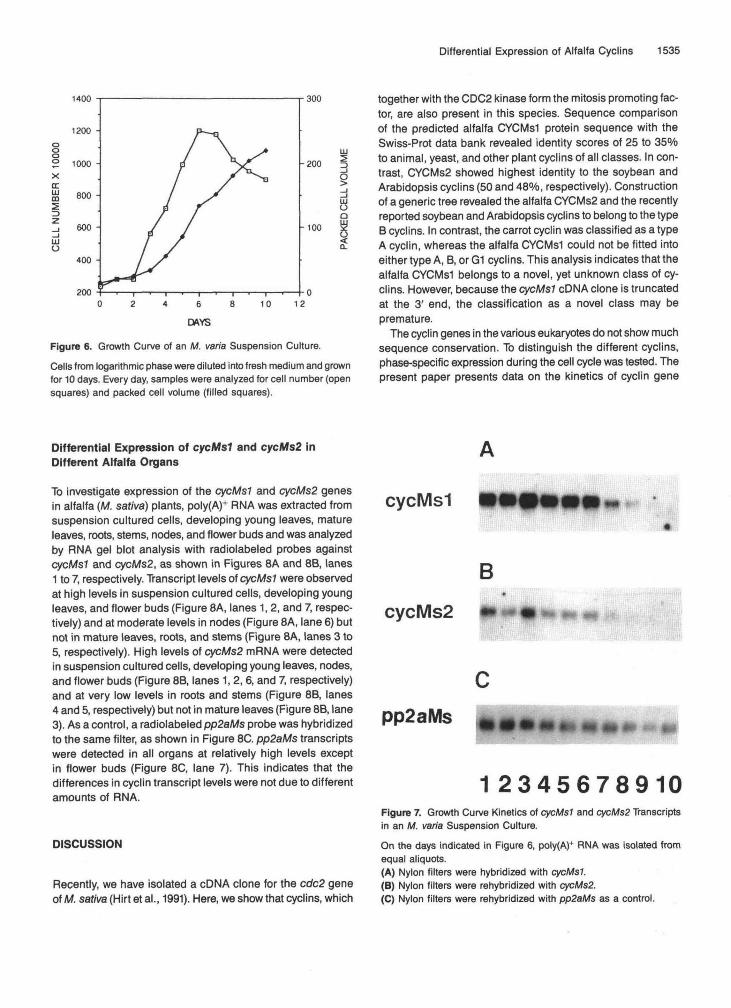
rithmic alfalfa suspension cultured cells (M. varia) were dilutedinto fresh medium and grown for 9 days. After a 2-day lagphase, maximal increase in cell number and packed cell vol-ume was observed during days 3 to 6, as shown in Figure 6.Then, the rate of cell number increase leveled off and the cellsbecame stationary after day 8. Every day during this growthcycle, an aliquot was withdrawn from the culture and analyzedfor cycMsl and cycMs2 transcript levels by RNA gel blot hy-bridization, as shown in Figure 7. During the first 6 days, highlevels of both cyclin transcripts were observed (Figures 7A and7B, lanes 1 to 6, for cycMsl and cycMs2, respectively). There-after, mRNA levels of both genes decreased and were notdetected anymore after day 8 (Figures 7A and 7B, lanes 8 to10, respectively). cycMsl transcript levels were consistentlyhigher than cycMs2 levels. These expression patterns paral-leled the growth kinetics during logarithmic and stationaryphase but not during lag phase. The pp2aMs gene used todemonstrate presence of RNA was expressed throughoutthe growth cycle, but transcript levels decreased gradually(Figure 7C).
cycMsl
asM
ifG1 S G2 G1
B
cycMs2 #• . -HW
pp2aMs
1 2 3 4 5 6 7 8 9Figure 5. Cell Cycle-Dependent Transcription of Alfalfa cycMsl andcycMs2 Genes.Lanes 1 to 9 correspond to profiles (A) to (I) in Figure 4. as, asynchro-nous suspension culture; M, mitotically arrested cells. RNA was isolatedfrom synchronized M. varia suspension cultured cells. Total RNA (100\ig) was poly(A)+ purified and applied to each lane.(A) Nylon filters were hybridized with cycMsl.(B) Nylon filters were rehybridized with cycMs2.(C) Nylon filters were rehybridized with pp2aMs as a control.
Differential Expression of Alfalfa Cyclins 1535
1400
g
300
200
-100
8Q
B
20012
Figure 6. Growth Curve of an M. varia Suspension Culture.Cells from logarithmic phase were diluted into fresh medium and grownfor 10 days. Every day, samples were analyzed for cell number (opensquares) and packed cell volume (filled squares).
together with the CDC2 kinase form the mitosis promoting fac-tor, are also present in this species. Sequence comparisonof the predicted alfalfa CYCMsl protein sequence with theSwiss-Prot data bank revealed identity scores of 25 to 35%to animal, yeast, and other plant cyclins of all classes. In con-trast, CYCMs2 showed highest identity to the soybean andArabidopsis cyclins (50 and 48%, respectively). Constructionof a generic tree revealed the alfalfa CYCMs2 and the recentlyreported soybean and Arabidopsis cyclins to belong to the typeB cyclins. In contrast, the carrot cyclin was classified as a typeA cyclin, whereas the alfalfa CYCMsl could not be fitted intoeither type A, B, or G1 cyclins. This analysis indicates that thealfalfa CYCMsl belongs to a novel, yet unknown class of cy-clins. However, because the cycMsl cDNA clone is truncatedat the 3' end, the classification as a novel class may bepremature.
The cyclin genes in the various eukaryotes do not show muchsequence conservation. To distinguish the different cyclins,phase-specific expression during the cell cycle was tested. Thepresent paper presents data on the kinetics of cyclin gene
Differential Expression of cycMsl and cycMs2 inDifferent Alfalfa Organs
To investigate expression of the cycMsl and cycMs2 genesin alfalfa (M. sativa) plants, poly(A)+ RNA was extracted fromsuspension cultured cells, developing young leaves, matureleaves, roots, stems, nodes, and flower buds and was analyzedby RNA gel blot analysis with radiolabeled probes againstcycMsl and cycMs2, as shown in Figures 8A and 8B, lanes1 to 7, respectively. Transcript levels of cycMsl were observedat high levels in suspension cultured cells, developing youngleaves, and flower buds (Figure 8A, lanes 1,2, and 7, respec-tively) and at moderate levels in nodes (Figure 8A, lane 6) butnot in mature leaves, roots, and stems (Figure 8A, lanes 3 to5, respectively). High levels of cycMs2 mRNA were detectedin suspension cultured cells, developing young leaves, nodes,and flower buds (Figure 8B, lanes 1,2, 6, and 7, respectively)and at very low levels in roots and stems (Figure 8B, lanes4 and 5, respectively) but not in mature leaves (Figure 8B, lane3). As a control, a radiolabeled pp2aMs probe was hybridizedto the same filter, as shown in Figure 8C. pp2aMs transcriptswere detected in all organs at relatively high levels exceptin flower buds (Figure 8C, lane 7). This indicates that thedifferences in cyclin transcript levels were not due to differentamounts of RNA.
DISCUSSION
Recently, we have isolated a cDNA clone for the cdc2 geneof M. sativa (Hirt et al., 1991). Here, we show that cyclins, which
cycMsl
B
cycMs2
pp2aMs
1 2 3 4 5 6 7 8 9 1 0Figure 7. Growth Curve Kinetics of cycMsl and cycMs2 Transcriptsin an M. varia Suspension Culture.On the days indicated in Figure 6, poly(A)+ RNA was isolated fromequal aliquots.(A) Nylon filters were hybridized with cycMsl.(B) Nylon filters were rehybridized with cycMs2.(C) Nylon filters were rehybridized with pp2aMs as a control.

1536 The Plant Cell
cycMs"!
B
cycMs2
pp2aMs
1 2 3 4 5 6 7Figure 8. Expression of Alfalfa cycMsl and cycMs2 Genes in Differ-ent Plant Organs.Poly(A)+ RNA was extracted from M. sativa suspension cultured cells,developing young leaves, mature leaves, roots, stems, nodes, and flowerbuds (lanes 1 to 7, respectively).(A) Nylon filters were hybridized with cycMst(B) Nylon filters were rehybridized with cycMs2.(C) Nylon filters were rehybridized with ppZaMs as a control.
expression during the cell cycle, and cycMsl and cycMs2 tran-scripts accumulate with different kinetics. cycMs2 mRNAappears late in G2 phase and is barely detectable in G1 orS phase. This periodicity is typical for type B cyclins and agreeswith its classification by primary sequence comparison. Weconclude that CYCMs2 is a type B cyclin.
Detectable transcripts of cycMsl were present in all cell cy-cle phases, increased at the onset of G2, and disappearedat the end of mitosis. This kinetics resembles the pattern oftype A cyclins. However, in various organisms, type A cyclinsappear in S phase and also disappear earlier during mitosis(Pines and Hunter, 1989; Whitfield et al., 1989; Lehner andO'Farrell, 1990; and Minshull et al., 1990). We could not detectthis type of periodicity in our experiments. However, when thesame RNAs were probed with cycMsl at low stringency, twomore transcripts appeared with a strikingly similar kinetics asthat reported for type A cyclins. These mRNAs appeared atthe onset of S phase, stayed constant over the entire G2 period,and vanished in M phase slightly before cycMsl and cycMs2transcripts disappeared (data not shown). It appears, there-fore, that plants contain typical type A cyclins. Based on
sequence comparison and expression kinetics during the cellcycle, CYCMsl seems to be a member of a novel class of mi-totic cyclins.
The two cyclin genes were both maximally expressed in G2phase of dividing suspension cultured cells, indicating that bothare mitotic cyclins. Their different periodicity indicates that theymay have different functions.
RNA gel blot analysis showed high transcript levels of cycMsland cycMs2 in those organs that contained meristematic ac-tivity (young leaves, nodes, and flower buds) but not in matureorgans. In young leaves expression levels similar to suspen-sion cultured cells were found, whereas in mature leaves nocycMsl and cycMs2 transcripts could be detected. This demon-strates that in plants as well as in animal cells cyclin expressionis a marker for cell division activity. Also in stationary phaseof batch cultured cells, transcripts of neither cyclin could bedetected.
In suspension cultured cells, about equal amounts of bothcyclin transcripts were observed. In plant organs, however,mRNA levels of the cycMsl and cycMs2 genes showed con-siderable variation. This result could reflect the differentdistribution of the cells in the cell cycle. Alternatively, the twocyclin genes might be expressed in different cells. Future work,particularly in situ hybridization, should help to answer thesequestions.
METHODS
Library Screening and Sequence Analysis
Two degenerate oligonucleotides A (AAA/GTAT/CGAA/GGAA/GAT-ITAT/CCC) and B (ATT/CC/TTIG/ATIGAT/CTGGT/CTIGT) encoding thehighly conserved cyclin boxes K-Y-E-E-M(I)-Y-P and I-L-V(I)-D-W-L(F)-Vwere synthesized. After gel elution from 10% denaturing polyacryl-amide gels, 50 ng of each batch of oligonucleotides was 32P-labeledwith 50 nCi v-ATP and polynucleotide kinase. Approximately 300,000colonies of a cDNA library from an alfalfa (Medicago sativa) cell sus-pension culture (Hirt et al., 1991) were screened with both probes.Putatively positive clones were rescreened to a minimum fit of 16 nucleo-tides according to Wood et al. (1985). The only positive clone cycMsJwas subcloned into pBluescript SK+ (Stratagene) and sequenced fromboth directions with the T7 polymerase sequencing kit from Pharma-cia. Several other M. sativa suspension culture cDNA libraries werescreened with the randomly primed 32P-labeled cycMsl cDNA frag-ment (Feinberg and Vogelstein, 1983). In one case, a 1.4-kb cDNA clonecycMs2 was isolated that was then sequenced as described above.
Cell Culture, Synchronization, and Flow Cytometry
A suspension culture of Medicago varia cells was grown in MS medium(Murashige and Skoog, 1962), containing 0.2 mg/L kinetin and 1 mg/L2,4-dichlorophenoxyacetic acid. Mitotic synchronization was achievedby treatment with aphidicolin. Aphidicolin was added at time zero andafter 12 hr to a final concentration of 20 ng/mL. Cells were then washedtwice with fresh medium and allowed to grow for the indicated times.Flow cytometric analysis was performed as described (Pfosser, 1989).

Differential Expression of Alfalfa Cyclins 1537
Genomic DNA Gel Blot Analysis
Ten micrograms of DNAof M. sativa was digested with Hindlll or Bglll and separated on 0.7°/o agarose gels. After blotting, the nylon filters were hybridized to randomly 32P-labeled fragments of cycMs7 or cycMs2.
RNA Extraction and RNA Gel Blot Analysls
Suspension cultured cells of M. varia or plant tissue of M. sativa was frozen in liquid nitrogen and extracted by grinding in a mortar as de- scribed (Cathala et al., 1983). RNA gel blot analysis was performed according to Gyorgyey et al. (1991). Probes for hybridization were the coding regions of cycMs7 and cycMs2 cDNA clones or the coding re- gion of a phosphoprotein phosphatase 2A cDNA clone from alfalfa as a control. All fragments were 32P-labeled by random priming (Feinberg and Vogelstein, 1983).
ACKNOWLEDGMENTS
This work was supported by Grant No. S 6004 BIO from the dsterreichischer Fonds zur Forderung der Wissenschaften and the Austrian Ministry of Science and Research.
Received August 3, 1992; accepted October 12, 1992.
REFERENCES
Booher, R., and Beach, D. (1987). lnteraction between cdcl3+ and cdc2+ in the control of mitosis in fission yeast; dissociation of the G1 and G2 roles of the cdc2+ protein kinase. EMBO J. 6,
Booher, R., and Beach, D. (1988). lnvolvement of cdcl3+ in mitotic control in Schizosaccharomycespombe: Possible interaction of the gene product with microtubules. EMBO J. 7, 2321-2327.
Cathala, G., Savouret, J.-F., Mendez, E., West, B.L., Karin, M., Martlal, J.A., and Baxter, J.D. (1983). A method for isolation of in- tact, translationally active ribonucleic acid. DNA 2, 329-335.
Colasantl, J., Tyers, M., and Sundaresan, V. (1991). lsolation and characterization of cDNA clones encoding a functional ~34°C' homologuefmnZeamays. Proc. Natl. Acad. Sci. USA88,3377-3381.
Evans, T., Rosenthal, E.T., Youngblom, J., Distel, D., and Hunt, T. (1983). Cyclin: A protein specified by maternal mRNA in sea ur- chin eggs that is destroyed at each cleavage division. Cell 33,
Felnberg, AR, and Vogelstein, B. (1983). A technique for radiolabeling DNA restriction endonuclease fragments to high specific activity. Anal. Biochem. 132, 6-13.
Ferrelra, ECG., Hemerly, A.S., Vlllarroel, R., Van Montagu, M., and Inzb, D. (1991). The Arabidopsis functional homolog of the p34CdC2 protein kinase. Plant Cell 3, 531-540.
3441-3447.
389-396.
Goebl, M., and Byers, B. (1988). Cyclin in fission yeast. Cell 54,
Gytirgyey, J., Gartner, A., Nemeth, K., Magyar, i!., Hirt, H., Heberle- Bors, E., and Dudlts, D. (1991). Alfalfa heat shock genes are differentially expressed during somatic embryogenesis. Plant MOI. Biol. 16, 999-1007.
Hagan, I.M., Hayles, J., and Nurse, P. (1988). Cloning and sequenc- ing of the cyclin related cdc13+ gene and a cytological study of its role in fission yeast mitosis. J. Cell Sci. 91, 587-595.
Hanks, S.K., Quinn, A.M., and Hunter, T. (1988). The protein kinase family: Conserved features and deduced phylogeny of the catalytic domains. Science 241, 42-52.
Hata, S., Kouchi, H., Susuka, I., and Ishii, T. (1991). lsolation and characterization of cDNA clones for plant cyclins. EMBO J. 10,
Hemerly, A., Bergounioux, C., Van Montagu, W., Ind , D., and Ferrelra, P. (1992). Genes regulating the plant cell cycle: lsolation of a mitotic-like cyclin from Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 89, 3295-3299.
Higgins, D.G., and Sharp, RM. (1988). CLUSTAL: A package for per- forming multiple sequence alignments on a microcomputer. Gene
Hirt, H., Pay, A., Gyorgyey, J., Bako, L., Nemeth, K., Bogre, L., Schweyen, R.J., Heberle-Bom, E., and Dudits, D. (1991). Com- plementation of a yeast cell cycle mutant with an alfalfa cDNA encoding a protein kinase homologous to ~ 3 4 ~ ~ ~ ~ . Proc. Natl. Acad. Sci. USA 88, 1636-1640.
Lehner, C.F., and OFarrell, P. (1989). Expression and function of Dro- sophila cyclin A during embryonic cell cycle progression. Cell 56,
Lehner, C.F., and OFarrell, P. (1990). The roles of Drosophila cyclins A and B in mitotic control. Cell 61, 535-547.
Minshull, J., Blow, J.J., and Hunt, T. (1989). Translation of cyclin mRNA is necessary for extracts of activated Xenopus eggs to enter mito- sis. Cell 56, 947-956.
Minshull, J., Golsteyn, R., Hill, C.S., and Hunt, T. (1990). The A- and B-type cyclin associated cdc2 kinases in Xenopus turn on and off at different times in the cell cycle. EMBO J. 9, 2865-2875.
Murashlge, T., and Skoog, F. (1962). A revised medium for rapid growth and bioassays with tobacco tissue cultura Physiol. Plant. 15,473-497.
Murray, A.M., and Kirschner, M.W. (1989). Cyclin synthesis drives the early embryonic cell cycle. Nature 339, 275-280.
Murray, A.M., Solomon, M.J., and Kirschner, M.W. (1989). The role of cyclin synthesis and degradation in the control of maturation pro- moter activity. Nature 339, 280-286.
Nurse, P. (1990). Universal control mechanism regulating onset of M-phase. Nature 344, 503-507.
Ffosser, M. (1989). lmproved method for critical comparison of cell cycle data of asynchronously dividing and synchronized cell cul- tures of Nicotiana tabacum. J. Plant Physiol. 134, 741-745.
Pines, J., and Hunter, T. (1989). lsolation of a human cyclin cDNA: Evidence for cyclin mRNA and protein regulation in the cell cycle and for interaction with ~ 3 4 ~ ~ ~ ~ . Cell 58, 833-846.
Solomon, M., Booher, R., Kirschner, M., and Beach, D. (1988). Cy- clin in fission yeast. Cell 54, 738-739.
Standart, N., Minshull, J., Plnes, J., and Hunt, T. (1987). Cyclin syn- thesis, modification and destruction during meiotic maturation of the starfish oocyte. Dev. Biol. 124, 248-254.
739-740.
2681-2688.
73, 237-244.
957-968.

1538 The Plant Cell
Swenson, K.I., Farrell, K.M., and Ruderman, J.V. (1986). The clam come localized in pole cells during embryogenesis. Nature 338,
Wood, W.I., Gltschier, J., Lasky, L.A., and Lawn, R.M. (1985). Base composition independent hybridization in tetramethylammonium chloride: A method for oligonucleotide screening of highly complex gene libraries. Proc. Natl. Acad. Sci. USA 82, 1585-1588.
embryo protein cyclin A induces entry into M phase and the resump- 337-340. tion of meiosis in Xenopus oocytes. Cell 47, 861-870.
Whitfleld, W.G.F., Gonzalez, C., Sanchez-Herrero, E., and Glover, D.M. (1989). Transcripts of one of two Drosophila cyclin genes be-


















