
alex7.pdf
29
Are Homologies (S-E or CR) Function-Free? Dobzhansky’s dictum, that nothing in biology makes sense except in the light of evolution, by which he meant evolution by Darwinian natural selection, has become almost a mantra among philosophers of biology. But there are some who take the claim a little more seriously than others. Some philosophers of biology hold that many biological predicates (including those employed in comparative anatomy and in making homology judgments) make explicit or implicit appeal to natural selection. Their argument is a simple one: many biological kinds of traits are func tional, and the functions involved are “(naturally) selected effects”. Ergo, many biological kinds of traits are individuated in terms of what Wright called a “consequence etiology.” (Wright, 1976) We have both argued for this claim elsewhere (most recently in Neand er, 2002, Rosenberg, 2006). 1 This claim has been subject to d ispute by those philosophers of biology who endorse the ‘causal role’ account of functions due to Cummins, and by an overlapping group of philosophers of biology who argue that many important biological predicates are defined by homology, and that homology has no need of, or indeed no role for selected-effect functions. Among the most prominent of this overlapping group are Amundson and Lauder (1994) and Griffiths (1994, 2006). In this paper we review the way homology claims figure in the understanding of biological “ characters”, and attempt to vindicate the importance of selected-effect function in the concepts of biology. Some of our argument involves reprising Neande r (2002), and identifying some of the misunderstandings of it in Griffiths (2006). One misunderstanding is best mentioned immediately. According to Griffiths, Neander rejects Functional Minimalism, which Griffiths defines as the view that “disciplines like anatomy, physiology, molecular biology and developmental biology individuate character by homology” (p.3). However, Neander (2002, 395) defines Functional Minimalism as “the view that there are no 1 One of us (Rosenberg) has argued for the thesis that all basic kinds in biology beyond the polynucleotide and the polypeptide sequence are (SE) functionally individuated. Here, however, we argue for the more limited thesis that many of these kinds are, including paradigmatic examples of homologous kinds.
-
Upload
brent-cullen -
Category
Documents
-
view
221 -
download
0
Transcript of alex7.pdf

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 1/29
Are Homologies (S-E or CR) Function-Free?
Dobzhansky’s dictum, that nothing in biology makes sense except in the light of
evolution, by which he meant evolution by Darwinian natural selection, has become
almost a mantra among philosophers of biology. But there are some who take the claim a
little more seriously than others. Some philosophers of biology hold that many biological
predicates (including those employed in comparative anatomy and in making homology
judgments) make explicit or implicit appeal to natural selection. Their argument is a
simple one: many biological kinds of traits are functional, and the functions involved are
“(naturally) selected effects”. Ergo, many biological kinds of traits are individuated in
terms of what Wright called a “consequence etiology.” (Wright, 1976) We have both
argued for this claim elsewhere (most recently in Neander, 2002, Rosenberg, 2006).1
This claim has been subject to dispute by those philosophers of biology who endorse
the ‘causal role’ account of functions due to Cummins, and by an overlapping group of
philosophers of biology who argue that many important biological predicates are defined
by homology, and that homology has no need of, or indeed no role for selected-effect
functions. Among the most prominent of this overlapping group are Amundson and
Lauder (1994) and Griffiths (1994, 2006). In this paper we review the way homology
claims figure in the understanding of biological “characters”, and attempt to vindicate the
importance of selected-effect function in the concepts of biology.
Some of our argument involves reprising Neander (2002), and identifying some of the
misunderstandings of it in Griffiths (2006). One misunderstanding is best mentioned
immediately. According to Griffiths, Neander rejects Functional Minimalism, which
Griffiths defines as the view that “disciplines like anatomy, physiology, molecular
biology and developmental biology individuate character by homology” (p.3). However,
Neander (2002, 395) defines Functional Minimalism as “the view that there are no
1 One of us (Rosenberg) has argued for the thesis that all basic kinds in biology beyond
the polynucleotide and the polypeptide sequence are (SE) functionally individuated.Here, however, we argue for the more limited thesis that many of these kinds are,
including paradigmatic examples of homologous kinds.

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 2/29
2
important functional categories in biology except perhaps for the analogous categories”.
This does not entail the view Griffiths attributes to her.2
Neander’s (2002) argues that homology does not suffice for classification because
claims of homology require a prior specification of the trait in question. This is also one
message of the present paper. Another important take home lesson from this dialectic
between proponents of the indispensability of selected-effects functions and the
exponents of selected-effects free homologies is how complicated and theoretically
intricate (not to say apparently circular, but in reality ‘spiral’) even the most basic
biological descriptions turn out to be. For if we are correct, homology—similarity due to
shared ancestry or common descent— is a property of traits which are often individuated
by their functions in the selected-effect sense, and selected effect traits are traits with a
certain type of ancestry, or that are in other words homologous.3 Both homology and
function, then, are notions shot through with Darwinian presuppositions about ancestry
and adaptation. The difference between them is that homologies don’t wear their
adaptational pedigrees “on their sleeves,” so to speak, while functions do.
1. More than you ever wanted to know about inner ear, jaw and gill bones and
cartilage.
One of the classic examples of a homologous structure is the pentadactyl (= five
digit) limb.
2 Neander (2002) makes this point repeatedly. She says, for instance, “(i)n what follows I
explain that homology is inadequate as a solitary principle of classification and that itrequires assistance from some further complementary principle or principles of
classification (p.392). And then, "I agree that homology is important in the classificationof traits. Where we differ is on the role of function” (p. 392), and “(s)ome homologous
categories might be functional categories too” (p.11), so that “(w)e can stress theimportance of homology all we like and yet still say nothing to any effect against
classification by function. … we can agree that homology is important withoutconcluding that function is thereby unimportant” (p.12). The very title of the paper is,
“Types of Traits: The Importance of Functional Homologues.” Perhaps the repetition ofher message lulled Griffiths to sleep, perchance to dream of a quite different paper, for he
responds to a straw person version of it.3 We assume rather than argue the second part of the conjunction in this paper, but it is
not in dispute and is discussed in Neander (2002, 403-4).
7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 3/29
3
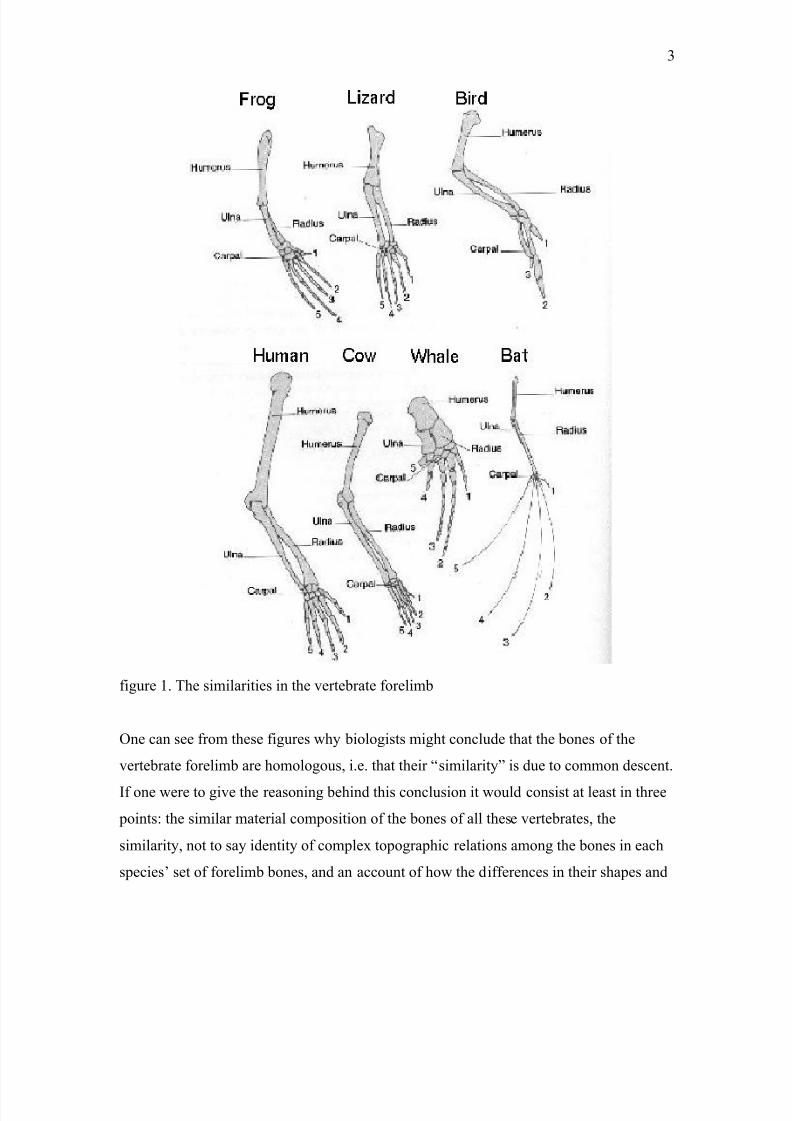
figure 1. The similarities in the vertebrate forelimb
One can see from these figures why biologists might conclude that the bones of the
vertebrate forelimb are homologous, i.e. that their “similarity” is due to common descent.
If one were to give the reasoning behind this conclusion it would consist at least in three
points: the similar material composition of the bones of all these vertebrates, the
similarity, not to say identity of complex topographic relations among the bones in each
species’ set of forelimb bones, and an account of how the differences in their shapes and

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 4/29
4
structure can be expected from the adaptational problems which the forelimb addresses
for each different species.
Homology claims are explanatory hypotheses. They presuppose causes for
similarities. Accordingly, the similarities among traits for which such hypotheses are
advanced are logically prior to homology hypotheses. As Neander (2002, p. 402) put it,
“before two traits can be identified as homologous with respect to each other, we need
some specification of the traits in question.” The traits adjudged similar to one another
cannot, on pain of circularity, be so judged on hypotheses of homology: their similarity
can be explained by their common descent but it cannot merely consist in their common
descent. Otherwise, claims of homology would be tautologous.
Such circularity is avoided in the case of the vertebrate forelimb by the
considerations adduced above: the similarity of the various forelimbs is pretty obvious,
the material composition and structure of the hard parts of the forelimbs--the bones--are
chemically the same, the topographic relations of the bones in each species are highly
similar, and the evident differences among them are to be expected given the
environmental differences within which these sets of bones are to be found at work, so to
speak. The considerable environmental differences between aquatic, avian, swamp,
savannah, steppe and forests to which selection has shaped these forelimbs make a
hypothesis of analogy or homoplasy implausible to say the least.
But such considerations are not always available to ground judgments of
similarity that may be explained by hypotheses of homology. Consider two of the bones
which in the mammal make up parts of the inner ear--the malleus, and the incus.4 These
bones are homologous to two bones, the articular and the quadrate, which in some fish
constitute the jaw hinge. Both of these pairs of bones are homologous to structures that,
in other fish, constitute the gill arch. Three pictures are worth more than three thousand
4 We expound this example at length, since it is employed by Griffiths as presumablyfavorable to his view. Additionally, understanding where Griffiths goes wrong requires a
much more detailed exposition of the case than Griffiths (2006) provides. The example’s prominence in the writing of the opponents of (SE) functional individuation may be due
to Steven J. Gould’s invocation of it. See footnote 4 below.

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 5/29
5
words: The incus and the malleus are two tiny bones better known as the hammer and
anvil:
figure 2. The incus and mellius of the inner ear
The quadrate and the articular in fish jaw skeleton, to which the incus and the malleus are
homologous are labeled ‘q’ and ‘art’ in diagram A of the generic fish jaw hinge below
(diagram B shows how the quadrate and the articulate were attached):

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 6/29
6
figure 3. the quadrate and the articular jaw bones.
The third picture is of a salmon gill arch. This is the boney structure to which the feathery
gill rakers are attached. Fish have several such arches. Early fish had no jawbones and a
series of these gill arches behind their mouths. In these early fish, which lacked bones,
the gill arch was made of cartilage.
figure 4. The gill arches of a Salmon
The homology of these three sets of bones and cartilage have been celebrated among
evolutionary biologists for several reasons, among them is the way in which

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 7/29
7
paleontological evidence about them has both vindicated the fossil record’s support for
the theory of natural selection and undercut the confident claims of creationists that there
are no intermediate forms between these three sets of bones and cartilage.5
These sets of body parts, the inner ear bones of mammals, the jawbone connectors
of fish and the gill-arch cartilage of jawed and jawless fish are plainly not materially,
compositionally the same, nor sufficiently similar in topographic structure to provide a
firm enough basis by themselves for any judgment that they are the same type of body-
part, or similar to one another in respects that require explanation. Surely the
environmental differences between mammals, jawed and jawless fish are not enough to
ground such similarity judgments in spite of the insufficiency of material, structural and
topographic properties as bases for similarity. In fact, there is considerable structural, and
topological evidence against similarity among these traits: mammals have three inner ear
bones, reptiles have only one; reptiles have two other distinct jawbones besides the
articular and the quadrate, while mammals have only one. Why should physically diverse
structures which came to function in hearing and ones that function in biting be similar
5 Gould (1994, 360-1) writes:
The anatomical transition from reptiles to mammals is particularly well
documented in the key anatomical change of jaw articulation to hearing bones.Only one bone, called the dentary, builds the mammalian jaw, while reptiles
retain several small bones in the rear portion of the jaw. We can trace, through alovely sequence of intermediates, the reduction of these small reptilian bones, and
their eventual disappearance or exclusion from the jaw, including the remarkable passage of the reptilian articulation bones into the mammalian middle ear (where
they became our malleus and incus, or hammer and anvil). We have even foundthe transitional form that creationists often proclaim inconceivable in theory —
for how can jawbones become ear bones if intermediaries must live with anunhinged jaw before the new joint forms? The transitional species maintains a
double jaw joint, with both the old articulation of reptiles (quadrate to articular bones) and the new connection of mammals (squamosal to dentary) already in
place! Thus, one joint could be lost, with passage of its bones into the ear, whilethe other articulation continued to guarantee a properly hinged jaw. Still, our
creationist incubi, who would never let facts spoil a favorite argument, refuse toyield, and continue to assert the absence of all transitional forms by ignoring
those that have been found, and continuing to taunt us with admittedly frequentexamples of absence.

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 8/29
8
with ones that function to filter for food and oxygen so that an explanation of their
similarity can be given by common descent? Surely current dissimilarity of function is
no basis for some sort of explanation-demanding similarity. Well, what if these sets of
bones share a similarity in historical function--in a selective etiology?
Here is the biologist’s rough answer to the question of what the similarity to be
explained by homology in this case consists in:
In several of the known intermediates, the bones have overlapping functions, and
one bone can be called both an ear bone and a jaw bone; these bones serve twofunctions…. For example, in Morganucodon, the quadrate (anvil) and the
articular (hammer) serve as mammalian-style ear bones and reptilian jaw bonessimultaneously. In fact, even in modern reptiles the quadrate and articular serve to
transmit sound to the stapes and the inner ear (see Figure 1.4.2). The relevanttransition, then, is a process where the ear bones, initially located in the lower
jaw, become specialized in function by eventually detaching from the lower jawand moving closer to the inner ear.6
The passage makes it plain that a considerable part of the argument for similarity here is
selected effects functional—it is because the different structures can be understood in
terms of a process of adaptation, at times for overlapping roles, that the biologist
concludes that there is a basis for the similarity judgment and also a homology which in
turn explains it.7
There is another potential basis for the similarity judgments about the
incus/malleus pair and the quadrate/articular pair and the gill arches. In embryological
development, the corresponding structures in reptile and mammal fetuses give rise to
these three pairs. So, the pairs share a developmental source. But this shared trait, this
new similarity, raises another question: on what basis does the fetal structure which is the
source of the pair of bones in mammals constitute the same trait-kind as the embryo
structure which is the source of the reptilian pair, or for that matter the same kind of
6 http://www.talkorigins.org/faqs/comdesc/section1.html , May 3, 2007, emphasis
added.
7 Could appeal to ‘function’ in the passage above be an invocation of “causal role” as
opposed to selected effect? Plainly not. Note the evolutionary time scale during which
hearing and chewing functions come apart from one another, and the attribution of thetwo functions to a single set of bones with presumably the same range of proximate
causes and effects (i.e. causal capacities). We return to this matter in section 3 below.

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 9/29
9
structure as the developmental source of the pairs of jaw connectors boney fish have and
the gill arches pairs of cartilaginous fish? The developmental sources of these three pairs
do not bear sufficient similarity of composition, of shape and topological relations, nor do
the differences among these developmental structures reflect obvious etiologies of
adaptation by the same trait-kind to the different past environments of mammals and fish
so as to cry out for an explanation of the similarity that appeals to homology. If they had
been more similar in these obvious respects, comparative embryology would have been a
lot easier, von Baer’s “law” less a subject of controversy one way or the other, and the
tree of life far more obvious a fact than it seems at first blush to be.
Here, at the level of development in a single lineage, similarity depends on
classification by ancestrally shared selected effects. After all, what does it mean to say
that the developmental structures in these diverse orders are the same? Roughly this: the
earliest of them persists over time because it was selected for giving rise to gill arch
cartilage, variation in the same structure was subsequently selected for giving rise to jaw
bone hinges, and further variation in that same structure was selected for giving rise to
inner ear bones in mammals. Notice that nowadays, these “sameness of developmental
structure” claims are being cashed in for sufficient similarity in the types, numbers and
order of homeobox genes, to explain how the same developmental structure gives rise to
diverse developed homologous traits. Similarity in development simply is a matter of
being in the same line of descent, partaking in overlapping parts of a series of
consequence etiologies.
Homology judgments require judgments of similarity, and biological similarity is
cashed in, at least in part, by way of selected effect function. The current example
illustrates why this is so rather clearly. Without an appeal to selected effect function, it is
hard to fathom how three pairs of such manifestly different pairs of traits could be judged
sufficiently similar to have their similarity explained by common descent—i.e.
homology. After all, the three pairs are not materially the same, one of the pairs—the gill
arches, are not even bones, but cartilage; moreover in many species these twin gill arches
do not physically make contact with one another as the jaw bones and inner ear bones do;
and of course the topographic features of all three pairs vary greatly across species, and
indeed differ even within species.

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 10/29
10
Of course, owing to Darwinian gradualism, in the long run and if paleontologists
are fortunate, a range of intermediate forms could be found which makes gradualist
“sense” out of structures radically different in shape, location and topographic locations
(differences great enough to be paraded by creationists). Such a program of research
eventuates in the exhibition of developmental and adult gradients of form, such as those
elaborated by D’Arcy Thompson. But it proceedes on a prior commitment to overlap of
functions among developmental structures that give rise to the diverse instances arranged
in a gradient of changes in form. In the best case, the paleontologists data vindicate the
hypothesis of the comparative developmental anatomist that diverse embryonic structures
across a range of species are instances of the same kind--a kind originally unified by their
functions in embryological development which give rise to homologous adaptations, i.e.
traits with selected effects.
When can a judgment of biological similarity avoid making recourse to selected
effects altogether? There is no general recipe, but a consideration of nucleotide and
amino acid sequence structure shows how limited such cases might be. When a character
can be unambiguously described in nonbiological terms, similarities—indeed identities
can be established free from functional hypotheses. In the simplest case, short sequences
of either nucleotides or amino acids may be molecule-for-molecule structurally identical
(sometimes, perhaps often, without reflecting any biologically interesting—i.e.
functional-- similarity). The longer the linear sequence of a pair of DNA strands or a pair
of polypeptides (proteins) are molecule-for -molecule identical, the greater is the
likelihood that their similarity is biologically significant, and calls for explanation
(usually as homologies, much more rarely as analogies). Even when the match is not
perfect, there may be purely structural, physico-chemical grounds to infer similarity
among such pairs of molecules and seek an explanation for it in terms of homology.
When two somewhat different DNA sequences produce the same protein or similar ones
with the same enzymatic role, there is good reason to conclude that they are homologous.
When differences in gene-sequence do not make a difference to the structure of the
protein they code for, the regions of the two genes which are the same in protein-
sequence coding are said to be “conserved” by selection, and the differences are credited
to neutral point-mutations. This claim is tantamount to a judgment of homology. It is one

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 11/29
11
sort of case, and perhaps the only sort of case, that does not require a prior role for
selected-effect functions to establish the initial similarity or shared kind-hood of the
sequences.8
Thus, when two nucleotide sequences, of sufficient length to constitute regulatory
or structural genes, differ from one another in sequence but produce the same gene
product, the inference to homology is often very firm and yet free from prior assumptions
about selected effects functions. This does not mean that such gene-sequences cannot be
characterized in terms of their selected effect functions. Functional individuation will
closely approximate sequence individuation when single nucleotide differences are
“neutral” substitutions that do not change amino acid codings, or codon differences only
change amino acid sequences far from the enzymatically active or allosteric sites, and are
therefore also selectively neutral. However, almost immediately “above” the level of
primary nucleic acid and amino acid sequence, the multiple realizability of biological
traits grows so rapidly that compositional and topographic differences will make
selected-effects functional identity (of developmental and intermediate structures)
indispensable for character individuation, as in the case of the inner ear bones, the jaw
bones and the gill arches of mammals and fishes.
2. Griffiths, Amundsen and Lauder on homology
In the abstract of “Function, homology, and character individuation”, Griffiths
writes, “I defend the view that many biological categories are defined by homology
against a set of arguments designed to show that all biological categories are defined, at
least in part, by selection function … I show that classification by selection function are
logically dependent on classifications by homology, but not vice versa.” (Griffiths, 2006,
p. 1) And later, “characters are homologies:…they are individuated by common ancestry
or common developmental mechanisms;” “Homology is a relation of biological
sameness.” (2006, p. 5) Amundsen and Lauder (1994/98) concur, excluding selected
functions from the ambit of homological classification: “Whatever the favored definition
of homology, one feature of the concept is crucial: the relation of homology does not
derive from the common function of homologous organs. Organs which are similar in

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 12/29
12
form not by virtue of phylogeny but because of common biological role (or SE function)
are said to be analogous rather than homologous—they have similar SE function and so
evolved to have similar gross structure… The fact that anatomical or morphological
terms typically designate homologies shows that they are not functional categories.”
(Amundson and Lauder, 1994, p. 455.)
Griffiths’ claim is two-fold. (SE) functions are dependent on homology, but not
vice versa. There is no dispute over the first part of his claim: (SE) functions arise when
lineages of traits are selected for certain of their dispositions. Neander (2002, 404) points
out that for this reason her earlier (1991) remarks to the effect that traits are classified by
(SE) function ought never have been interpreted as a denial of the importance of
homology. Since the first part is not in dispute, we focus on the “and not vice versa” part
of Griffith’s claim, which Amundson and Lauder also make.
It is evident that Griffiths’ understanding of homology is at variance with the
biology reported in section 1. As introduced there, homologies are invoked, and
contrasted with analogies, to explain characters already individuated in biological
description, an individuation that often involves thinking about selection history.
Amundson and Lauder evidently must also demur from the biology reported in section 1,
since they too restrict the role of selected effect function to the category of ‘analogy’,
which contradicts the repeated appeal to selected effect function to ground both
anatomical and developmental claims about homology that we have seen above.
To articulate his own and Amundsen and Lauder’s account of the nature of
biological concepts, Griffiths erects a view which he calls “functional revanchism”
which he attributes to Neander:
Functional revanchism: the biological sciences are always at least implicitlyinvestigating function in the selected-function sense.
By contrast, he attributes to Amundson and Lauder and endorses the view that, “unless
anatomy, physiology, molecular biology, developmental biology, and so forth” turn their
attention to specifically evolutionary questions, they investigate function in the causal
sense”(3).
Of course, we do not deny that biologists investigate ‘functions in the causal
sense’, if this means that, among other things, they investigate what traits do, or the

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 13/29
13
adaptive things that traits do. As long as we are not taken to be denying this, which would
be absurd, we agree that ‘functional revanchism’ captures one end of a spectrum of views
which we approach more closely than Amundson, Lauder and Griffiths do. We believe
that the biological sciences very often, at least implicitly, investigate function in the
selected-function sense, even when they turn their attention away from specifically
evolutionary questions.
However, our interest is in the more specific question of how traits are classified.
On this, Griffiths writes, “Neander … thinks that all biological categories are at least
partly defined by selected function” (2006, 3, original italics), and he further attributes to
her the view that biologists “must of necessity supplement considerations of homology
with considerations of selected function” (2006, 13, italics added). In fact, Neander says
something rather more nuanced. She wrote, “I will argue that homologous kinds are
sometimes differentiated in terms of structure, sometimes in terms of function, and
sometimes, and often ideally, in terms of both” (2002, 392, italics added). She even
allows for some classification by ‘functions in the causal sense.’
If ‘functional revanchism,’ is to name a position that the authors of this paper
share, it names the claim that many very important homologous kinds of traits are in part
constituted by selected-effect functions. Griffiths’ view, in startling contrast, is that none
of them are. He sees a “stark contrast” between “the view that biological categories of
parts and processes are defined by their selected function” and the assumption that
“characters are homologies” (Griffiths, 2006, 4). We see no such stark contrast. We claim
that the two are complementary.
On Griffith’s view, homology is more a taxonomic concept than an explanatory
one, providing the basis or criterion for dividing the biological realm at its cladistic joints.
By contrast, if the biology reported in section 1 above is correct, specific claims of
homology are invoked to identify the causes of logically prior classifications-- already
reflecting some shared pedigree.
Griffiths seeks to establish 3 theses about homology as a (SE) function-free basis
of biological classification:
2.1 Homology defines a hierarchy of sets of characters, like taxa, homologous
parts of organisms form groups within groups.

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 14/29
14
2.2 There are levels of homology: Evolution can preserve a morphological
structure while transforming the molecular mechanism that produces it and
conversely can redeploy an existing molecular mechanism to underpin
development of a novelty.
2.3 There are two distinct homology-concepts: a) taxic or Darwinian homology
(which in Griffith 1994 were called ‘cladistic homologies), and b) developmental
homology.
We find his use of 2.1 puzzling, for this is a point that Neander (2002) was
particularly at pains to stress, though Griffiths ignores this fact in his response to her.
Homologous groupings of traits do, to be sure, form hierarchies. Thus larger, more
inclusive groups of homologous characters can be divided into more fine-grained, more
closely related groups in a hierarchical fashion. Neander argued that we therefore “…
have ways of distinguishing smaller homologous groupings within larger homologous
groupings” (2006, 402). She further argued that it followed that homology alone cannot
suffice for classification.9 The fact that classifications of characters are homologous
leaves open the question of what further classificatory principles are involved. Robin
wings are homologous to human arms in some respect or respects (e.g., they are
homologous as vertebrate forelimbs) but they are also homologous to an owl’s wings insome other respect or respects (e.g., as wings). Once we have acknowledged the
importance of homology, we are still left with the task of explaining the logically prior
9 Neander takes this opportunity to disown the argument Griffiths tentatively attributes to
her, when he fixes on a sentence in which she says, “homology is a relation of degree,somewhat akin to the relation of resemblance or genetic relatedness” (Neander, 2002,
402). Perhaps this was an unfortunate way for her to have expressed her point but the page in which it appears makes it fairly clear that she is saying that traits can be more or
less closely homologous, and that just as we must specify the respects in which two itemsresemble each other, we must also specify the respects in which two items are
homologous. Griffiths has her concluding that traits form a ‘continuum’ and that the problem is ‘gradualism’. Neander at one point expresses the view that there will be few
sharp boundaries in what she refers to as ‘the phylogenetic trait tree’, but her argumentdoes not rely on a controversial form of gradualism, and nor should she be read as
claiming, as Griffiths (2006,12) suggests, that homology relations behave formally like ameasure of overall similarity, which is quite contrary to her view. Her argument was the
argument that follows above.

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 15/29
15
principles of classification that are involved. The earlier arguments that Griffiths (1994)
and Amundson and Lauder (1994) offered against (SE) functional categories mostly
consisted in arguments for the importance of homology, as if this by itself could show
that (SE) functions were unimportant. However, this is clearly not so.
Perhaps an even stronger claim can be made. If homology is similarity due to
descent then homology is an explanatory concept, for homologies will explain the
correctness of cladistic taxonomies in the way that atomic theory explains the correctness
of the Periodic Table of the Elements. Griffiths wants to treat homologies in the way
Mendeleev treated chemical reactivities, as diagnostic, not explanatory grounds for
classification in his period table of the elements. There is of course no non-controversial
analytic/synthetic, or classificatory/explanatory distinction, but there is a difference and
Griffith does not accept that homology lies on the explanatory side of biology. He treats
‘homology’ as the basement level classificatory scheme in biology.
Note that the hierarchy of homologies means that if selected effect functions are
required to establish similarity at lower levels of the hierarchy, as was illustrated in
section 1, their roles in determining similarity will continue to be carried by higher-level
homologies even when they are invisible at these higher level.
We agree that 2.2 is a correct observation. However, it reflects the degree to
which homology attributions are explanatory and rely on a great deal of biological theory.
2.2 makes it plain that judgments of developmental homology cannot always rely on
uncontroversial purely structural or non-(SE) functional characterizations of nucleotide or
protein product molecular sequence. Rather, they presuppose theoretical judgments of
similarity that are likely to be based on or qualified by (SE) functional analyses of gene-
action. How, after all, can an underlying molecular developmental pathway be judged to
remain the same pathway to the same morphological structure, during a substantial or
even largely complete turnover of its nucleotide sequence foundation, unless the genetic
basis and the developmental pathway’s identity is fixed by their (SE) functions? Shades
of the ship of Theseus!
Amundson and Lauder (1994) allege that the (SE) function-free homologous
kinds they advocate are “more observational” and “less inferential” than those that rely
on (SE) function. But, as Neander (2002) argued, and as we showed in section 1, claims

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 16/29
16
of homology are highly inferential, and 2.2 gives us further reason to reject their
allegation.10
This also has implications for 2.3’s distinction between what Griffith calls
taxic or Darwinian homology claims and developmental homologies. As in the case
described in section 1, when similarities are unobvious, taxic homology judgments will
require developmental homology grounds. Indeed, there is good reason to hold that
developmental homology judgments will completely undermine taxic homology claims
when they conflict! If two topographically, structurally or topologically similar traits can
be shown to emerge developmentally from distinct pathways that begin at diverse genetic
loci (not themselves homologous through duplication events for example), it is hard to
see what reason there is for treating them as homologies instead of analogies. This
evidential primacy of developmental homology reflects the special and ubiquitous role of
the genome (including RNA and DNA) in carrying information about traits (a claim
which Griffith has elsewhere rejected11
).
Griffiths concludes:
What then is homology? It is a manifest fact that the same parts and processes can
be found in different organisms and in different places in one organism, just as itis a manifest fact that organisms form species. In the early 19th century, biologists
started to develop powerful operational methods for identifying these parts and processes and that research tradition has ever since provided the basis for the
investigation of structure and (causal) function--‘the hierarchical basis ofcomparative biology’ (Hall, 1994). So homology, like the existence of species, is
a phenomenon that stands in need of explanation.
These are not obvious and anodyne observations. We dissent from them, owing to a
fundamentally different appreciation of the role of biological theory in classification. To
begin with, it is not a manifest fact, but an inferred conclusion that the same part- and
processes- kinds or types can be found in different organisms. For, as we have seen,
sameness of parts is a theoretically tendentious conclusion, one that requires use of
considerable theoretical apparatus. Homology and its sister concept, analogy, do not
describe explananda--phenomena to be explained. They provide explanantia--different
10 Their claim that SE (function) free classifications were more observational and less
inferential lead Neander (2002, 409-10) to wonder if they saw no constitutive role evenfor homology.11
E.g. in Griffiths and Grey, 1994.

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 17/29
17
explanations for similarities which biologists’ operational measures have uncovered since
the 19th
century.
3. What’s wrong with “functionalist revanchism”?
This recall, is Griffith’s name for the thesis that all biology, including
experimental biology, is always at least implicitly investigating function in the selected-
function sense. The reason Griffiths thinks it is false is that, as he believes he has shown,
and as Amundsen and Lauder before him argued, except when it comes to evolutionary
biology (when they concern themselves with analogy or homoplasy), biologists almost
never classify in terms defined by selected effect functions. To dispose of functional
revanchism, once and for all, Griffiths addresses several arguments he attributes to
Neander (2002).
“Functional revanchism” begins with the observation made frequently so far in
this paper, as Neander (2002, p. 402) expresses it and Griffith quotes it (Griffith, 2006, p.
12), “…before two traits can be identified as homologous with respect to each other, we
need some specification of the traits in question.” Neander (2002, 401-2) argued that
cladistic divisions alone cannot serve to classify traits, because clades do not allow us to
draw the required biologically significant distinctions among traits that are within
clades.12
Consider the clade that starts somewhere among the fish when gill arches appear
and goes on to include fish with jaw bones and mammals with inner ear bones. Selected
effect functional characterizations, in contrast, do allow us to distinguish inner ears, and
jawbones from gill arches. They thus provide the resources that experimental or
evolutionary biology requires to make obvious distinctions between traits of mammals,
boney fish and cartilaginous ones. Whence the capacity of selected effect functions to
carve nature’s joints more finely.
12 Griffiths (1994, p. 212) claimed that, “A homologous trait is a character that unites aclade.” Griffiths (2006, section 3) says: “A clade is a taxon which contains an ancestral
species and all its descendant species and this was an attempt to capture the notion oftaxic homology, which I then referred (in 1994) to as ‘cladistic homology’. My (1994)
definition is inadequate because it takes no account of the approaches to homologydescribed above (developmental and serial homology)…” As we have seen, sensitivity to
developmental homology makes Griffiths’ problem more rather than less serious.

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 18/29
18
Griffiths’ response is interesting and revealing for it turns on a counterexample
from molecular biology, where similarities and differences are easy to establish
chemically, without appeal to function, just as noted in section 1.
In cladistics, homologies are inferred from a set of measured similarities betweenorganisms, known as shared characteristics (not ‘characters’--a term Griffith
reserves for homologous traits). Suppose we want to construct a clad gram usingsequence data from a suitable molecule, such as 28S ribosomal RNA. The aligned
sequences from any two species will be identical at some points and different atothers. When they are identical, this shared character (sic) may be a homology or
it may be a homoplasy…(W)hen we say that the character state of the firstnucleotide in each sequence is C(ytosine) we are giving a physical specification…
(2006, p. 12)
Thus, Griffiths concludes, we can specify the ‘shared characteristics’ that judgments of
homology presuppose without invoking selected effects functions. But what Griffiths
needs is a counterexample from a level of organization where there is no scope for
recourse to physics and chemistry to establish uncontroversial chemical identity or
similarity. If homologies were called upon to explain only similarities in molecular
sequence, we could certainly dispense with (SE) functions. But Griffiths is perhaps the
last philosopher to make this concession. He not only rejects the claim that genes can be
individuated by their molecular sequences13
, and rejects the centrality of the gene in
determining biological traits –characters— but as he notes in his elaboration of thesis 2.2,
cladistic or taxic homology will sometimes remain the same under major changes in
underlying molecular mechanisms! Further, since Griffiths makes the remarkably strong
claim that selected-effect function is never involved in trait classification, outside of the
analogous categories, it is incumbent on him to show that this stability of homologous
kinds never rests on the stability in (SE) functions. Our position is a pluralist position; in
many cases it does so rest, though in some cases, as in the nucleotide sequences, it does
not.
So, how do we draw distinctions between biologically important characters, such
as inner ears, jaw hinges, and gill-rake supporters within individual clades? Neander’s
(2002, p. 403) suggestion is that “One main way in which this is done is by drawing
13 (Griffiths and Neumann-Held (1999)

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 19/29
19
conceptual lines at those places where there is significant change in what there was
selection for.” She also points out that there is selection for both dispositions (selected
effects, as it were) and selection of the structures and mechanisms that are responsible for
them, and argues that these two sides of the same coin have complementary classificatory
roles. Selection for supporting gill rakes creates the gill arch, selection for biting changes
the gill arch into the jaw hinge, and selection for hearing moves bones which used to
hinge the jaw away from the jaw so they can play a role in hearing. In our view, the
different stages of this selection history provide a particularly fruitful basis for
differentiating among the traits involved.
What is wrong with this claim, according to Grififths? First, that the three pairs of
bone and cartilage are not defined in terms of their selected function “is evident from the
usual theories about their evolution.” It is unclear what Griffiths has in mind in making
this claim, but recall that functional revanchism is not the view that (SE) functions are the
invariable and sole basis of classification. He continues with what might be a second or a
supporting point: in some reptiles one pair of bones sometimes discharges both functions
of jaw hinge and hearing. Why this should be thought a problem for “functional
revanchism” is obscure, since functional revanchism does not entail that each trait has or
is individuated by a single function, only that distinctions among trait types often track
changes in (SE) function: a change from selection for one function (jaw hinge) to
selection for two functions (jaw hinge and hearing) is such a change. Further, as noted
above, this fact to which Griffiths appeals is one of the most powerful arguments for the
claim that reasoning to homologies rest on prior attributions of function. After all, the
topographic and compositional differences between inner ear bones, jaw hinges and gill-
arch cartilage are so great as to make (SE) functional considerations indispensable to
judgments that they constitute instances of a single, more inclusive kind, and therefore
require a homology explanation.
In the light of the biology of section 1, Griffiths’ third argument really sounds like
a strong consideration in favor of “functional revanchism:”
The transformation of the quadrate into the incus may have been driven byincreased selection for hearing in early, nocturnal mammals, but what creates an
obvious break at this point in the series is not a change of function but traditional
morphological criteria--the quadrate bone in all other osteichtyans (boney fish)

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 20/29
20
have no obvious resemblance to the incus that we see in mammals. Therealization that the incus is a modified quadrate was the result of the comparative
anatomists looking for a way to bring their descriptions of different vertebrateskeletons under a single, general account of the structure of the vertebrate
skeleton (emphasis added, working 15)
Of course, Neander’s (2002) “functional revanchism” has a place for morphological
criteria. Of greater immediate importance, however, is how nicely this paragraph
illustrates the need for and role of prior selected effects classification in comparative
anatomy. As Griffiths indicates, the quadrate bone of contemporary fish has little
resemblance to the mammalian incus, other than that both are evolutionarily derived from
a single type of structure, a type which discharged two distinct functions. In making their
apparently counterintuitive claim that the incus is a modified quadrate so as to make
Darwinian sense out of the very different vertebrate skeletons (it was not anything like as
easy as the vertebrate forelimb), the comparative anatomists relied on the fact that
differentiation of function could drive a single structure into two different ones that
looked nothing like one another even though they are homologous.
Neander (2002) alluded to the manifestly evident fact that many experimental
biologists, for example physiologists studying muscle fibers, persistently employ
functional vocabulary when differentiating among traits. Griffiths seeks to dispose of this
evidence for functional revanchism by appeal to causal-role functions: “The problem
with this appeal to practice is that it equivocates on two senses of the word ‘function’:
actual causal roles (Cummins functions) and selected effects,” he says. The former, he
insists, plays an important role in classification. He continues, “There is a sense in which
this reply is unfair to Neander, since she believes that all references to function (and
indeed structure) are implicitly references to adaptive function” (2006, p. 17). And a
moment later, “not marking this distinction causes her to misunderstand Amundson and
Lauder” (Griffiths, 2006, 17).
This allegation is not accurate. One of Neander’s (2002) criticisms of Amundson
and Lauder’s (1994) views is that they seem to eschew even the causal-role conception of
function in classification. They say that, “we could identify all the members of the
category ‘kidney’ by morphological criteria alone…so in that sense, ‘kidney’ is not a
functional category or at least not essentially a functional category” (Amundson and

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 21/29
21
Lauder, 95/98, p. 456), and elsewhere, “even a severely malformed heart, completely
incapable of pumping blood (or serving any biological role at all) could be identified as a
heart by histological examination” (Amundson and Lauder, 95/98, p. 456). Although they
speak of biological investigation into causal-role functions, one can search their paper in
vain for any description of its role in classification. Moreover, Neander (2002, 393)
explicitly marked the distinction between (SE) functions and causal role functions. She
prefaced her whole paper with what she asserted to be a shared assumption: that if
functions were implicated in abnormality inclusive categories, then they were (SE)
functions. This assumption turns out to have been not shared at all. But on Neander’s
(2002) reading, neither Amundson and Lauder (1994) nor Griffiths (1994) had seemed to
concede any classificatory role to functions of either kind (SE or CR) outside of the
analogous categories.14 Neander’s dual aims were thus to show (a) that functions of some
kind were involved, and (b) that past selection history was also involved.15
Reaching back to earlier discussions of this issue, the dialectical ground shifts
again. For example, Neander’s (1991) brief remarks to the effect that (SE) functions were
necessary for classifying traits was directed against the idea that Cummins-style causal-
role functions could suffice at a time when the latter were lauded as an ahistorical
alternative to SE functions, and one that many (particularly in philosophy of mind, but
also in philosophy of biology) therefore regarded as preferable on methological
14 Neander took Griffiths (1994: 213-14) at his word when he concluded, “(I)f functional
classifications are to be of value to biology it must be because of their superior
generality—the fact that they unite disjunctions of cladistic homologues.” Griffiths mayhave meant to speak of only selected effect functional classifications but he did not say
so. Nor did he mention any role for either kind of function in homologous classificationsin the text leading to this conclusion. This is a surprising omission if he had meant that
they were not (SE) functional but were instead causal-role functional. The truth is thatGriffiths (1994) does not mark the distinction, making Neander’s interpretation
understandable. In any case, Neander was responding to a number of authors, not justGriffiths, in whom she detected symptoms of the view that function (unqualified) was not
required for trait classification.15 The discussion of muscle fibers was used to address (a), in a section concerned with
the related roles of structure versus function (Neander, 2002, 405-409. Of the musclefiber case, Neander (2002, 408) rhetorically asks just this question, “which kinds of
criteria –the functional or the structural—will become constitutive in this case?” Sheanswers, “But this sets up a false dichotomy. The two are intimately interwoven and are
more or less two aspects of the same thing.”

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 22/29
22
(particularly Methological Solipsist) grounds. When exchanges take place over decades,
we should try to keep the changing context in mind.
But at least Griffiths (2006), and, according to him, also Amundson and Lauder,
agree with “functional revanchists” on this: that functional considerations of some kind
are important in the classification of traits, and not only in the case of the analogous ones,
and that these classifications might moreover be historical. The remaining issue then is
whether selected-effects functions are needed, or if only causal-role functions combined
with homology are. The gap between opposing views has shrunk, and it will now need to
be more delicately defined than before. What exactly the difference comes down to is not
entirely clear. For one thing, Griffiths does not define ‘causal function’, but instead offers
an example that is open to interpretation, though he does reference Cummins, 1975.16
Now, an (SE) function depends on there being a lineage of traits which has
undergone selection for a Cummins’ style causal-role function, give or take a few niceties
(e.g., Cummins’ causal-role functions are essentially interest-relative, a feature we do not
endorse, but which has played no role in the arguments offered on either side). Assume
again that Griffiths is willing to combine causal functions with history, in the form of
homology. Two further questions of clarification arise: Does he also agree that selection
is important in classifying traits? And does he also agree that past selection (i.e., of the
traits being classified) is? If he concedes the former, the gap shrinks further. If he
concedes the latter, there is no gap between his view and ours. So we will assume that he
does not concede at least this, on pain of collapsing the distinction between the two
views, and turning the remaining dispute into a merely terminological one.
What is at issue in that case is the role of selection history in trait classification.
We maintain that many homologous types of traits are types of adaptations, whereas
Griffiths is obliged to deny it.
16 Griffiths (2002, 2) contrasts SE functions and causal role functions as follows:“Selected function— e.g., a sequence of nucleotides GAU has the selected function of
coding for aspartic acid if one reason that sequence evolved by natural selection was because it had the effect of inserting that amino acid into some polypeptide in ancestral
organisms. Causal function—e.g., a sequence of nucleotides GAU has the causal rolefunction of coding for aspartic acid if that sequence has the effect of inserting that amino
acid into some polypeptide in the organism in which it occurs”.

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 23/29
23
Note, in making the claim that adaptation is important in homologous trait
classification,we are not denying the importance of other things. In contrast, the
proponents of the opposite view deny that adaptation has any role in trait classification,
outside of the analogous categories. Why take this extraordinary view? Why would
biologists, who are after all so very concerned with adaptation and adaptations (among
other things) not want a taxonomy of adaptations? We are not aware that any argument
has been offered to persuade us that selection history is not present and operative in
constituting categories of traits. As was remarked above, the arguments that were
originally offered, when the first sallies among the present parties were sent forth by
Griffiths (1994) and Amundson and Lauder (1994), were merely arguments to the effect
that most interesting biological classifications were homologous, as if this precluded their
being functional, or at least (SE) functional. But their status as homologies leaves the role
of selection history undecided, as we have seen. In contrast, we have offered a number of
arguments for the classificatory indispensibility of selection history.
One powerful consideration seems almost too patently obvious to need stating. It
is that the dispositions for which there has been selection, along with the structures and
mechanisms that are responsible for these dispositions, are of great biological
significance. If classifications are sensitive to the kinds of similarities and differences
among traits that mattered to selection, they will be sensitive to many of the kinds of
similarities and differences that are important to biological theory, whether it be
evolutionary, ecological, morphological, physiological or psychological. More esoteric
arguments are hardly needed. The task of classification is to carve nature at its most
theoretically important joints, not just at any old joints, for there are vastly too many of
the latter, especially in the biological sciences. One main way biologists do this, we claim
(following, Neander, 2002), is by carving at those places where there has been significant
change in what there has been selection for.
A second consideration is less obvious and so we have taken some pains in this
paper to further articulate it. Not only does homology require prior specification of the
trait in question, an inference to homology also often requires prior reasoning about
adaptive pathways. It is often because a certain sequence in a proposed lineage makes
adaptive sense that an inference to homology is justified.

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 24/29
24
A third consideration still turns on the significance of what Neander (2002) calls
abnormality inclusive categories. Since the dialectical ground has shifted, the argument
needs some recalibration. So let us reconstruct it here. We have argued that similarity
judgments are prior to homology claims: a resemblance in two separate species is
regarded as homologous if it is derived from a common ancestor. As we know, traits are
not homologous simpliciter . They require some specification of the resemblance in
question. Something is homologous to something else as a forelimb, or as a wing , or as
an osephegeal valve, for instance. But how do such italicized phrases tell us which
respect of resemblance is involved? Suppose that two traits are homologous as wings. Is
there, then, some resemblance that wings share in virtue of being wings? Or suppose that
two traits are homologous as osephageal valves. Is there some resemblance that
osephegeal valves share in virtue of being osephegeal valves? We do not assume that
there must be a neat set of necessary and sufficient conditions, but there had better be
something to say about it. We must then ask: What is a wing? What is an osephegeal
valve? But this is, no less, and as we said above, a question about how traits are
classified. This is the first step of the argument. Now for the second step.
One answer is, to say the least, very tempting here. Whether something is a wing
or is an osephegeal valve is, in part at least, a matter of its (SE) function. But now
suppose that someone tells us at this point that there is no need for (SE) functions in
classifying these traits because causal-role function and homology suffice. Of course,
causal-role functions on their own do not suffice because the classifications in question
are abnormality inclusive. Causal role functions are actual causal dispositions, but some
wings and some osephegeal glands are abnormal and consequently lack the causal
dispositions that are characteristic of their types. Some wings can and some cannot assist
in flight. And some osephegeal glands can and some cannot help move food to the gut.
However, the suggestion is no longer that causal-role function on its own suffices for the
classification of traits. The suggestion, from those who object to SE functional
classification, is that causal-role can be combined with homology for these purposes. But
how does homology help at this point? What resemblance is thus shared by common
descent? How is this supposed to work exactly?

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 25/29
25
We further believe that, when we speak of two traits in two separate species as
being homologous, we are implicitly speaking of them being homologous with respect to
their normal structural features and capacities. This remains implicit rather than explicit
because when we speak of traits being homologous we are usually speaking in general
terms, of some traits in general, and not of their particular instances. We are, for example,
speaking of human and chimpanzee esophageal valves, and not specifically of some
individual human’s (Bob’s) and of some individual chimpanzee’s (Bonzo’s). However, if
the esophageal valves of Bob and Bonzo’s respective species are homologous, then by
extension their individual instances are too. They have inherited their esophageal valves
from shared ancestors. But this relation of homology relates to certain of their normal
features, those for which there was selection, and not to any abnormal features they might
happen to have. Acid reflux can damage an esophageal valve, and prevent it from
performing its (SE) function. If this happened to both Bob and Bonzo, their damaged
esophageal glands could still be considered homologous to each other with respect to
certain of their normal structural features and capacities (to the extent that they still
instantiate them) but they are obviously not homologous to each other with respect to
their abnormal features and impaired capacities.
Of course, congenital impairments can be inherited, and so there are lineages of
such ‘traits’ as (e.g.) the hemophilia that plagued the royal families of Europe for a few
hundred years. Perhaps someone might want to say that the hemophilia of one royal male
was homologous to the hemophilia of another. We see no great conceptual impropriety in
speaking this way, but it is not typical homology-speak. Nor, to repeat, do we claim that
typical homology-speak always refers to adaptations. We do not deny, for instance, that
some ‘spandrels’ are homologous to others. Ours is a pluralist position. We maintain that
many, not all, homologous types of traits are adaptations.
It is true that SE functions are possessed by traits in virtue of their lineage. An
instance of a trait has an SE function because it is the product of lineages of genes and
other developmental mechanisms that have been selected for producing it for that reason
(i.e., for performing that function). As we said in the introduction to the present paper,
this discussion reveals just how complex even the most basic seeming biological
descriptions turn out to be. If we are correct, homology, construed as similarity due to

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 26/29
26
shared ancestry or common descent, is a property of traits which are often individuated
by their functions in the selected-effect sense, and selected effect traits are traits with a
certain type of ancestry, or that are in other words homologous.
The similarity judgments, and the intra-species trait classifications, that are prior
to inter-species homology claims will have to bring together both normal , properly
working, and abnormal , defective and diseased instances of structures that vary along
many topographic, morphological and compositional dimensions. As Darwin recognized,
variation is the rule and not the exception, it has many sources and most variations are
deleterious. Such deleterious effects have causal-role consequences. What they retain, in
the face of their abnormal causal-roles and abnormal morphology, and sometimes in the
face of their abnormal development, is a shared selection history. The role of a natural
selection etiology in drawing the normal/abnormal distinction needed for correct
judgments of similarity makes such judgments ‘essentially historical’ in Neander’s terms.
To classify something as either biologically normal or abnormal –by the by, a
mode of classification in itself— requires that there be a standard of biological normality,
and this is given, not by a statistical distribution, but by the solution to the design
problem which created selection pressure on the lineage within which it figures.17
Biologists, including and, perhaps, especially those concerned with questions of
physiology, and certainly not only those concerned with evolutionary theory, have a great
many classifications of traits that implicitly or explicitly appeal to this distinction.
Griffiths grants the importance of abnormality inclusive categories. However, he
counters, first, that “even if abnormality inclusive categories have to be ‘essentially
historical’ this is no argument against their being defined by homology.” Of course, this
is something that Neander (2002) did not deny, as noted in the introduction (and ftn.2).
The question is what else is involved, because homology alone cannot suffice for
classification. But there is a much more consequential oversight, according to Griffiths’
second response to the claim that abnormality inclusive categories are ‘essentially
historical’ in Neander’s sense. To show this he invokes the notion of developmental
homology: “The developmental approach to homology yields abnormality inclusive
categories which are not essentially historical. This should come as no surprise. It would
17 We take it that this is not in dispute by Griffiths, although it is disputed by some.

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 27/29
27
be puzzling if an approach designed to identify characters across evolutionary
transformations could not identify them across perturbing causes such as diseases process
… Neander’s ‘master argument’ fails, and the need for abnormality inclusive categories
can be met using homology.” (2006, p. 18)
Griffiths’appeal to developmental homology cannot show that homology is
abnormality inclusive without presupposing selected effects, unless developmental
homologies themselves are innocent of any selective effect presupposition. That they do
often have such presuppositions is plainly obvious. For one, there is the issue of abnormal
development, and thus of what counts as an instance of the same developmental
homology. For another, recall the role in developmental homology of SE functions to
ground the taxic homologies of inner ear, jaw-bone joint and gill arch expounded in
section 1. The crucial point there was that, as with taxic homologies, before a claim of
homology can be made for a type of developmental structure, we need criteria of
similarity to bring together a diverse set of topographies, compositions, and structures, as
its instances. The kind in question is often (usually) constituted by the developmental
structures’ (SE) function— giving rise to diverse adapted characters.
4. Conclusion
Biology encompasses many different fields of specialization and there will be no
one-size-fits-all formula for taxonomizing biological traits. We have therefore exercised
some caution when it comes to making universal generalizations about how trait
classifications are constituted. However, we continue to be thoroughly impressed by the
truth of Dobzhansky’s dictum, that nothing in biology makes sense except in the light of
evolution, and more specifically in the light of Darwinian natural selection. Natural
selection, we all agree, sheds abundant light on how traits evolved. We maintain that it
also sheds light on how traits are classified in theoretically interesting and useful ways.
We conclude –still— that very many types of traits (including those explained as
homologies) are types of adaptations, classified at least in part on the basis of their
selected effect functions.
Objections to this view have taken different forms over the past several decades.
In this paper we defend it against those who argue that a combination of homology and

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 28/29
28
causal role function can achieve the goals of biological classification, in the absence of
considerations of past selection history. Even if this claim were true, it would not show
that our view was false, in so far as we claim that very many types of traits are
(constitutively) types of adaptations. It would at most only show that they did not need to
be, in which case the parties to the debate about the nature of such classifications could
call it a draw, at least for this round. However, we have shown here that a combination of
homology and causal role function, when divorced from considerations of past selection
history, can not achieve the same classificatory goals in the most favored case of the
opponents of SE functional classification. The judgment that the same trait, or two
similar traits in two separate species is or are homologous usually requires considering a
chain of adaptation. And the claim that the same trait, or two similar traits in two separate
species is or are homologous always requires prior specification of the traits in question,
a specification that cannot –for the abnormality inclusive categories— be given in terms
of mere causal roles, for reasons we have explained.
Karen Neander
Alex Rosenberg
Department of Philosophy &
Center for Philosophy of Biology
Duke University
References
Amundson, Ron and G. V. Lauder (1994) “Function without Purpose: The Uses ofCausal Role Function in Evolutionary Biology,” Biology and Philosophy, 9: 443-469.
Cummins, Robert (1975) “Functional Analysis,” Journal of Philosophy, 72: 741-765.

7/27/2019 alex7.pdf
http://slidepdf.com/reader/full/alex7pdf 29/29
29
Gould, Stephen Jay (1994) “Hooking Leviathan by Its Past”, reprinted in Dinosaur in a Haystack: Reflections in Natural History, (1997) New York: Crown Trade Paperbacks,
359-377.
Griffiths, Paul E. (2004) “Cladistic Classification and Functional Explanation,”
Philosophy of Science, 61: 206-227.
____ (2006) “Function, Homology and Character Individuation,” Philosophy of Science,73: 2006, 1-25.
Griffiths and Grey, (1994) Developmental Systems and Evolutionary Explanation,The Journal of Philosophy, Vol. 91, No. 6, 277-304.
(Griffiths, P. E. and E. Neumann-Held, (1999) “The many faces of the gene,”
BioScience 49(8): 656-662.
Neander, Karen (1991) “Functions as Selected Effects: The Conceptual AnalystsDefense,” Philosophy of Science, 58:168-184.
________ (2002) “Types of Traits: The Importance of Functional Homologues,” inAndre Ariew, Robert Cummins, and Mark Perlman (eds.) Functions: New Essays in thePhilososphy of Psychology and Biology, Oxford, Oxford University Press: 390-415.
Rosenberg, Alex (2006) Darwinian Reductionism, or How to Stop Worrying and Love Molecular Biology, Chicago, University of Chicago Press.
Wright, Larry (1976) Teleological Explanations, Berkeley and Los Angeles, Universityof California Press.


















