
Adrenoleukodystrophy. The chain shortening of erucic acid (22:1(n−9)) and adrenic acid...
5
Biochimica et Biop,hysica Acta, 1002 (1989) 79-83 Elsevier 79 BBA 53048 Adrenoleukodystrophy. The chaff1 shortening of erucic acid (22: 1( n - 9)) and adrenic acid (22: 4( n - 6)) is deficient in neonatal adrenoleukodystrophy and normal in X-linked adrenoleukodistrophy skin fibroblasts Erik Christensen 1, Morten Gmnn ~, Tor-Arne Hagve 1, Bengt Frode Kase 2 and Bjorn O. Christophersen 1 i Institute of Clinical Biochemistry of the University of Oslo and ' Department of Pediatric Research, Rikshospitalet, Osio (Norwm') (Received 13 July 1988) (Revised manuscript received 1 November I )88) Key words: Lignoceryl-CoA ligase; Peroxisomal g-oxidation; Erucic acid; Adrenic acid; X-linked adrenoleukodysuophy: Neonatal adrenoleukodystrophy; (Human fibroblast) The metabolism of long cha|n unsaturate~ fatty acids was studied in cultured fibroblasts from patients with X-linked adrenoleukodystrophy (ALD) an:i with neonatal ALD. By using [14-14C] erucic acid (22: l(n- 9)) as substrate it was shown that the peroxisomal g-oxidation, measured as chain shortening, was impaired in cells from patients with neonatal ALD. The g-oxidation of adrenic acid (22:4(n - 6)), measured as acid-soluble products, was also reduced i'n the neonatal ALD cells. The peroxisomal g-oxidation of [14-14C]erucic acid (22: |(n- 9)) and [2-t4C]adrenic acid (22:4(n-6)) was normal in cells from X-ALD patients° The g-oxidation, esterification and chain elongation of [l-14C]arachidon|c acid (20:4(n- 6)) and [l-14Cleicosapentaenoic acid (20:5(n- 3)) was normal in both X-linked ALD and in neonatal ALD. Previous studies suggest that the activation of very long chain fatty acids by a lignoceryl (2A:0)-CoA figase is deficient in X-linked ALD, while the peroxisomal ft..oxidation enzymes are deficient in neonatal ALD. The present results suggest that the peroxisomal very long-chain acyl-CoA ligase is not required for act~,vation of unsaturated C20 and C22 fatty acids and that these fatty acids can be effidently activated by the long chain acyl-(palmltyi)-CoA ligase. Introduction X-linked adrenoleukodystrophy is an inborn error of metabolism with deficient peroxisomal oxidation of very long chain fatty acids. The biochemicaJ defect is prob- ably localized at a peroxisomal very long chain acyI-CoA ligase (lignoceryl (24:0)-COA ligase) [1,2]. In X-linked adrenoleukodystrophy the three peroxisomal g-oxida- tion enzyme proteins (acyI-CoA oxidase, the bifunc- tional protein with em'~,yI-CoA hydratase and 3-hy- droxyacyl-CoA dehydrogenase activities and 3-oxoacyl- CoA thiolase) are present. All three of these enzyme proteins are deficient in the Zellweger (cerebro-hepato- renal) syndrome where peroxisomes are absen! [3,4]. In neonatal adrenoleukodystrophy the number of per- oxisomes is reduced and the bifunctional enzyme is Correspondence: Erik Chdstensen, Institute of Clinical Biochemistry. Rik~h,~p,italet, N-0027 Oslo 1, Norway. probably absent while acyl-CoA orddase and fl-keto- thiolase are both detectable [5]. In all these peroxisomal diseases very long chain fatty acids accumulate as demonstrated by an increased C26:0/C22:0 ratio in serum and other body fluid.~anti in cultured fibroblasts [6]. The accumulation of yew long chain fatty acids are more pronounced in the Zeliweger syndrome and in neonatal adrenoleukodystrophy than in X-linked adrenoleukodystropiiy [7,8]. We nave recently demonstrated a deficient chain shortening of erucic acid (22 : l(n - 9)) and adrenic acid (22:4(n-6)) in cultured fibroblasts from Zellweger patients [91. The purpose of the present work was to study the chain shortening of C22 unsaturated fatty acids in X-lin- I~,d adrenoleukodystrophy and neonat~[ adrenoleuko- a,strophy. The chain shortening of (-22 unsaturated fatty acids was also studied in fibroblasts from a patient with rhizomelic chondrodysplasia punctata, a per- oxisomal disease where the oxidation of very long chain 0005-2760/89/$03.50 © 1989 Elsevier Science Publishers B.V. ,(Biomedical Division)
-
Upload
erik-christensen -
Category
Documents
-
view
215 -
download
0
Transcript of Adrenoleukodystrophy. The chain shortening of erucic acid (22:1(n−9)) and adrenic acid...

Biochimica et Biop,hysica Acta, 1002 (1989) 79-83 Elsevier
79
BBA 53048
Adrenoleukodystrophy. The chaff1 shortening of erucic acid (22: 1( n - 9)) and adrenic acid (22: 4( n - 6)) is deficient
in neonatal adrenoleukodystrophy and normal in X-linked adrenoleukodistrophy skin fibroblasts
Erik Christensen 1, Morten G m n n ~, Tor-Arne Hagve 1, Bengt Frode Kase 2 and Bjorn O. Christophersen 1
i Institute of Clinical Biochemistry of the University of Oslo and ' Department of Pediatric Research, Rikshospitalet, Osio (Norwm')
(Received 13 July 1988) (Revised manuscript received 1 November I )88)
Key words: Lignoceryl-CoA ligase; Peroxisomal g-oxidation; Erucic acid; Adrenic acid; X-linked adrenoleukodysuophy: Neonatal adrenoleukodystrophy; (Human fibroblast)
The metabolism of long cha|n unsaturate~ fatty acids was studied in cultured fibroblasts from patients with X-linked adrenoleukodystrophy (ALD) an:i with neonatal ALD. By using [14-14C] erucic acid (22: l ( n - 9)) as substrate it was shown that the peroxisomal g-oxidation, measured as chain shortening, was impaired in cells from patients with neonatal ALD. The g-oxidation of adrenic acid (22:4(n - 6)), measured as acid-soluble products, was also reduced i'n the neonatal ALD cells. The peroxisomal g-oxidation of [14-14C]erucic acid (22: | ( n - 9)) and [2-t4C]adrenic acid ( 2 2 : 4 ( n - 6 ) ) was normal in cells from X-ALD patients° The g-oxidation, esterification and chain elongation of [l-14C]arachidon|c acid ( 2 0 : 4 ( n - 6)) and [l-14Cleicosapentaenoic acid (20 :5 (n - 3)) was normal in both X-linked ALD and in neonatal ALD. Previous studies suggest that the activation of very long chain fatty acids by a lignoceryl (2A:0)-CoA figase is deficient in X-linked ALD, while the peroxisomal ft..oxidation enzymes are deficient in neonatal ALD. The present results suggest that the peroxisomal very long-chain acyl-CoA ligase is not required for act~,vation of unsaturated C20 and C22 fatty acids and that these fatty acids can be effidently activated by the long chain acyl-(palmltyi)-CoA ligase.
Introduction
X-linked adrenoleukodystrophy is an inborn error of metabolism with deficient peroxisomal oxidation of very long chain fatty acids. The biochemicaJ defect is prob- ably localized at a peroxisomal very long chain acyI-CoA ligase (lignoceryl (24:0)-COA ligase) [1,2]. In X-linked adrenoleukodystrophy the three peroxisomal g-oxida- tion enzyme proteins (acyI-CoA oxidase, the bifunc- tional protein with em'~,yI-CoA hydratase and 3-hy- droxyacyl-CoA dehydrogenase activities and 3-oxoacyl- CoA thiolase) are present. All three of these enzyme proteins are deficient in the Zellweger (cerebro-hepato- renal) syndrome where peroxisomes are absen! [3,4]. In neonatal adrenoleukodystrophy the number of per- oxisomes is reduced and the bifunctional enzyme is
Correspondence: Erik Chdstensen, Institute of Clinical Biochemistry. Rik~h,~p,italet, N-0027 Oslo 1, Norway.
probably absent while acyl-CoA orddase and fl-keto- thiolase are both detectable [5].
In all these peroxisomal diseases very long chain fatty acids accumulate as demonstrated by an increased C26:0/C22:0 ratio in serum and other body fluid.~ anti in cultured fibroblasts [6]. The accumulation of yew long chain fatty acids are more pronounced in the Zeliweger syndrome and in neonatal adrenoleukodystrophy than in X-linked adrenoleukodystropiiy [7,8].
We nave recently demonstrated a deficient chain shortening of erucic acid (22 : l(n - 9)) and adrenic acid ( 2 2 : 4 ( n - 6 ) ) in cultured fibroblasts from Zellweger patients [91.
The purpose of the present work was to study the chain shortening of C22 unsaturated fatty acids in X-lin- I~,d adrenoleukodystrophy and neonat~[ adrenoleuko- a,strophy. The chain shortening of (-22 unsaturated fatty acids was also studied in fibroblasts from a patient with rhizomelic chondrodysplasia punctata, a per- oxisomal disease where the oxidation of very long chain
0005-2760/89/$03.50 © 1989 Elsevier Science Publishers B.V. ,(Biomedical Division)

80
fatty acids is normal, but the plasmalogen synthesis is deficient [10].
Materials and Methods
Chemicals [14..t4C]Erucic acid (22: l ( n - 9)) was from Centre
d'Etudes Nucl~ires de Saclay, Gif sur Yvette, France. [1-14 C]Arachidonic acid (20: 4 ( n - 6 ) ) and [1-14 C]eicosapentaenoic acid ( 2 0 : 5 ( n - 3)) were from New England Nuclear, Boston, MA, U.S.A. [2-14C]Adrenic acid (22: ,gn - 6)) and [1-t4C]linolenic acid (18: 3(n - 3)) were from The Radiochemical Center, Amersham, U.K. Eagles medium was obtained from Flow Labora- tories, irvine, U.K. Unlabelled erucic acid and linolenic acid were from Sigma Chemical Co., St. Louis, MO, U.S.A. Unlabelled adrenic acid was from Nu-Chek Prep. F, lysian, MN., U.S.A. 2,6.Ditert-butyl-p-creso! was from Koch-Light, Colnbrook, U.K.
Tissue culture Fibroblasts from two patients with X-linked adren-
oleukodystrophy were kindly supplied by Dr. Ann Moser, Kennedy Institute, John Hopkins University, Baltimore. Fibroblasts from a patient with neonatal adrenoleukodystrophy and a patient with rhizomelic chondrodysplasia punctata were kir, dly supplied by Dr. Ola Skjeldal at our institute. Skin biopsies were also taken from the forearm of two healthy individuals. Cultures of skin fibroblasts were initiated and main- tained in basal Eagle's medium with 17~$ fetal calf serum. Penieilfin (100 U/ml), streptomycin (100 ttg/'ml), amphotedcin (2 U/ml) and L-glutamine (1.5 ~mol/ml) were added to the medium. The cells were cultured at 37°C in 25 cm z flasks containing 2 ml of the medium, and gassed with 5~$ COa in air. All the metabolic studies were done between the eight and the eleventh passage in culture. Microscopically, the fibroblasts from both patients and controls had a normal appearance, and showed a normal speed of 8rowth and cell division.
Incubations •
The incubations were started by adding albumin- bound labelled fatty acids to the medium in the culture flask, with confluent cells 8rowing in monolayer. The final fatty acid concentration was 0.1 raM, with a spe~fic activity of 20 mCi/mmol. The fatty acid to total albumin molar ratio was !.5. Each flask contained 1.5-2.4 m8 of cellular protein in a total incubation volume of 2 ml. The medium consisted of basal Eagle's medium without calf serum.
Analytical methods
The measurements of radioactive CO 2 and radioac- five acid-soluble products were perforated as described by Christiansen et el. [11]. The lipids were extracted by
the method of Folch et al. [12] and separated on silicic acid thin-layer plales (Stahi H +) (hexane/diethylether/ glacial acetic acid, 70:30:1, v /v /v) . Aliquots of the total lipid extracts were transmethylated [13] and analyzed by radio gas chromatography using a Hewlett Packard 5890A connected to an ISOMESS RAGA IM 202b RGC (Raytest Strahlungsmessgefiite, Strauben- h~,rdt, D.D.R.) radioactivity detector with a 1 : 10 outlet splitter. Fatty acid methyl esters were separated at 170°C with 10 min initial hold followed by a pro- grammed temperature increase of 3°C per min to 250°C, using 10~ SP-2340 on Supelcoport 100/120 (Supelco Inc., Bellefonte, PA, U.S.A.). Data acquisition and analysis were achieved by using an IBM PC (IBM), Boca Raton, FL, U.S.A.) and a Tecmar Labmaster (Tecmar Inc., Cleveland, Ohto, U.S.A.) timer, counter and analog to digital converter. Special software was written allowing the IBM PC ~to simultaneously control 2 radio gas chromatographs [14], with storage and in- tegration of the chromatograros. The solutions used for lipid extraction and thin-layer chromatography con- tained 2,6-ditert-butyl-p-cresol (50 rag/l) as an antioxi- dant and the lipid extracts were stored under .~trogen gas in the dark at - 2 0 °C to prevent peroxidation of unsaturated fatty acids. Ceilular protein was determined according to the method of Lowry et al. [15].
Results
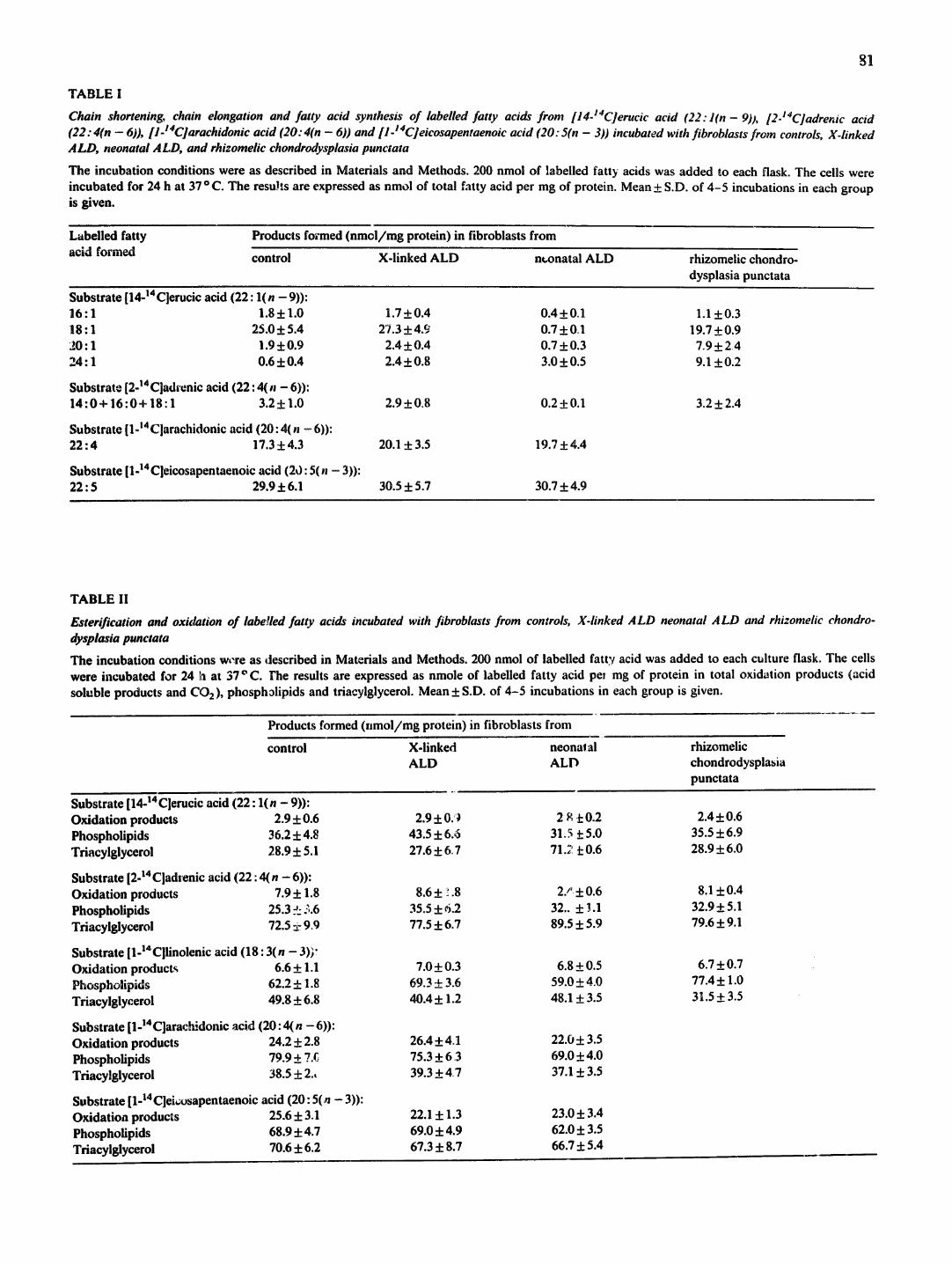
Table I shows that [14-14C]erucic acid (22 : l(n - 9)) is efficiently shortened to labelled oleic acid (18: l ( n - 9)) and smaller amounts of palmitoleic acid (16 : l (n - 9)) and eicosenoic acid (20: l (n - 9)) in fibroblasts from patients with X-linked adrenoleukodystrophy as well as in cel~s from controls.
In fibroblasts from the patient with neonatal adreno- leukodystrophy the chain shortening of 22: l(n - 9) was deficient. A small but significant amount, approx. 1-2~ of the erucic acid substrate was however shortened to labelled 16: l(n - 9), 18: l(n - 9) and 20: l(n - 9) in the fibroblasts from the patient with neonatal adren- oleukodystrophy. In cells from patients with the Zell- weger syndrome the chain shortening of erucic acid was even lower and in some experiments not detectable [9].
Table I also shows that small but significant amounts of labelled newly synthesized fatty acids, 14: 0, 16 : 0 and 18: 1 were formed from [2-14C] 22 : 4 in fibroblasts from the patients with X-linked adrenoleukodystrophy, rhizomelic choadrodysplasia punctata and from con- trols. In fibroblasts from the patients with neonatal adrenoleukodystrophy newly synthesized fatty acids were not detectable or formed in very small amounts.
The chain elongation of the C20 fatty acids arachidonic (2¢~ " ~ n - 6)) and eicosapentaenoic acid (20: 5(n - 3)) v:~s not increased in cells with defects in
81
TABLE I
Chain shortening, chain elongation and fatty acid synthesis of labelled fatty acids from [14-14C]erucic acid (22:1(n-9)) , [2-1~C]adre~ic acid (22: 4(n - 6)), [1-14C]arachidonic acid (20: 4(n - 6)) and [1-14C]eicosapentaenoic acid (20: 5(n - 3)) incubated with fibroblasts from controls, X-linked ALl), neonatal ,4LD, and rhizomelic chondrodysplasia punctata
The incubation conditions were as described in Materials and Methods. 200 nmol of labelled fatty acids was added to each flask. The cells were incubated for 24 h at 37 o C. The results are expressed as nmol of total fatty acid per mg of protein. Mean ± S.D. of 4-5 incubations in each group is given.
L'-belled fatty acid formed
Products formed (nmol /mg protein) in fibroblasts from
control X-linked ALD nt, onatal ALD rhizomelic chondro- dysplasia punctata
S u b s t r a t e [ l ~ 1 4 C ] e r u c i c a c i d ( 2 2 : l ( n - 9 ) ) :
1 6 : 1 1.8±1.0 ~8:1 25.0±5.4 20:1 1.9±0.9 24:1 0.6±0.4
Substrate [2-14Cladrenic acid (22: 4(n - 6)): 1 4 : 0 + 16 :0+ 18:1 3.2± 1.0
Substrate [l-m4Clarachidonic acid (20: 4(n - 6)): 22:4 17.3±4.3
Substrate[1-14Cleicosapentaenoicacid(20:5(n-3)): 22:5 29.9±6.1
1.7±0.4 0.4±0.1 1.1±0.3 27.3±4.9 0.7±OA 19.7±0.9
2.4±0.4 0.7±0.3 7.9±2.4 2.4±0.8 3.0±0.5 9.1±0.2
2.9±0.8 0.2±0.1 3,2±2.4
20.1±3.5 19.7±4.4
30.5±5.7 30.7±4.9
TABLE 11
EMerification and oxidation of labelled fatty acids incubated with fibroblasts from controls, X-linked ALD neonatal ALD and rhizomelic chondro- dysplasia punctata
The incubation conditions w,-re as described in Materials and Methods. 200 nmol of labelled fatt~ acid was added to each culture flask. The cells were incubated for 24 L~ at 37 o C. l'he results are expressed as nmole of labelled fatty acid pe~ mg of protein in total oxidation products (acid soluble products and CO2), phosphalipids and tdacylglycerol. Mean ± S.D. of 4-5 incubations in each group is given.
Products formed (nmoi /mg protein) in fibroblasts from
control X-linked neonatal rhizomelic ALD ALD chondrodysplasia
punctata
Substrate [14-14Clerucic acid (22: l (n - 9 ) ) :
Oxidation products 2.9 ± 0.6 Phospholipids 36.2 ± 4.8 Triacylglycerol 28.9 ± 5.1
Substrate [2-14C]adlenic acid (22:4(n - 6 ) ) :
Oxidation products 7.9 ± 1.8 Phospholipids 25.3 f..~.6 Triacylglycerol 72.5 -~- 9.9
Substrate [l-14C]finolenic acid (18: 3(n - 3))" Oxidation product~ 6.6 ± 1.1 Phospholipids 62.2 ± 1.8 Triacylglycerol 49.8 ± 6.8
Substrate [1-14 C]araclfidonic acid (20: 4( n - 6)): Oxidation products 24.2 ± 2.8 Phospholipids 79.9 ± 7.C Triacylglycerol 38.5 _+ 2.,
Substrate [1-14 C]ei,;osapentaenoic acid (20: 5 ( n - 3)) :
Oxidation produces 25.6 ± 3J Phosphofipids 68.9 ± 4.7 Triacylglyceroi 70.6 + 6.2
2.9 ± 0/# 2g J:O.2 2.4±0.6 43.5±6.5 31..~ ~-5.0 35.5±6.9 27.6 ± 6, 7 71.7~ 4- 0.6 28.9 ± 6.0
8.6±2.8 2.P±0.6 8.1±0.4 35.5±6.2 32..±~.1 32.9±5.1 77.5±6.7 89.5±5.9 79.6±9.1
7.0±0.3 6.8±0.5 6.7±0.7 69.3±3.6 59.0±4.0 77.4±1.0 ~ . 4 ± 1 . 2 48.1±3.5 31.5±3.5
26.4±4.1 22.0±3.5 75.3±6,3 69.0±4.0 39.3±4.7 37.1±3.5
22.1±1.3 23.0±3.4 69.0±4.9 62.0±3.5 67.3±8.7 66.7±5.4

82
the pewxisomal ~-oxidation compared to the control (Table l). Arachidonic acid (20 :4 (n -6 ) ) was not chain-elongated to the same extent as eicosapentaenoic acid (20: 5(n - 3)) was in adrenoleukodystrophy as well as in controls.
The distribution between chain elongation and chain shortening of erucic acid (22: l(n - 9)) in the cells from the patient with rhizomelic chondrodysplasia punctata was different from the control cells, the chain elonga- tion was increased and the chain shortening to oleic acid (18: l(n - 9)) was decreased while the chain shor- tening to eicosenoic acid (20: l ( n - 9)) was increased compared to the controls.
Table !! shows that very little erucic acid (22; l(n - 9)) is oxidized to CO2 and acid soluble products with all the fibroblast cultures used.
With [14-14C] erucic acid (22: l ( n - 9 ) ) a:~ the sub- strafe the radiolabelled esterified fatty acidg are to a large, extent incorporated in the phospolipid fraction in controls and X-linked adrenoleukodystrc~phy fibro- blasts. In the neonatal adrenoleukodystrophy fibro- blasts ¢ruci¢ acid (22: l ( n - 9)) is not chain shortened and incorporated in the triacylglycerol fraction. Previ- ous studies with hepatocytes and heart myocytes have also shown that little erucic acid is esterified in phos- pholipids while the fatty acid is efficiently incorporated in triacylglycerol [16].
The oxidation of adrenic acid (22:4O t - 6 ) ) to 14C- labelled CO2 and acid-soluble products was higher than with erucic acid (22: l ( n - 9)) as substr~te in the con- trol fibroblasts (Table II and Ret, 9). Ir the neonatal adrenoleukodystrophy fibroblasts the oxidation of adrenic acid (22 :4 (n - 6)) was impaired as previously observed in the Zellweger cells [9].
Table II shows that the oxidation and the ¢sterifica- tion of linolenic acid (18:3(n-3))~ arachidonic acid (20: 4(n - 6)) and eicosapentaenoic ~eid (20: 50, - 3)) was not influenced by the pcwxisomal defects in the neonatal adrenoleukodystrophy fibroblasts. The D- oxidation of the two C2o fatty acids was higher than with the Cls and the C22 fatty acids. Arachidonic acid ( 2 0 : 6 ( n - 6)) was preferentially incorporated into the phospholipids, while eicosapentaenoic acid ( 2 0 : 5 ( r - ~)) was equally incorporated into the phospholipids and into the triacylglycerols.
Discmslon
X-linked adrenoleukodystrophy differs clinically from the other peroxisomal diseases mentioned in that the patients appear healthy at birth and during the first years of life until the middle of the first decade when neurological symptoms develop progressively.
Since lignoceryl (24:0)-COA and palmityl (16:0)- CoA oxidation has been found to proceed normaly in X-linked adrenoleukodystrophy, fibroblasts and liver
biopsies [1,2,8], it is probable that the very long chain fatty acids in normal ceils are activated by a new enzyme, lignoceryl-CoA ligase before peroxisomal fl- oxidation, and that this enzyme is deficient in X-linked adrenoleukodystrophy.
The present finding of an efficient chain shortening of erucic acid (22:1(n-. 9)) and probably of adrenic acid ( 2 2 : 4 ( n - 6)) in fibroblasts from patients with X-linked adrenoleukodystrophy indicates a normal activation of the C22 unsaturated fatty acids to acyI-CoA esters in these cells. Thi,,; suggests ~hat long chain acyl- CoA synthetase (palmityl CoA ligase) is sufficiently active with the C22 unsaturated fatty acids as substrates to permit a normal pewxisomal chain shortening of these fatty acids. The existense of another very long chain acyl-CoA synthethase which might activate erucic a,:id and be different form both palmitoyl-CoA ligase and lignoceryl-CoA ligase cannot, however, be excluded.
Palmityl-CoA ligase is located to the endoplasmatic reticulum and to the outer mitochondrial membrane and is also present in the peroxisomes [17-19]. Al- though the enzyme is much more active with C1,~ and CIs fatty acid substrates, experiments with subcellular fractions have shown that erucic acid is probably also activated to a small but significant extent [20].
Previous works with rat hepatocytes strongly indicate that the chain shortening of erucic acid to eicosenoic acid (20: 1), oleic acid (18: 1) and palmitoleic acid (16:1) is a peroxisomal reaction [21,22].
A slow activation of C,4:0 and C:6: 0 fatty acids by a long chain acyI-CoA synthetase different from lignoceryi-CoA iigase, could also explain the observa- tion that C26:0/C22:0 and C24:o/C2::o ratios are lower in X-linked adrenoleukodyst~ophy [23] than in patients lacking peroxisomes such as neonatal adrenoleuko. dystrophy and the Zellweger syndrome.
The difierence in chain elongation between arach- ironic acid ( 2 0 : 4 ( n - 6)) and eicosape~taenoic acid (20: 5 ( n - 3)) observed both in human fibroblasts [24] and in isolated rat hepatocytes [25] has been explained to be the result of a preferential chai~ shortening (retro- conversion) of the chain-elongated product fermed from arachidoni,v acid (20: 4(n - 6)), adrenic acid (22 : 4(n - 6)). A futile cycle of elongation of 2 0 : 4 ( n - 6) and shortening of 22: 4(n - 6) might give a constant pool of available arachidonic acid (20: 4(n - 6 ) ) for prostanoi~ synthesis.
The present results show that the chain elongation to adrenic acid (22: 4(n - 6)) is not increased in cells with defects in the peroxisomal //-oxidation, and tha~ arachidonic acid (20: 4(n - 6)) is not chain-elongated to the same extent that eicosapentaenoic acid (20: 5(n - 3)i! is in tiles¢ cells. This difference in chain elongation i,*, thus not dependent on an intact peroxisomal fl-oxida- tion. The increased chain elongation of eicosa- pentaenoic acid (20: 5(n - 3)) compared to arachidonic

acid ( 2 0 : 4 ( n - 6)) is thus probably due to increased substrate availability or substrate specificity of the chain-elongation system or both.
The very long chain (24:0 and 26:0) fatty acids which accumulate in X-linked adrenoleukody~trophy and other peroxisomal fl-oxidation defects, are prob- ably formed by microsomal fatty acid chain elongation of shorter saturated fatty acids. Attempts have been made to inhibit this chain elongation by giving patients with X-linked adrenoleukodystrophy a diet rich in oleic acid [23,26], which competes with the saturated fatty acids for the microsomal chain-elongation enzymes [27,28]. This diet could, however, result in increased chain elongation of oleic acid to longer monounsatu- rated fatty acids such as erucic acid (22: l(n - 9)) which might accumulate and in itself be harmful. The present result that fibroblasts from the patients with X-linked adrenoleukodystrophy are able to shorten erucic acid (22: l ( n - 9)) should support the rationale of treating patients with X-linked adrenoleukodystrophy with a diet rich in oleic acid.
Acknowledgements
We thank Dr. Ola Skjeldal for providing fibroblasts. Erik Christensen is a Fellow of the Norwegian Cancer Society. The technical assistance of Mette Gregersen, Yngvar Johansen and Siri Tverdal, and the secretarial assistance of Marit S.K. Thoresen are greatly appreci- ated.
References
1 Hashmi, M., Stanley, W. and Singh I. (1986) FEBS Lett. 196, 247-250.
2 Wanders, R.J.A., Van R~ermund, C.WT, Van Wijlana, M.J.A., Nijenhuis, A.A., Tromp, A., Schutgens, R.B.H., Brouwer-Kelder, E.M., Schram, A.W., Tager, J.M., Van den Bosch, H. and Schalk- wijk, C. (1987) Ciin. Chim. Acta 165, 321-329.
3 Tager, J.M., Ten Harmsen Van Der Beek, W.A., Wanders, R.J.A., Hashimoto, T., Heymans, H.S.A., Van Den Bosch, H, Schutgens, R.B.H. and Scram, A.W. (1985) Biocbem. Biophys. Res. Commun. 123, 1269-1275.
4 Suzuki, Y., Orii, T., Mori, M., Tatibana, M. and Hashimoto, T. (1986) Clin. Chiw Acto !56, 191-196.
83
5 Chen, W.W., Watkins, P.A., Osumi, T., Hashimoto, T. and Moser, H.W. (1987) Proc. Natl. Acad. Sci. USA 84, 1425-1428.
6 Moser, H.W., Moser, A.B., Frayer, K.K., Chen, W., Schulman, J.D., O'Neil, B.P. and KJshimoto, Y. (1981) Neurology 31, 1 2 4 1 - 1249.
7 Moser, A.E., Sing, h, I., Brown, F.R., Ill, Solish, G.I., Kelley, R.I., Benke, P.J. and Moser, H.W. (1984) N. Engl. J. Med. 310, 1141-1146.
8 Goldfisclaer, S., Collins, J., Rapken, I., Coltoff-Schiller, B., Chang, C.H., Nigro, M., Black, V.H., Javit, N.B., Moser, H.W. and Lazarow, P.B. (1985) Science 227, 67-70.
9 Christensen, E., Hagve, T.A. and Christophersen, B.O. (1988) Biochim. Biophss. Acta 959, 95-99.
10 Heymans, H.S.A., Oorthys, J.W.E., Nelck, G., Wanders, R.J.A. and Schutgens, R.B.H. (1985) N. Engl. J. Med. 313, 187-188.
11 Christiansen, R., Borrebaek, B. and Bremer, J. (1976) FEBS L e t t .
62, 313--317. 12 Folch, J., Lees, M. and Sloan-Stanley, G.H. (1957) J. Biol. Chem.
226, 497-509. 13 Metca!f, L.D. and Schmitz, A.A. (1961) Anal. Chem. 33, 363-368. 14 Hagve, T.A. and Christophersen, B.O. (1986) Biochim. Biophys.
Acta 875, 165-173. 15 Lowry, O.H., Rosenbrough, N.J., Farr, A.L. and Randall, R.J.
(1959) Biol. Chem. 193, 265-275. 16 Bremer, J. and Norum, K.R. (1982) J. Lipid Res. 23, 243-256. 17 Osumi, T. and Hashimoto, T. (1984) Trends Biochem. Sci. 9,
317-319. 18 Mishina, M., Kamiryo, T., Tashiro, S. and Numa, S. (1978) Eur. J.
Biochem. 82, 347-354. 19 Krisans, S.K., Mortensen, R.M. and Lazarow, P.B. (1980) J. Biol.
Chem. 255, 9599-9607. 20 Normann, P.T., Thomassen, M.S., Christiansen, E.N. and
Flatmark, T. (1981) Biochim. Biophys. Acta 664, 416-427. 21 Christiansen, R.Z., Osmundsen, H., Borreb~ek, B. and Bremer, J.
(1978) Lipids 13, 487-491. 22 Norseth, J. and Clu'istophersen, B.O. (1978) FEBS Lett. 88,
353-357. 23 Watkins, P.A., Naidu, S. and lqoser, q.W. (1987) J. lnher. Metab.
Dis., 10 Suppl. 1, 46-53. 24 Bannerjee, N. and Rosenth~d M.-D. (1986) Biochim. Biophys.
A c t a 878, 404-411. 25 Christensen, E. Hagv~, T.-A. and Christophersen, Ii.O. (1986)
Biochim. Biophys. Acta 879, 713-321. 26 Rizzo, W.B., Philips, M.Wo, Dammann, A.L., Leshner, R.T., Jen-
nings, S.S., Avigan, J. and Proud, V.K. (1987) Ann. Neurol. 21, 232-239.
27 Saitoh, T., ¥oshida, S. artd Takeshita, M. (1988) Biochim. Bio- phys. Acta 960, 410-416.
28 Yoshida, S. and Takeshita, M. (1987) Arch. Biochem. Biophys. 254, 170-179.





![The endogenous subcellular localisations of the …...pholipid oxidation [22]. ACSL4 can supply arachidonyl-CoA and adrenic-CoA for incorporation into phosphatidy-lethanolamine and](https://static.fdocuments.us/doc/165x107/5ec2fce62e4af71b3e52bfc6/the-endogenous-subcellular-localisations-of-the-pholipid-oxidation-22-acsl4.jpg)












