
Acclimation of a non-indigenous sub-Arctic population: seasonal variation in thyroid function in...
6
Pergamon 0300-%29(95)00016-X Camp. Biochem. Physiol. Vol. I1 IA, NO. 2, pp. 209-214, 1995 Copyright 0 1995 Elsevier Science Ltd Printed in Great Britain. All rights resewed 0300-9629/95 $9.50 + 0.00 Acclimation of a non-indigenous sub-Arctic population: seasonal variation in thyroid function in interior Alaska Matthew Levine ,* Lawrence Duffy,? Dan C. Moore1 and Louis A. Matejl *Department of Psychiatry, Brooke Army Medical Center, Fort Sam Houston, TX 78234, U.S.A.; tInstitute of Arctic Biology and Department of Chemistry, University of Alaska Fairbanks, Fairbanks, AK 99775, U.S.A.; and $,Madigan Army Medical Center, Tacoma, Washington, U.S.A. Total, as well as free, T4 and T3 levels were obtained over four seasons for young male infantry soldiers assigned to interior Alaska. Significant seasonal variations were found in both T3 and T4. Total T4 and T3 levels were highest in winter, while free T4 and T3 levels were highest in early spring. Correlations with melatonin levels from a concurrent study showed an association between late day (17.00) mean spot melatonin levels during the preceding summer and T3 levels in winter and spring. Differences in seasonal T4 and T3 levels between indigenous and newly arrived people in the s&Arctic may be related not only to cold acclimation but also to light. Key words: Thyroid; Seasonal; Acclimation; Alaska; T3 (triiodothyronine); T4 (thyroxine); Melatonin; Diurnal. Comp. Biochem. Physiol. 1 I IA, 209-214, 1995. Introduction Seasonal fluctuations in thyroid function and size have been reported by a number of investigators at various latitudes and under a variety of climatic conditions (Duruisseau, 1965; Leppaluoto, 1994; Smals et al., 1977). With some exceptions. serum triiodothyronine (T3) levels have been found to be higher i,n winter than in summer (Lagoguey et al., 1977; Hamada et al., 1984; Nagata et al., 1976; Konno, 1978; Pasquali et al., 1984; Saljukov et al., 1992; Tkachev et al., 1991). Somewhat different findings have been reported from two studies performed on males before, during, and after assignment to duty in Antarctica (Reed et al., 1986, 1988). Serum T3 levels in- creased after 42 weeks of polar living, when Correspondence IO: Dr Lawrence K. Duffy, Institute of Arctic Biology. University of Alaska Fairbanks, AK 99715. U.S.A. Tel. 9074747525. Received 15 April 1994; revised 24 October 1994; accepted 26 October 1994. observations concluded, but the change was not significant. In Denmark, measurement of thyroid volume in 13 healthy males was found to vary by 23% from summer to win- ter, though no significant variations in T4, T3, or TSH could be demonstrated (Hegedus, 1987). Thyroid glands are necessary for animals to survive the winter in northern areas (Leppaluoto, 1994). Studies on animals have demonstrated increased thyroid activity after cold exposure (Seller, 1971; Hefco et al., 1975). Also, pineal- ectomized rats have shown significant increases in thyroid weight, with an opposite effect being obtained by the administration of melatonin (De Fronzo and Roth, 1972). To expand understanding of the acclimatiz- ation process and physiological adaptation to the arctic, as well as to clarify the function of the pineal-thyroid axis in the arctic, the thyroid function of a group of young, healthy, male infantry soldiers, passing their first full year 209
-
Upload
matthew-levine -
Category
Documents
-
view
212 -
download
0
Transcript of Acclimation of a non-indigenous sub-Arctic population: seasonal variation in thyroid function in...

Pergamon
0300-%29(95)00016-X
Camp. Biochem. Physiol. Vol. I1 IA, NO. 2, pp. 209-214, 1995 Copyright 0 1995 Elsevier Science Ltd
Printed in Great Britain. All rights resewed 0300-9629/95 $9.50 + 0.00
Acclimation of a non-indigenous sub-Arctic population: seasonal variation in thyroid function in interior Alaska
Matthew Levine ,* Lawrence Duffy,? Dan C. Moore1 and Louis A. Matejl *Department of Psychiatry, Brooke Army Medical Center, Fort Sam Houston, TX 78234, U.S.A.; tInstitute of Arctic Biology and Department of Chemistry, University of Alaska Fairbanks, Fairbanks, AK 99775, U.S.A.; and $,Madigan Army Medical Center, Tacoma, Washington, U.S.A.
Total, as well as free, T4 and T3 levels were obtained over four seasons for young male infantry soldiers assigned to interior Alaska. Significant seasonal variations were found in both T3 and T4. Total T4 and T3 levels were highest in winter, while free T4 and T3 levels were highest in early spring. Correlations with melatonin levels from a concurrent study showed an association between late day (17.00) mean spot melatonin levels during the preceding summer and T3 levels in winter and spring. Differences in seasonal T4 and T3 levels between indigenous and newly arrived people in the s&Arctic may be related not only to cold acclimation but also to light.
Key words: Thyroid; Seasonal; Acclimation; Alaska; T3 (triiodothyronine); T4 (thyroxine); Melatonin; Diurnal.
Comp. Biochem. Physiol. 1 I IA, 209-214, 1995.
Introduction
Seasonal fluctuations in thyroid function and size have been reported by a number of investigators at various latitudes and under a variety of climatic conditions (Duruisseau, 1965; Leppaluoto, 1994; Smals et al., 1977). With some exceptions. serum triiodothyronine (T3) levels have been found to be higher i,n winter than in summer (Lagoguey et al., 1977; Hamada et al., 1984; Nagata et al., 1976; Konno, 1978; Pasquali et al., 1984; Saljukov et al., 1992; Tkachev et al., 1991). Somewhat different findings have been reported from two studies performed on males before, during, and after assignment to duty in Antarctica (Reed et al., 1986, 1988). Serum T3 levels in- creased after 42 weeks of polar living, when
Correspondence IO: Dr Lawrence K. Duffy, Institute of Arctic Biology. University of Alaska Fairbanks, AK 99715. U.S.A. Tel. 9074747525.
Received 15 April 1994; revised 24 October 1994; accepted 26 October 1994.
observations concluded, but the change was not significant. In Denmark, measurement of thyroid volume in 13 healthy males was found to vary by 23% from summer to win- ter, though no significant variations in T4, T3, or TSH could be demonstrated (Hegedus, 1987).
Thyroid glands are necessary for animals to survive the winter in northern areas (Leppaluoto, 1994). Studies on animals have demonstrated increased thyroid activity after cold exposure (Seller, 1971; Hefco et al., 1975). Also, pineal- ectomized rats have shown significant increases in thyroid weight, with an opposite effect being obtained by the administration of melatonin (De Fronzo and Roth, 1972).
To expand understanding of the acclimatiz- ation process and physiological adaptation to the arctic, as well as to clarify the function of the pineal-thyroid axis in the arctic, the thyroid function of a group of young, healthy, male infantry soldiers, passing their first full year
209

210 M. Levine et al.
WINTER SPRING _ ,,-,
TIME
Fig. I. Daily and seasonal variations in total serum T4. ANOVA with univariate analysis between seasons indicated significant differences for all four times of day, across all four seasons: (02.00, N = II, P ~0.001; 08.00, N = 12. P <0.001; 10.30. N=7, P<O.OOl; 17.00, N=6, P = 0.030). Multivariate analysis also showed T4 levels to differ significantly between seasons (02.00, P = 0.017; 08.00, P < 0.001; 10.30, P = 0.025; 17.00, P = 0.015). Compari- sons between different times were non-significant. Bars
indicate standard error.
at 65”N, was studied as a psychophysiological research
Materials and Methods
Subjects
part of a larger project.
The study population consisted of volunteer male infantry soldiers, all assigned to Fort Wainwright, Alaska in Fairbanks (65”N). After IRB review, written informed consent was obtained from all volunteers prior to the start of the study. The mean age of the study group was 20.3 years (SEM = 0.4) with their ages ranging from 18 to 29 years. The average duration of stay in Alaska at the beginning of the year long study was 10.8 months. Mean height was 1.8 m, and mean weight was 75 kg. The racial com- position was 79% (34) Caucasian, 19% (8) black, and 2% (1) Hispanic. Birthplaces of subjects varied, with a tendency to under-representation from the southeastern and southwestern United States. Only one subject was born in Alaska. All but two participants were single and lived in the barracks.
Subjects were studied while living and working under conditions which were normal for infantry, assigned to Alaska. The study started with 43 subjects in June, 199 1. The number of subjects at each time point is denoted in figure legends. Throughout the year, the daily routine for the Second Brigade, to which all subjects were assigned, was 05.00 Wake-up, Physical Training from 05.30-06.30 daily for 5 days/week, and a daily morning outdoors formation at 08.45. Physical Training consisted of calisthenics and a 2-4 mile run in formation, generally in all but
the very harshest of weather conditions, such as during very heavy snowstorms. Exact measure- ments of the time spent outdoors are not avail- able, but an estimate would be a minimum of 2 hr per day, even during the coldest period in winter. This was probably considerably longer in the spring, as the temperatures rose somewhat.
Exposure to light was controlled at only the time of the 02.00 blood sampling. Subjects were instructed to don sunglasses on arising, and artificial light exposure was reduced to only that necessary for safely walking within the building. Exposure to artificial light was also minimized at the time of 08.00 samplings in winter, when no natural light was present. Light exposure was not controlled at other times. Subjects slept in their assigned rooms in the barracks, ate regular meals in the unit mess halls, and wore the required uniforms and clothing during duty hours. The basic uniform during the entire year long study period was the standard Army battle dress uniform (BDU). which in winter might be supplemented by thermal underwear. In winter, a light jacket was usually worn over the stan- dard BDU. Head cover was a standard utility cap, though in winter, a woolen balaclava was substituted to provide additional head and fa- cial protection. Gloves were worn in winter. The temperature in winter tended to remain at or below -20°C. All participants were screened for health problems and medications, and also For travel out of Alaska during the study period. They were also administered a brief recent events questionnaire. The questionnaire in- cluded eight major life events, which might have occurred at any time during the month prior to the quarterly samplings. In addition to thyroid studies, a number of other physiological measures were obtained; these measures were serial seasonal melatonin, cortisol, and testos-
SUMMER AUTUMN WINTER SPRING
Fig. 2. Free T4 in serum. Bar samples drawn at four separate times of day were pooled for analysis. ANOVA with multivariate repeated measures of analysis showed signifi-
cant variation (N = 19. P -c 0.001).
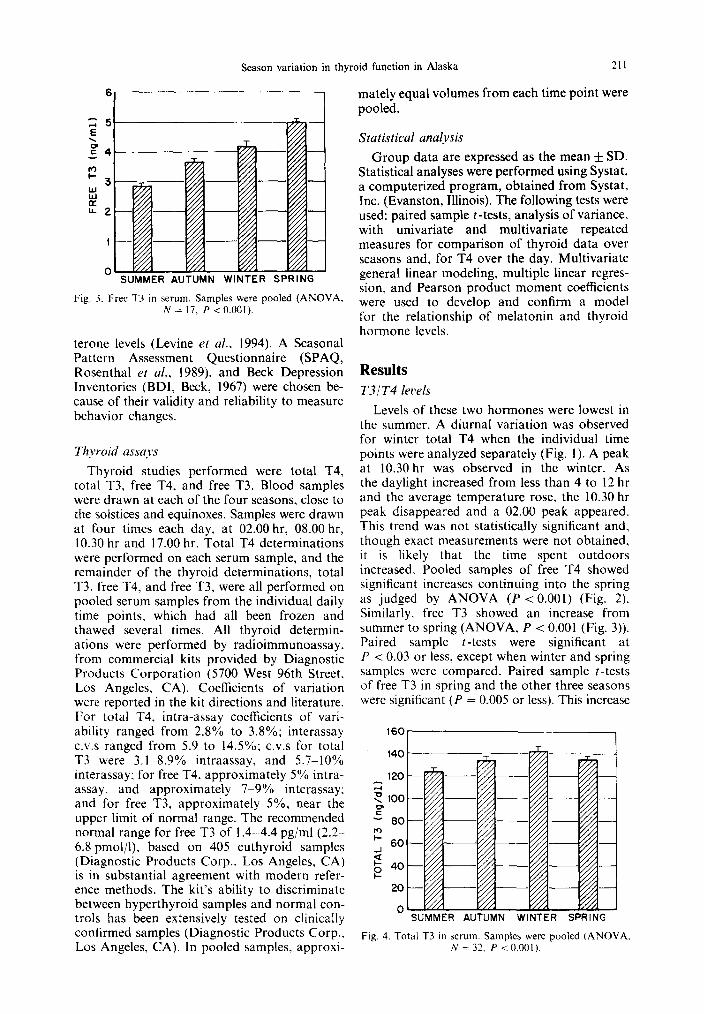
Season variation in thyroid function in Alaska 211
SUMMER AUTUMN WINTER SPRING
Fig. 3. Fret T3 in serum. Samples were pooled (ANOVA, N = 17, P <O.OOI).
terone levels (Levine et al.. 1994). A Seasonal Pattern Assessment Questionnaire (SPAQ, Rosenthal et al., 1989), and Beck Depression Inventories (BDI, Beck, 1967) were chosen be- cause of their validity and reliability to measure behavior changes.
Thyroid assu~‘s
Thyroid studies performed were total T4, total T3, free T4. and free T3. Blood samples were drawn at each of the four seasons, close to the solstices and equinoxes. Samples were drawn at four times each day, at 02.00 hr, 08.00 hr, 10.30 hr and 17.00 hr. Total T4 determinations were performed on each serum sample, and the remainder of the thyroid determinations, total T3. free T4, and free T3, were all performed on pooled serum samples from the individual daily time points, which had all been frozen and thawed several times. All thyroid determin- ations were performed by radioimmunoassay, from commercial kits provided by Diagnostic Products Corporation (5700 West 96th Street, Los Angeles, CA). Coefficients of variation were reported in the kit directions and literature. For total T4, intra-assay coefficients of vari- ability ranged from 2.8% to 3.8%; interassay c.v.s ranged from 5.9 to 14.5%; c.v.s for total T3 were 3.1-8.9% intraassay, and 5.7-10% interassay; for free T4, approximately 5% intra- assay, and approximately 7-9% interassay; and for free T3. approximately 5%, near the upper limit of normal range. The recommended normal range for free T3 of 1.4-4.4 pg/ml (2.2- 6.8 pmol/l), based on 405 euthyroid samples (Diagnostic Products Corp., Los Angeles. CA) is in substantial agreement with modern refer- ence methods. The kit’s ability to discriminate between hyperthyroid samples and normal con- trols has been extensively tested on clinically confirmed samples (Diagnostic Products Corp., Los Angeles, CA). In pooled samples, approxi-
mately equal volumes from each time point were pooled.
Statistical analysis
Group data are expressed as the mean + SD. Statistical analyses were performed using Systat, a computerized program, obtained from Systat, Inc. (Evanston, Illinois). The following tests were used: paired sample t-tests, analysis of variance. with univariate and multivariate repeated measures for comparison of thyroid data over seasons and, for T4 over the day. Multivariate general linear modeling, multiple linear regres- sion, and Pearson product moment coefficients were used to develop and confirm a model for the relationship of melatonin and thyroid hormone levels.
Results T.31 T4 lewls
Levels of these two hormones were lowest in the summer. A diurnal variation was observed for winter total T4 when the individual time points were analyzed separately (Fig. 1). A peak at 10.30 hr was observed in the winter. As the daylight increased from less than 4 to 12 hr and the average temperature rose, the 10.30 hr peak disappeared and a 02.00 peak appeared. This trend was not statistically significant and, though exact measurements were not obtained, it is likely that the time spent outdoors increased. Pooled samples of free T4 showed significant increases continuing into the spring as judged by ANOVA (P < 0.001) (Fig. 2). Similarly, free T3 showed an increase from summer to spring (ANOVA, P < 0.001 (Fig. 3)). Paired sample z-tests were significant at P cc 0.03 or less, except when winter and spring samples were compared. Paired sample r-tests of free T3 in spring and the other three seasons were significant (P = 0.005 or less). This increase
SUMMER AUTUMN WINTER SPRING
Fig. 4. Total T3 in serum. Samples were pooled (ANOVA. N=32. P<O.O01).

212 M. Levine et (I/
.I’ - , - _I - ,
FALL WINTER SPRING
Fig. 5. Ratios of seasonal free T4 and free T3
contrasts with total T3 where the mean levels fell slightly in spring (P < 0.001 ANOVA (Fig. 4)). When the free T3/free T4 ratio was calculated, a steady increase was found from summer to the following spring (Fig. 5).
Other hortnones and w,eig/zt gain
Modeling with a backward stepping pro- cedure, entering and removing at the P = 0.05 level of significance with a minimum tolerance of P = 0.01 for entrance into the model, and based on previously reported data on melatonin (Levine et al., 1994b) and mood (Levine et ~1.. 1994a), a model was developed which indicated a correlation (P = 0.048) between the late day (17.00) mean spot melatonin level in summer. and free T3. Free T4 was also correlated with melatonin levels in the model.
The same level of significance was obtained when this model was tested by multiple linear regression. Specifically, summer free T4 and free T3, as well as free T3 later in the year, in winter and spring. were found to be significantly related to late daytime summer melatonin levels. .‘.dditionally. moderately strong correlations were found between thyroid hormone levels and weight gain (1. = 0.576, 0.578, and 0.432 for fall. winter, and spring, P = 0.002) though not with appetite, subjective energy levels, or mood.
Discussion
The findings of significant seasonal variations in thyroid function in a non-indigenous popu- lation in the sub-Arctic confirm previous reports of seasonal variations in thyroid hormonal levels (Dessypris it al.. 1978; Hamada rt al., 1984; Tkachev et al., 1991). The magnitude of the increase in the metabolically active
triiodothyronine (free T3). beyond normal levels in spring, is the first report of seasonal elevations to this range in normal individuals (Borst et a/., 1983). Tkachev et al. (1991) reported that serum T3 levels in natives were higher in winter in the Archangels region of Russia. This contrasts with our finding of higher levels in the spring for newly arrived non-natives in Alaska. The significance of these differences is not clear, but it appears from these findings that physiological adap- tation to extreme cold in a sub-Arctic environ- ment, with seasonal variation in photoperiod, may be associated with a relative hyperthyrox- inemia during different seasons. This seasonal variation may be related to duration of acclimatization.
A number of other unique aspects of this study, including the relatively short period of time, less than I year, spent by the subjects in the sub-Arctic prior to the start of the study period, the relative uniformity of protective clothing and exposure to the elements, as well as uniform diet, living and working patterns. suggests that the regulation of thyroid related metabolism at high latitudes may be more com- plicated than previously believed. The findings reported here, which differ from those in other polar populations (Saljukov ef ul., 1992) as well as those studied within a very short period of time after arrival in the Antarctic (Reed et (I/., 1986) strongly suggest that psychophysiological adaptation may have other determinants prior to actual cold exposure. Melatonin may serve as a regulator or synchronizer of thyroid glandular activity by permitting the priming of the organ- ism early. for increased demands placed on it by seasonal cold exposure in later months. Small animal studies have shown a similar relationship between thyroid cellular activity and the pineal (Lewinski rt ~1.. 1987).
Experimental support for this hypothesis is available from animal research (Lewinski et (I/., 1992; Haldar et al., 1992; Zwirska-Korezza et a/., 1992; Kniazewski et al., 1990). The exist- ence of different melatonin effects on thyroid secretory processes, depending not only on the route of administration but also on timing and OII the dose of melatonin, is well documented in hamsters (Hoover ci (II., 1992). From studies on male Wistar rats (Haldar rt ul., 1992). late afternoon melatonin injections or melatonin im- plants are inhibitory both for thyroid hormone secretion, and also for growth of the gland. The relationship between the two glands appears to be reciprocal, as it has also been shown experimentally that thyroid hormones have the capacity to increase the mean cellular volume of pinealocytes in organ culture (Lewinski et trl.. 1987).

Season variation in thyroid function in Alaska 213
Thyroid hormones undoubtedly play a significant role in cold adaptation through their effects on thermogenesis. The near linear found in the free T3/free T4 ratio from summer through autumn, winter and spring points towards an increase in the peripheral conversion of T4 to T3 as a major source of the increase in T3. Melatonin may exert an effect on conversion of T4 to T3 through its action on the tissue enzyme, thyroxine 5’-deiodinase, which con- verts T4 to T3. In experimental studies on certain mammals (Puig-Domingo et al., 1989), melatonin has been found to stimulate type-II thyroxine 5’-deiodination in the brown adipose tissue of Syrian hamsters. Type-II thyroxine 5’-deiodinase is a selenocysteine protein, which generates T3 largely for the plasma pool. Also, studies by Silva (1993) have shown that the concentration of T3 can be locally regulated in brown adipose tissue.
A local feedback mechanism, with a mild elevation of serum T3 levels, as reported here, may indicate the role for melatonin in thermo- genesis in humans. A hypothesis of a dual mechanism of action of melatonin on thyroid metabolism and thermogenesis, should be the object of further study.
Acknon,lrdRements-This work was supported, in part, by the Institute of Arctic Biology, the Alaska-Siberian Medical Research Program and the U.S. Army. We gratefully acknowledge the excellent technical assistance of Andree Porchet and helpful discussions with Dr Putilov. We are especially appreciative to Linnea Delatour in putting together this manuscript, and also to Paula Fernald for assistance with the graphics.
References
Beck A. T. (1967) Measurement of depression: The de- pression inventory. In Depression, Causes and Treatment.
pp. I86 -207. University of Pennsylvania Press. Borst G. C.. Eil C., Burman K. D. (1983) Euthyroid
hyperthyroxinemia. Ann. Inr. Med. 98, 366-378. De Fronzo R. A.. Roth W. D. (1972) Evidence for the
existence of a pineal-adrenal and pineal-thyroid axis. ,4cta Endocr. 70, 31 -42.
Dessypris A., Saki T., Kujarinta M., Miettinen A., Lamberg B. A. (1978) Serum thyroid hormones, TSH, testosterone and auto-antibody levels in adult inhabitants of Lapland in summer. Arcf. Med. Res. Rep. 23, 33-42.
Duruisseau J. (1965) Seasonal variation of PBI in healthy Montrealers. J. clin. Endocr. Metnb. 25, 1513-I 515.
Hamada N.. Ohno M., Morii H., No J., Yamakawa J., Inaba M.. Ikeda S. and Masahisa W. (1984) Is it necessary to adjust the replacement dose of thyroid hormone to the season in patients with hypothyroidism? Melobolism 33, 215 218.
Hefco E. A., Krulich L., Illner P. and Larsen P. R. (1975) Effect of acute exposure to cold on the activity of the hypothalamic-pituitary-thyroid system. Endocrinology 97. I185m 1195.
Hegedus L. (1987) Seasonal variation in thyroid size in healthy males. Horm. Metab. Res. 19, 391-392.
Hoover P. A., Vaughan M. K., Little J. C. and Reiter R. J. (1992) N-methyl-p-aspartate does not prevent effects of
melatonin on the reproductive and thyroid axes of male Syrian hamsters. J. Endocr. 133, 51-58.
Kniazewski B.. Sobieraj H., Zwirska-Korczala K.. Buntner B. and Ostrowska Z. (1990) Influence of melatonin on thyroid secretion in pinealectomized rats. Endocr. E.YP.
24, 317-324. Konno N. (1978) Comparison between the thyrotropin
response to thyrotropin-releasing hormone in summer and that in winter in normal subjects. Endocr. Japan. 25, 635 -639.
Lagoguey M., Reinberg A.. Cesselin F.. Antreassian J., Delassalle A. and Lagoguey A. (1977) Circannual and circadian rhythms in plasma cortisol, renin activity, thyroxine, and in urinary aldosterone 01 five healthy young Parisian males. ChronohiologJ 2, 126.
Leppaluoto J. (1994) The thyroid gland and winter. .4rc,r. Med. Res. 53, 2-4.
Levine M. E., Duffy L. K, and Bowyer R. T. (1994a) Relationship of fatigue, sleep and alcoholism, with sea sonal hormone levels in the far north. Drugs and Society 8, 61-70.
Levine M. E., Duffy L. K. and Milliron A. N. (l994b) Diurnal and seasonal rhythms of melatonin. cortisol and testosterone in interior Alaska. Arc!. Med. Rex 53, 25_~34.
Lewinski A., Wajs E., Karbownik M. (1992) Effect of pineal-derived indolic compounds and of certain neuro- peptides on the growth processes in the thyroid gland. Thvroid. clin. E.up. 4, I1 15.
Lewinski A.. Webb S. M. and Reiter R. J. (1987) Inhibition of thyroid growth: its involvement in a possible negative feedback interaction between both glands. Pineul Rcs. Ret). 5, 69-94.
Nagata H., Izumiyama T.. Kamata K.. Kono S., Y ukimura Tawata M., Aizaw T. and Yamada T. (1976) An increase of plasma triiodothyronine concentration in man in a cold environment. J. c/in. mdocr. Meroh. 43, 1153. 1156.
Pasquali R., Baraldi G., Cassimirri F., Matteoli L.. Capelli M.. Melchionda M., Capani F. and Labo G. (1984) Seasonal variations of total and free thyroid hormones in healthy men: a chronobiological study. .4(,/a Endow. 107, 42 48.
Puig-Domingo M.. Guerrero J. M., Menendez-Pelaez A. and Reiter R. J. (1989) Melatonin specifically stimulates type-II thyroxine 5’-deiodination in brown adipose tissue of Syrian hamsters. J. Endocr. 122, S53- 556.
Reed H. L.. Burman K. D., Shakir K. M. M. and O’Brian J. T. (1986) Alterations in the hypothalamic-pituitary thyroid after prolonged residence in Antarctica. C/m Endow. 25, 55 65.
Reed H. L., Ferreior J. A., Shakir K. M. M.. Burman K. D. and O’Brian J. T. (1988) Pituitary and peripheral hormone responses to T3 administration during Antarctic residence. Am. J. Phvsiol. 254, E733 E739.
Rosenthal N. E.. BradtG. H. and Wehr T. A. (1989) How seasonal are you’? In Serrson.s c!/’ t/n, Mind. Bantam Books.
Saljukov V. B.. Lemza S. V., Kucher A. M. and Puzyrev V. P. (1992) The role of hereditary factors in phenotypic variability of hormone levels in the population genetically adapted to circumpolar environment. Arc,/. Med Re.v. 51, I43 -149.
Seller E. A. (1971) Thyroid status in relation to catechol- amines in cold and warm environment. Can. J. Ph~.sro/.
Pharmac. 49, 268-275. Silva J. E. (1993) Hormonal control of thermogenesis
and energy dissipation, Trends Endocr. Meruh. 4, 25-32.
Smals A. G. H.. Ross H. A. and Kloppenberg P. W. C. (1977) Seasonal variation in serum T3 and T4 levels in man. J. c/in. endow. Mrtob. 44, 998 1001.

214 M. Levine et al.
Tkachev A. V.. Ramendskaya E. B. and Bojko J. R. (1991) growth process in male Wistar rats: comparison with Dynamics of hormone and metabolic state in polar effects of other, indole substances. J. Pined Re.7, 13, state in polar inhabitants depend on daylight duration. 158-166. Arc!. Med. Res. SO, 152-155. Zwirska-Korezza K., Kniazewski B.. Ostrowska 2.
Wajs E. and Lewinski A. (1992) Inhibitory influence of late and Bunter B. (1992) Influence of melatonin on rat afternoon melatonin injections and the counter-inhibitory thyroid, adrenals and testis secretion during the day. action of melatonin-containing pellets on thyroid Folia. Histochem. Cyrobiol. 29, 19-24.


















