
AAssppeecctt ooss aeeccoollóóggiiccoss dde...
150
Universidad Central de Venezuela Facultad de Ciencias Instituto de Zoología y Ecología Tropical Postgrado en Ecología Tesis Doctoral “Aspectos ecológicos de microalgas con potenciales biotecnológicos” Presentada por Lic. Rubén Torres CI V-11.405.173 Tutores Dra. Evelyn Zoppi de Roa Dr. Diego Rodríguez Caracas – 2012
Transcript of AAssppeecctt ooss aeeccoollóóggiiccoss dde...

Universidad Central de Venezuela
Facultad de Ciencias Instituto de Zoología y Ecología Tropical
Postgrado en Ecología
Tesis Doctoral
““AAssppeeccttooss eeccoollóóggiiccooss ddee mmiiccrrooaallggaass
ccoonn ppootteenncciiaalleess bbiiootteeccnnoollóóggiiccooss””
PPrreesseennttaaddaa ppoorr
LLiicc.. RRuubbéénn TToorrrreess
CCII VV--1111..440055..117733
TTuuttoorreess
DDrraa.. EEvveellyynn ZZooppppii ddee RRooaa
DDrr.. DDiieeggoo RRooddrríígguueezz
Caracas – 2012

2

3
Las algas son vegetales que crecen en agua, tanto dulce como salada. En el
océano constituyen el principal componente del plancton marino. Tuvieron mucho que ver con el origen de la vida en el ámbito marino; fueron los
primeros organismos en realizar la fotosíntesis clorofílica. Van desde los microscópicos organismos unicelulares (como las espirulinas) hasta las
gigantescas kelp (el ser vivo más largo del planeta).
Fuente: Almacén Natural

4

5
CONTENIDO
Índice de figuras ........................................................................................................... 7
Índice de tablas ........................................................................................................... 10
Resumen .................................................................................................................... 12
1. Introducción ............................................................................................................ 14
1.1 Una breve reseña histórica ............................................................................ 14
1.2 Evolución y ecología de las microalgas ......................................................... 14
1.3 Aspectos biogeográficos ............................................................................... 17
1.4 Importancia biotecnológica de las microalgas ............................................... 18
2. Antecedentes en Venezuela ................................................................................... 22
3. Justificación ............................................................................................................ 23
4. Hipótesis ................................................................................................................. 24
5. Objetivos ................................................................................................................. 25
6. Metodología ............................................................................................................ 26
6.1 Ecología de poblaciones de especies cultivadas ........................................... 26
6.2 Ecología de comunidades fitoplanctónicas .................................................... 39
7. Resultados .............................................................................................................. 57
7.1 Cultivos ......................................................................................................... 57
7.2 Ensayos con poblaciones cultivadas de Arthrospira platensis ....................... 65
7.3 Comunidades fitoplanctónicas ....................................................................... 86
8. Discusión .............................................................................................................. 116
8.1 Cultivos ....................................................................................................... 116
8.2 Ensayos con poblaciones de Arthrospira platensis ...................................... 117
8.3 Comunidades fitoplanctónicas ..................................................................... 123
9. Conclusiones ........................................................................................................ 127
10. Bibliografía .......................................................................................................... 128
11. Enlaces ............................................................................................................... 135
Apéndice 1 ................................................................................................................ 137
Apéndice 2 ................................................................................................................ 142
Apéndice 3 ................................................................................................................ 147

6

7
Índice de figuras
Figura 1. Esquema ilustrado donde se muestra el escalamiento de microalgas de medio líquido a medio sólido con el empleo de la técnica del asa de estaño. ............. 29
Figura 2. Esquema donde se muestra el escalamiento de poblaciones de microalgas a fiolas de diferentes volúmenes hasta alcanzar los cultivadores a gran escala. ........... 30
Figura 3. Imágenes donde se muestran las diferentes etapas de escalamiento del cultivo en medio líquido en el sistema integrado LOA-Ficotrón: (a) cultivos a escala pequeña en condiciones controladas (Cámara de Crecimiento, LOA - IZET); (b), (c) y (d) cultivos a cielo abierto en el Ficotrón, IDEA, en botellones de 5 L, tanques circulares de 500 L y cultivadores tipo carrusel de 2000 L, respectivamente. ............. 31
Figura 4. Esquema para la preparación de cada medio (tratamiento) del diseño factorial fraccionado 26-1. ............................................................................................. 33
Figura 5. Esquema de la disposición espacial de los tanques cilíndricos en el Ficotrón para la ejecución del diseño de bloques completos aleatorizados. ............................. 34
Figura 6. Ubicación de los sitios de muestreo (el mapa se encuentra en www.guiageo-ameri cas.com/mapas/venezuela.htm). ....................................................................... 39
Figura 7. (a) Mapa de Venezuela donde se destaca a la península de Paria en un recuadro, (b) ubicación del área de estudio y (c) vista panorámica del humedal “Palmares III” desde una carretera que lo bordea al norte, la vegetación herbácea cubre la totalidad de su superficie y forma bandas monoespecíficas, evidenciadas por las tonalidades distintas del color verde, que cubren toda su superficie sin formar espejos de agua (tomado de Torres y Zoppi de Roa 2010)......................................... 41
Figura 8. Pluviodiagrama con precipitaciones medias de 47 años (1953-2000) del sur de la península de Paria (datos tomados de la Dirección de Meteorología del MARN 2000). ......................................................................................................................... 42
Figura 9. Vista satelital del área de estudio (humedal herbáceo de El Clavo). La línea blanca dibujada en el centro de la imagen señala el transecto levantado en la salida de campo (fuente: http://earth.google.com/). .................................................................... 43
Figura 10. Tomas fotográficas parciales de las dos zonas de vegetación emergente estudiadas: (a) vista de la amplia zona central de Hymenachne amplexicaulis, en primer plano la zona de Heliconia marginata que bordea todo el litoral sur; (b) zona de H. marginata (fotos: Carlos Lugo). .............................................................................. 44
Figura 11. Ubicación geográfica del área de estudio al sur de Monagas (Orinoco bajo), con detalle de la localización en el mapa y fotografía panorámica de Macapaima, uno de los cuerpos de agua visitados (foto: Rubén Torres). .............................................. 46
Figura 12. Imagen correspondiente a un sector de la orilla de la fosa El Caracol, (Municipio Aragua, Edo. Anzoátegui, 2009), donde se pueden apreciar los desechos petroleros que conforma parte del fondo de la misma (Foto: Olaf Ilzins). .................... 47
Figura 13. Ubicación de la laguna de Boca Chica (círculo azul) en la península de Macanao, isla de Margarita, estado Nueva Esparta (mapa: http://www.disfrutevenezuela.com/Municipio-Peninsula-de-Macanao-Mapa.html)....... 48
Figura 14. Marcha analítica simplificada para la determinación de nitrógeno en muestras de agua con el método Kjeldahl (1883) (continúa en la página siguiente). .. 52
Figura 15. Marcha analítica simplificada para la determinación del fósforo por el método colorimétrico de Murphy – Riley (1962). Las dos imágenes inferiores fueron tomadas de una presentación digital del curso de Ecología de Humedales, Postgrado en Ecología, IZET, 2009. ............................................................................................ 53

8
Figura 16. Algunas imágenes tomadas bajo microscopio invertido de las poblaciones de las diferentes cepas de Arthrospira spp. en la Cámara de Crecimiento (LOA-IZET).
................................................................................................................................... 59
Figura 17. Algunos cultivos de microalgas de interés biotecnológico presentes en la Cámara de Crecimiento (LOA, IZET). ......................................................................... 62
Figura 18. Curvas de crecimiento poblacional de Arthrospira platensis en (a) fiolas y (b) cilindros. La densidad poblacional está expresada en términos de absorbancia a 680 nm. ............................................................................................................................. 66
Figura 19. Producción de biomasa seca en fiolas y cilindros de 2 L para cada medio mineral. La biomasa está expresada en gramos por dos litros. ................................... 67
Figura 20. Valores comparativos de pH mostrados por los diferentes medios de cultivo. En la figura se señala con un óvalo rojo el lapso de disminución de pH del medio – – y la contaminación del mismo por la cianobacteria Microcystis aeruginosa. .................. 67
Figura 21. Valores comparativos de conductividad (S/cm) mostrados por los diferentes medios de cultivo. ....................................................................................................... 68
Figura 22. Curva de calibración A680 vs. Peso seco (mg). Las variables tienen una relación lineal (R2 = 0,9935). ....................................................................................... 68
Figura 23. Crecimiento poblacional de Arthrospira platensis en cuatro medios de
cultivo. En la fase de crecimiento rápido A680 y el tiempo presentaron correlaciones lineales fuertes, como se evidencia en los valores del coeficiente de determinación (R2). ............................................................................................................................ 70
Figura 24. Crecimiento poblacional de Arthrospira platensis en cuatro medios de
cultivo (filamentos por litro) en la Cámara de Crecimiento. ......................................... 70
Figura 25. Curvas de crecimiento poblacional de Arthrospira platensis obtenidas en un
ensayo factorial fraccionado 26-1 con combinaciones aleatorias de niveles mínimos y centrales de cinco macronutrientes (tratamientos). La densidad poblacional fue medida indirectamente con la absorbancia a una longitud de onda de 680 nm, correspondiente al rojo dentro del espectro visible, pico de absorción de la clorofila a. ........................ 72
Figura 26. Diagrama de columnas mostrando en orden creciente los tiempos de duplicación (tg) de los diferentes tratamientos del ensayo multifactorial. ..................... 79
Figura 27. Biomasa seca (g/L) cosechada en los diferentes tratamientos del ensayo factorial 26-1. Los valores se ordenan en forma creciente. ........................................... 81
Figura 28. Diagramas circulares que muestran las variaciones porcentuales en grupos de filamentos de diferentes tallas en una población de Arthrospira platensis, a lo largo
de un periodo de incubación que duró 30 días. La cepa se cultivó en el medio optimizado en el ensayo factorial fraccionado 26-1 (tratamiento 36). ............................ 82
Figura 29. Dinámicas de crecimiento poblacional de los tres componentes de tallas de filamentos de Arthrospira platensis: (a) filamentos con 2 células, (b) filamentos con
cuatro células y (c) filamentos con más de cuatro células. La letra “Y” en la ordenada es la densidad (filamentos/L) y en la abscisa el tiempo está dividido en intervalos de tres días. Salida: PAST. .............................................................................................. 83
Figura 30. Dinámicas de crecimiento poblacional de Arthrospira platensis a partir de
tres extracciones de volúmenes en fase de saturación. De izquierda a derecha se muestran las curvas de crecimiento a partir de 25%, 50% y 75% de extracción. ........ 84
Figura 31. Absorbancias medias (λ = 680 nm) de los cultivos en fase de crecimiento rápido, para tres profundidades, izquierda a derecha: 15 cm (barras azules), 25 cm (barras rojas) y 35 cm (barras amarillas). .................................................................... 85

9
Figura 32. Biomasa seca total (g/L) en los tres bloques. ............................................. 85
Figura 33. Biplot de los dos primeros componentes principales del ACP para las variables (especies) y casos (parches de vegetación) escogidas para caracterizar el ecosistema del humedal Palmares III en noviembre de 2008. Los dos primeros componentes principales acumularon 89,965% de la inercia total del sistema. Salida: MVSP 3.0. .................................................................................................................. 91
Figura 34. Dendrograma del Análisis de Agrupamiento o “Cluster Analysis” derivado del conjunto de especies fitoplanctónicas y zooplanctónicas más importantes colectadas en el ACP. Salida: MVSP 3.0. ................................................................... 92
Figura 35. Algunas especies de microalgas representantes del fitoplancton del humedal de El Clavo, Barlovento, Edo. Miranda: (a) Lyngbya lutea (Cyanobacteria), 400x; (b) Pinnularia sp. (Bacillariophyta), 400x; (c) Asterococcus limneticus (Chlorophyta), (d) Micrasterias sol (Chlorophyta), 250x; (e) Closterium ehrenbergii (Chlorophyta); 250x; (f) Spirogyra ternata (Chlorophyta), 250x. Fotos: Rubén Torres,
cámara digital PAX-CAM acoplada a microscopio invertido y a computador (programa PAX-IT!). ..................................................................................................................... 98
Figura 36. Variaciones temporales de (a) conductividad eléctrica y (b) oxígeno disuelto a medida que la lámina de agua disminuyó en el parche de Heliconia marginata (enero
– febrero 2009). .......................................................................................................... 99
Figura 37. Concentraciones de nitrógeno (N) en las muestras de agua colectadas en la zona de Heliconia marginata en la primera salida de campo al humedal de El Clavo
(05/01/2009). ............................................................................................................ 100
Figura 38. Curva de calibración para la obtención de fósforo total en agua de las muestras tomadas en la zona de Heliconia marginata en la primera salida de campo al
humedal de El Clavo (05/01/2009). ........................................................................... 101
Figura 39. Biplot de los dos primeros componentes principales del ACP para los ambientes lagunares del Orinoco bajo, sur de Monagas. Los dos primeros componentes principales retuvieron 95,4% de la inercia total del sistema multidimensional original. Salida: MVSP 3.0. ............................................................ 107
Figura 40. Imágenes que muestran la composición de especies fitoplanctónicas de una charca fangosa, específicamente proveniente del borde exterior de un área de pozos petroleros en el norte del Edo. Bolívar, 2009. ........................................................... 109
Figura 41. Composiciones porcentuales de las divisiones de organismos procariotas y eucariotas integrantes de la comunidad del fitoplancton en las regiones estudiadas en el país a lo largo del estudio. .................................................................................... 115

10
Índice de tablas
Tabla 1. Especies de microalgas con potencialidades para generación de biocombustibles. Contenido de aceites en base seca (tomada de Albarracín 2007). .. 20
Tabla 2. Composición química del medio Spirulina (Schlösser 1994). ........................ 26
Tabla 3. Diseño factorial 22 para ensayos de laboratorio. El experimento se simplificó a cuatro medios de cultivo sin replicación. .................................................................. 31
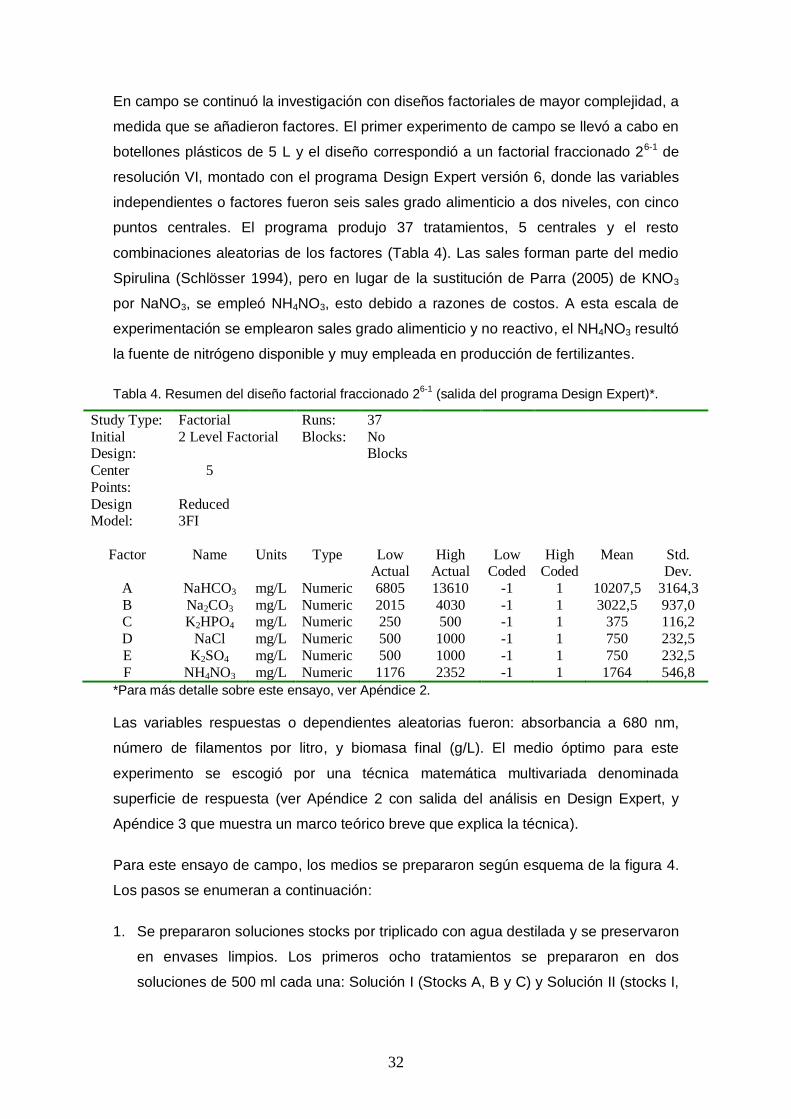
Tabla 4. Resumen del diseño factorial fraccionado 26-1 (salida del programa Design Expert)*. ...................................................................................................................... 32
Tabla 5. Medios de cultivos preparados en laboratorio para crecimiento de poblaciones de microalgas y modalidades de preparación y recipientes. ....................................... 57
Tabla 6. Concentraciones milimolares (mM) de los macronutrientes totales (representados como elementos, a excepción del carbono que se representa en las formas de los aniones carbonato y bicarbonato) y relaciones milimolares sodio/potasio en los cuatro medios minerales comparados en el diseño factorial 22. En rojo se destacan las concentraciones milimolares de Na+ y K+, y en azul las de Cl-, anión directamente involucrado con el Na+. .......................................................................... 65
Tabla 7. Concentraciones milimolares totales de los macronutrientes que integran cada uno de los medios de cultivo preparados. En rojo se destaca la relación 4:1 de K y Na en el medio 1. ............................................................................................................. 69
Tabla 8. Tasa de crecimiento per cápita de Arthrospira platensis en la fase de crecimiento rápido, biomasa seca producida en cada medio de cultivo y pH inicial y final. ............................................................................................................................ 71
Tabla 9. Tasas de crecimiento per cápita y capacidades de carga en cultivos de Arthrospira platensis para el ensayo factorial fraccionado 26-1. En rojo se destacan los
medios con valores mayores para uno o ambos parámetros poblacionales obtenidos de forma experimental. ............................................................................................... 77
Tabla 10. Análisis de varianza para el diseño factorial fraccionado 26-1 (salida del programa Design Expert). Los números destacados en rojo son valores de p<0,05. .. 80
Tabla 11. Resumen del Análisis de Varianza (salida de MICROSOFT EXCEL). ......... 86
Tabla 12. Composición de especies y abundancia (células/litro) de los taxa fitoplanctónicos presentes en las zonas de vegetación en noviembre de 2008. Bm (Brachiaria mutica), EcBmSe (ecotono B. mutica - Sesbania exasperata), Se (S. exasperata), EcSeCa (ecotono S. exasperata - Cyperus articulatus), Ca (C. articulatus), EcCaTd (ecotono C. articulatus-Typha dominguensis), Td (T. dominguensis) y EcTdSe (ecotono T. dominguensis - S. exasperata)......................... 87
Tabla 13. Algunos índices de diversidad empleados para caracterizar la comunidad fitoplanctónica de Palmares III en sus diferentes parches de vegetación monoespecífica y ecotonos (noviembre 2008). Salida: PAST. .................................... 89
Tabla 14. Distancias Jaccard entre los parches de vegetación acuática del humedal Palmares III, noviembre de 2008, definidas a partir a las abundancias de las especies fitoplanctónicas. Los valores están acotados entre 0 y 1, los destacados en rojo indican similitudes altas y en azul se señala a la pareja de vegetaciones con menor similitud. Salida: PAST. ............................................................................................................. 90
Tabla 15. Variables fisicoquímicas y concentraciones de cationes y aniones del agua, determinados en las zonas de vegetaciones monoespecíficas estudiadas en el humedal de Palmares en noviembre de 2008. ............................................................ 92

11
Tabla 16. Composición de especies y abundancias (células/litro) de los taxa fitoplanctónicos presentes en las zonas de vegetación para agosto de 2009. EcBm1Ca y EcCaBm2 (ecotonos B. mutica - C. articulatus); EcBm2Td (ecotono B. mutica - T. dominguensis). ........................................................................................................... 93
Tabla 17. Algunos índices de diversidad empleados para caracterizar la comunidad fitoplanctónica de Palmares III en sus diferentes parches de vegetación monoespecífica y ecotonos (agosto 2009). Salida: PAST. .......................................... 94
Tabla 18. Distancias Jaccard entre los parches de vegetación acuática del humedal Palmares III, agosto de 2009, definidas a partir a las abundancias de las especies fitoplanctónicas. Los valores están acotados entre 0 y 1, los destacados en rojo indican similitudes altas y en azul se señala a la pareja de vegetaciones con menor similitud. Salida: PAST. ............................................................................................................. 94
Tabla 19. Variables ambientales medidas en agosto de 2009..................................... 95
Tabla 20. Grupos (Taxa) fitoplanctónicos identificados en los diferentes ambientes de vegetación en El Clavo, Barlovento, Edo. Miranda (enero de 2009). .......................... 95
Tabla 21. Algunos índices de diversidad empleados para caracterizar la comunidad fitoplanctónica del humedal de El Clavo (enero 2009). Salida: PAST.......................... 97
Tabla 22. Índice de similitud de Jaccard para las tres zonas estudiadas en el Clavo, Edo. Miranda (enero de 2009). El análisis se hizo en PAST. ...................................... 97
Tabla 23. Valores medios desviaciones estándares de las variables fisicoquímicas tomadas en la zona de Heliconia marginata del humedal herbáceo de El Clavo, Edo.
Miranda en enero - febrero de 2009. ........................................................................... 99
Tabla 24. Lista de especies y abundancia (cel./L) del fitoplancton en aguas libres de las lagunas estudiadas en el bajo Orinoco, sur de Monagas. .................................... 102
Tabla 25. Algunos índices de diversidad empleados para caracterizar la comunidad fitoplanctónica de lagunas de inundación del bajo Orinoco (sur de Edo. Monagas), para enero de 2010. Salida: PAST. ................................................................................... 105
Tabla 26. Índice de similitud de Jaccard para las lagunas estudiadas en el bajo Orinoco, sur del Edo. Monagas (enero de 2010). En rojo se resaltan los valores más altos. Salida: PAST. .................................................................................................. 106
Tabla 27. Composición y abundancia (células por litro) de la comunidad fitoplanctónica de tres ambientes de los módulos inundables de Mantecal (noviembre 2010). ......... 107
Tabla 28. Algunos índices de diversidad empleados para caracterizar la comunidad fitoplanctónica entres ambientes de los módulos inundables de Mantecal (Edo. Apure), en noviembre de 2010. Salida: PAST. ...................................................................... 108
Tabla 29. Índice de similitud de Jaccard para comparación de los tres ambientes estudiados en Mantecal (Edo. Apure). Salida: PAST. ............................................... 109

12
Resumen
Las poblaciones naturales suelen estar bajo presión ambiental constante, y sus
tamaños poblacionales están limitados por la disponibilidad de recursos. En el caso de
la ecuación logística, el parámetro que define la magnitud de la población en equilibrio
es la capacidad de carga (K), mientras que la dinámica de retorno al mismo, luego de
una perturbación, depende de la tasa de crecimiento per cápita (r). Las comunidades
están caracterizadas por atributos estructurales como diversidad y relaciones
interespecíficas. Las cianobacterias son los organismos fotosintéticos más antiguos
del planeta; se encuentran a mitad de camino entre las bacterias y las algas
eucariotas, pues su organización es procariótica pero su aparato fotosintético es
similar al de las algas. Arthrospira y Spirulina son dos géneros de cianobacterias
filamentosas que han colonizado diversos ambientes y algunas especies son
extremófilas y de distribución muy restringida. Estas especies son propias de
ambientes alcalinos (lagos de soda) y han experimentado pocas presiones
ambientales, entre ellas competencia y depredación escasas. Poseen un gran valor
alimenticio para animales y humanos, como lo demuestran numerosas
investigaciones. Del mismo modo, existen microalgas eucariotas que poseen gran
potencial para la biotecnología alimentaria y petrolera (bioenergética), esta última con
la finalidad de generar combustibles alternativos (biocombustibles), con valores
ecológicos y económicos para generar energía eléctrica y tracción para vehículos.
Esta investigación se enfocó en el estudio de las dinámicas poblacionales de A.
platensis (especie en revisión) y muestreos de comunidades fitoplanctónicas con
interés en especies autóctonas de gran valor biotecnológico. Se diseñaron
experimentos para optimizar medios de cultivos para A. platensis a escala pequeña en
sistemas controlados (Cámara de Crecimiento) y escalas mayores en sistemas no
controlados (Ficotrón). Se obtuvieron medios de cultivo idóneos para el crecimiento de
A. platensis en condiciones controladas (medio 1 = medio Parra (2005)) y naturales
(tratamiento 36: medio central), con miras a su aprovechamiento biotecnológico a gran
escala. La dinámica de crecimiento poblacional de A. platensis evidencia
densodependencia logística, con una estructura de tallas que varía a medida que la
población crece desde etapas tempranas hasta la capacidad de carga. Se encontraron
representantes de varios géneros y especies de cianobacterias (Arthrospira spp.,
Spirulina subsalsa, Lyngbya spp., Oscillatoria spp. y Anabaena spp.) y microalgas
eucarióticas (Scenedesmus spp., Isochrysis galbana, Chlorella sp., Chaetoceros sp.,
Navicula platalea, Dunaliella salina y D. viridis) de gran interés biotecnológico en
diferentes lugares de la geografía variada del país, lo que conduce a la idea de que
Venezuela cuenta con un gran potencial en su diversidad para la biotecnología de
microalgas.

13

14
1. Introducción
1.1 Una breve reseña histórica
Las referencias más antiguas del consumo de microalgas por el hombre datan del
Antiguo Testamento, puesto que el maná que permitió la supervivencia del pueblo de
Israel es un liquen del desierto (simbiosis de hongo y alga) (García-Blairsy 2008).
Spolaore y col. (2006) mencionan que el primer uso de las microalgas por los seres
humanos se remonta a 2000 años atrás en China; para entonces, los chinos utilizaban
Nostoc sp. para sobrevivir durante la hambruna. No obstante, no se conoce con
precisión cuándo el humano empezó a emplear las microalgas (Sánchez y col. 2003).
El uso corriente de estos recursos tiene tres precedentes: tradición, desarrollo
científico y tecnológico, y la denominada “tendencia verde” (Henrikson 1994). En la
América de la conquista europea, Bernal Díaz del Castillo, miembro de las tropas de
Hernán Cortés, reportó en 1521 que una pasta azulada (la hoy conocida Arthrospira
maxima) era cosechada del lago Texcoco, secada y vendida para consumo humano
en un mercado de Tenochtitlán (hoy Ciudad de México). Los aztecas dieron a este
alimento el nombre de tecuitlalt, el cual en su lengua literalmente significa
“excrementos de las piedras” e indiscutiblemente formó parte de su cultura alimentaria,
social, económica y política (Ciferri 1983, Sánchez y col. 2003).
Se sabe entonces que el empleo alimentario de microalgas por la humanidad no es
reciente. Hoy día, algunas culturas de la zona del lago Chad en África subsahariana,
como la etnia kanembu, conservan las mismas prácticas de cosecha artesanal de A.
platensis (identificación en revisión) legadas desde tiempo inmemorial. Esta
cianobacteria filamentosa constituye la base de la dieta diaria de esa tribu, la que
extraen del lago y secan al sol para preparar una galleta denominada dihé; este hábito
alimentario peculiar le ha brindado a los kanembu mejor estado de salud que tribus
vecinas que no consumen el dihé (Ciferri 1983). Cuando los científicos descubrieron la
rapidez con la que las poblaciones de estos microorganismos crecen, con un
rendimiento 20 veces mayor que la soja o soya (Glycine max) por unidad de superficie,
los describieron como el alimento del futuro (Henrikson 1994).
1.2 Evolución y ecología de las microalgas
Dentro del vasto grupo de seres vivos considerados bajo la denominación de
microalgas, éstas incluyen en su definición más amplia a las cianobacterias
(anteriormente cianofitas o cianofíceas y conocidas comúnmente como algas verde-
azules). Estos seres constituyen el grupo de organismos fotosintéticos más antiguos

15
del planeta y se puede decir que se encuentran a mitad de camino entre las bacterias
y las algas, porque su organización es procariótica pero su aparato fotosintético es
similar al de las algas (Prosperi 2000).
La Tierra tiene una edad aproximada de 4.600.000.000 de años y se ha podido
comprobar que 1.000.000.000 de años después de su formación ya había actividad
orgánica en la corteza terrestre. Los sedimentos no metamorfoseados más antiguos
de hace 3.500.000.000 de años muestran las primeras bacterias y los estromatolitos,
las más antiguas comunidades coloniales de las que se tenga conocimiento,
constituidas principalmente por algas verdes-azules filamentosas semejantes a las
Oscillatoriales (Orden al que pertenecen los géneros Oscillatoria, Lyngbya, Spirulina y
Arthrospira, entre otros), colonias que se asentaron en las costas de los mares
primitivos. Hoy sólo quedan estromatolitos vivos en la costa sur de Australia, las
Bahamas y algunas otras costas e islas remotas y prístinas (Woese 1987).
Hace unos 2.000.000.000 de años las cianobacterias produjeron suficiente oxígeno
para modificar sustancialmente la atmósfera terrestre. Muchos anaerobios obligados
(aquellos que no viven en presencia de oxígeno) fueron dañados por el oxígeno y
algunos desarrollaron modos de neutralizarlo, o se restringieron a vivir en áreas donde
este gas no penetra. Por selección natural algunos organismos aerobios se adaptaron
a vivir desarrollando una vía respiratoria que utilizaba el oxígeno para extraer más
energía de los alimentos y transformarla en ATP, prosperaron y radiaron en múltiples
formas de vida. La respiración aerobia se incorpora así al proceso anaerobio ya
existente de la glucólisis (Woese 1987, Mercado 1999).
Las cianobacterias y las microalgas eucarióticas al ser productoras que utilizan la luz
solar como fuente de energía contienen clorofila y otros pigmentos accesorios que les
otorgan una gran eficiencia fotosintética. Por el proceso de fotosíntesis que regula el
contenido de oxígeno y dióxido de carbono en la atmósfera, las microalgas contribuyen
notablemente a aliviar el efecto invernadero y se constituyen en protagonistas de la
producción inicial de materia viva en ecosistemas acuáticos (Mercado 1999).
La ecología de microalgas está determinada por un sinnúmero de factores ambientales
bióticos y abióticos que regulan sus poblaciones y determinan la amplitud de su
dispersión y la capacidad de invadir nuevos hábitats. Posiblemente el factor más
importante en la determinación de la abundancia del fitoplancton, comunidad acuática
errante constituida por microalgas, sea la disponibilidad de nutrientes. Las poblaciones
fitoplanctónicas aumentan sus números aceleradamente en la época de crecimiento

16
(surgencias, afloramientos, estación de mezcla en la zona limnética de los lagos, etc.),
por lo que ciertos nutrientes pueden hacerse limitados (Paulson 2005).
Otro factor primario a tomar en cuenta, principalmente en lo que respecta a la
distribución espacial del fitoplancton, es la luz, fenómeno físico que restringe a las
poblaciones de microalgas a parches en la zona iluminada (eufótica) de los cuerpos de
agua, único lugar donde pueden hacer la fotosíntesis. Si bien, las diferentes especies
de microalgas pueden entablar competencias intra e interespecíficas fuertes por la luz,
las comunidades fitoplanctónicas mantienen una riqueza elevada con tamaños
poblacionales moderados, fenómeno que ha sido denominado por Hutchinson (1961)
la paradoja del plancton.
De lo anterior se desprende la idea de que las poblaciones naturales suelen estar
siempre bajo presión ambiental constante, y sus tamaños poblacionales están
limitados por la disponibilidad de recursos. Aun cuando matemáticamente el
crecimiento poblacional es exponencial a densidades bajas, progresivamente se hace
densodependiente a medida que el tiempo avanza y la densidad crece. El modelo
logístico, el modelo densodependiente más conocido, explica el crecimiento de las
poblaciones durante lapsos más prolongados. Gráficamente se observa un periodo
inicial de latencia y adecuación (crecimiento lento), seguido de un incremento
exponencial (crecimiento rápido) para luego culminar en un plateau donde se alcanza
la capacidad de carga del ambiente, K (May 1976).
Las microalgas pueden ser organismos con un gran impacto negativo en la ecología
de los ambientes húmedos del planeta. Muchas especies de cianobacterias son
problemáticas en lagos y embalses usados para abastecimiento de agua para
consumo humano, donde crecen muy bien y frecuentemente producen florecimientos o
“blooms”, fenómenos en los que muchas especies se pueden acumular en las
espumas superficiales con densidades poblacionales sumamente altas. Estos
afloramientos son inducidos por enriquecimientos con macronutrientes como el fósforo
y el nitrógeno, principalmente derivados de actividades antrópicas (aguas ricas en
fertilizantes provenientes de zonas agropecuarias, vertidos industriales, aguas
domésticas, etc.); están acompañados de la producción de malos olores y sabores en
el agua, así como de neuro y hepatotoxinas mortales para animales domésticos y
seres humanos que la consumen (Carmichael 1994). En el mar ocurren las
denominadas mareas rojas, compuestas principalmente por dinoflagelados altamente
tóxicos, que producen una mortandad masiva de peces; también se dan las mareas
blancas o “blooms” en época de surgencia, congregaciones constituidas por diatomeas

17
que originan el efecto opuesto a las mareas rojas, pues desencadenan una gran
producción secundaria del zooplancton y de los siguientes peldaños de la cadena
alimentaria marina. Estas zonas de surgencia son de gran importancia en la economía
pesquera de muchas naciones del mundo.
1.3 Aspectos biogeográficos
En el contexto evolutivo y ecológico, Spirulina y Arthrospira son dos de los géneros de
cianobacterias más interesantes de este grupo antiguo, debido a la historia natural y
evolución singulares de algunas de sus especies en ambientes extremos y sumamente
restringidos, muchos de ellos agrestes e inhóspitos para casi cualquier otra forma de
vida, donde han tenido muy poca competencia y depredación. No obstante el carácter
extremófilo de ciertas especies de Spirulina y Arthrospira, estos géneros son ubicuos,
pues se encuentran representantes en ambientes marinos costeros, estuarinos,
humedales dulceacuícolas, lagos, etc., lo que también sugiere una idea del éxito
colonizador de estas cianobacterias filamentosas (Ciferri 1983).
Las microalgas extremófilas, como Arthrospira platensis, han evolucionado en sitios
que eran vastos en el precámbrico, y de los que ahora sólo quedan unos pocos sitios
aislados como relictos de esos mares antiguos por procesos continuos de evaporación
y desertificación. Estos procesos hoy día aún prosiguen y se intensifican con el
proceso de calentamiento global, como es el caso del lago Chad (África
Subsahariana), cuya superficie ha retrocedido en forma notable en el curso de menos
de un siglo. Dicho lago constituye uno de los últimos hábitats naturales de
microorganismos antiguos como A. platensis (Ciferri 1983).
De acuerdo al concepto de especie morfológica, la mayoría de las especies de algas
de agua dulce son cosmopolitas. Esto implica que las especies poseen mecanismos
muy eficientes para la dispersión o sus caracteres morfológicos han permanecido
“estáticos” en un tiempo evolutivo amplio. No obstante, hoy día muchos ficólogos
prestan atención a variaciones intraespecíficas en aspectos fisiológicos y bioquímicos.
Por otro lado, observaciones cuidadosas de poblaciones naturales pueden revelar
aislamiento reproductivo dentro de una morfoespecie o entre taxa intraespecíficos
(Ichimura 1996).
Se puede demostrar que los patrones de distribución de organismos sobre la
superficie del planeta no son aleatorios. Esta no aleatoriedad requiere explicaciones
en términos de procesos y reconstrucciones de eventos geológicos y biológicos en la
historia de la vida en la Tierra. Los procesos implican la formación de patrones

18
biogeográficos a partir de procesos abióticos muy lentos y a gran escala que incluyen
los movimientos tectónicos de placas, cambios en los niveles de los mares y océanos
y cambios climáticos, entre otros aspectos geológicos. Estos procesos han operado
casi en concierto, pues el clima puede ser afectado por los movimientos de los
continentes y los cambios en la circulación oceánica; los movimientos tectónicos
pueden alterar las corrientes oceánicas; el clima ha podido influir en la eustasia1 de la
periodicidad interglaciar. A un nivel más local, las erupciones volcánicas,
desertificaciones, cataclismos terrestres, huracanes, etc. también han contribuido a la
creación de patrones de distribución en diferentes hábitats (Myers y Giller 1991).
La determinación de patrones de distribución parte de un análisis biogeográfico. Tales
patrones son dóciles a análisis sin supuestos específicos de procesos subyacentes o
pueden ser usados para probar hipótesis sobre procesos. La biogeografía histórica y
ecológica indirectamente ha usado patrones observados de distribución de organismos
para probar hipótesis, con miras a explicar procesos tales como la vicariancia
(separación de grupos por barreras geográficas), dispersión, interacciones de especies
y eventos de perturbación (Myers y Giller 1991).
1.4 Importancia biotecnológica de las microalgas
Pocas personas, fuera del ámbito científico, conocen qué son las llamadas
cianobacterias, a pesar de que estos organismos son responsables tanto de beneficios
como de problemas en asuntos humanos (Prosperi 2000). Tampoco se conoce a la
mayor parte de las microalgas eucariotas, aún las bien sabidas propiedades abrasivas
de las tierras de diatomeas; o las especies que producen las perjudiciales mareas
rojas; o todavía sus “primas mayores”, las macroalgas, tan apreciadas en la
gastronomía japonesa y europea, así como en las industrias de geles como el agar y
carragenina, esta última para espesar jaleas y gelatinas. Son también invaluables los
aportes de las microalgas a la industria de los colorantes por la gran diversidad de
pigmentos que producen. Clorofitas del género Dunaliella acumulan elevados
contenidos de carotenoides, varias veces más que la zanahoria en base seca, por un
proceso denominado carotenogénesis, que las hacen fuente primaria de provitamina A
para humanos (Guevara y col. 2005).
1 El término eustasia se emplea para designar los hundimientos y posteriores ascensos de la corteza terrestre en aquellas zonas en que existieron grandes glaciares continentales durante el Pleistoceno. Con la formación de los grandes glaciares, el relieve se fue hundiendo por el peso del propio hielo y al finalizar el período glacial y desaparecer esos glaciares, el relieve previamente hundido tiende a "rebotar" hacia arriba debido a que se queda liberado de dicho peso (http://es.wikipedia.org/wiki/Eustasia).

19
A pesar del empleo milenario de las microalgas como alimento humano, la
biotecnología de estos microorganismos en realidad sólo comenzó a desarrollarse a
mediados del siglo pasado. Hoy en día, existen numerosas aplicaciones comerciales
de las microalgas. Por ejemplo, (i) las microalgas pueden ser utilizadas para mejorar el
valor nutritivo de los alimentos y la alimentación animal debido a su composición
química, (ii) desempeñan un papel crucial en la acuicultura y (iii) pueden ser
incorporadas a los cosméticos. Además, se cultivan como fuentes muy valiosas de
diversas moléculas. Respecto a esto último, por ejemplo, los ácidos grasos
poliinsaturados de los aceites se añaden a las fórmulas infantiles y complementos
nutricionales y los pigmentos son importantes como tintes naturales. Isótopos
bioquímicos estables ayudan en la determinación estructural y los estudios
metabólicos. La investigación futura deberá centrarse en la mejora de los sistemas de
producción y la modificación genética de las cepas. Las microalgas constituyen de
esta manera una alternativa económica cada vez más diversificada y competitiva
(Spolaore y col. 2006).
Entre los beneficios que la humanidad obtiene de las cianobacterias, por citar uno de
múltiples ejemplos, se encuentra su utilización como biofertilizantes en agricultura,
gracias a su capacidad de fijar nitrógeno atmosférico (N2) y producir compuestos
orgánicos ricos en nitrógeno que contribuyen a incrementar los rendimientos de las
cosechas de arroz (Prosperi 2000). El empleo de algunas especies de los géneros
Spirulina y Arthrospira para la alimentación humana está ampliamente comprobado
por un sin fin de investigaciones científicas, las que coinciden en su inocuidad (no son
tóxicas), gran palatabilidad (más de 90% comestible) y sus importantes cantidades de
proteínas (65-70% en base seca), vitaminas y otros nutrientes (Ciferri 1983, Henrikson
1994, Mani y col. 2000, Pervushkin y col. 2001, Sachdeva y col. 2004).
Está ampliamente comprobado el beneficio alimentario de Spirulina y Arthrospira en
animales de granjas pecuarias (aves de corral, porcinos, etc.) y acuícolas (peces,
camarones, etc.), y consumo humano (Martínez-Palacios y col. 1996, Soler y col.
2000, Nandeesh y col. 2001, Jaime-Ceballos y col. 2004). Las tecnologías empleadas
para cultivar masivamente microalgas y obtener productos alimenticios, farmacéuticos
y cosméticos de su biomasa son limpias en su totalidad; las granjas a cielo abierto
tienen un gran valor ecológico como trampas vivientes de gases de efecto invernadero
y purificación de biogás (Conde y col. 1993), así como recuperación de cuerpos de
agua contaminados (Ayala y Vargas 1987, Sim y Goh 1988, Cañizares y col. 1993).

20
Existen diversas especies de microalgas que metabolizan hidrocarburos, los
bioacumulan en sus vacuolas y producen aceites, a partir de los cuales se pueden
producir biocombustibles (biodiesel). La clorofita Botryococcus braunii se caracteriza
por su capacidad de producir hidrocarburos insaturados de cadena larga, llegando a
contenidos que van de 15 a 75% de su peso seco. Además, esta microalga produce
polisacáridos extracelulares que inducen la formación de colonias, el tamaño de las
mismas depende de la hidrodinámica de estrés en el biorreactor. Su gran potencial
como fuente renovable de combustibles de base o de productos químicos ha sido
demostrado por diferentes grupos de investigación (Casadevall y col. 1985, Banerjee y
col. 2003).
Otras especies de microalgas con gran potencial en biotecnología petrolera, debido a
sus gran contenido de lípidos, son Schizochytrium sp., Nannochloropsis sp., Nitzschia
sp., Isochrysis sp. y Tetraselmis suecica, entre otras (Tabla 1).
Tabla 1. Especies de microalgas con potencialidades para generación de biocombustibles. Contenido de aceites en base seca (tomada de Albarracín 2007).
Especie Contenido de aceite (% de peso seco)
Botryococcus braunii 25 – 75
Chlorella sp. 28 – 32
Crypthecodinium cohnii 20
Cylindrotheca sp. 16 – 37
Dunaliella primolecta 23
Isochrysis sp. 25 – 33
Monallanthus salina 20
Nannochloris sp. 20 – 35
Nannochloropsis sp. 31 – 68
Neochloris oleoabundans 35 – 54
Nitzschia sp. 45 – 47
Phaeodactylum tricornutum 20 – 30
Schizochytrium sp. 50 – 77
Tretraselmis suecica 15 – 23

21
Por su parte, la diatomea marina Chaetoceros muelleri puede tener potencial para su
explotación como un precursor renovable de los combustibles líquidos o como fuente
de lípidos, esto sobre la base de su alta tasa de crecimiento, la tolerancia a una amplia
gama de temperaturas y conductancias específicas, y una gran cantidad de lípidos
intracelulares (Mcginnis y col. 1997).
El fitoplancton ha sido ampliamente estudiado en sus dinámicas poblacionales y
comunitarias. Los estudios teóricos a partir de observaciones empíricas y de campo
han crecido notablemente debido a la importancia de muchas especies de microalgas
en la alimentación, farmacia, medicina, control de calidad de aguas, etc. Un aspecto
aún incipiente es el estudio biogeográfico de microalgas en sus ambientes nativos.
Este trabajo trata de dos temas: (1) estudio de las dinámicas poblacionales en
especies cultivadas y (2) muestreo y caracterización de hábitats y aspectos
comunitarios de especies autóctonas de gran valor en la biotecnología alimentaria y
petrolera.

22
2. Antecedentes en Venezuela
En Venezuela se han hecho numerosos trabajos taxonómicos y ecológicos de
microalgas. Diversas especies fitoplanctónicas de importancia biotecnológica son
nativas de Venezuela; las clorofitas Botryococcus braunii y Chlorella vulgaris y
cianobacterias del género Spirulina han sido reportadas para el embalse de Guri
(González de Infante y Riehl 1992). Anteriormente, B. braunii también se encontró en
la laguna de San Javier del Valle, Edo. Mérida (Yacubson 1974). En aguas salobres y
estuarinas, como el sistema del lago de Maracaibo (norte del lago) se ha reportado la
cianobacteria Spirulina subsalsa y otras cianobacterias eurihalinas (Rodríguez 2001).
En diversos sistemas marinos costeros y lagunas hipersalinas del país se han
colectado diferentes especies de clorofitas del género Dunaliella (Guevara y col.
2005), así como prasinofitas del género Tetraselmis (Romero y col. 2002), entre otras
especies.
Por otra parte, algunas investigaciones se han orientado a ensayos de cultivos de
microalgas en condiciones controladas con fines biotecnológicos. Algunos análisis han
permitido la determinación de las composiciones nutricionales y producciones de
metabolitos de diferentes especies de microalgas: Anabaena sp. (Morales y col. 2002),
Chlorella sp. (Mora y col. 2004) y Dunaliella salina (Guevara y col. 2005). Otros
ensayos han conducido a la cuantificación de efectos inmunomoduladores de Spirulina
subsalsa (Cheng-Ng y col. 2005). Parra (2005) aisló y purificó c-ficocianina y
aloficocianina de Arthrospira platensis, dos pigmentos de gran interés en la industria
de los colorantes y marcadores moleculares. Naranjo y col. (2010) hicieron una
revisión del uso de A. platensis como biofactoría de metabolitos secundarios de interés
farmacológico, con énfasis en el ácido pipecólico.
La Asociación Civil Gente de Ciencias (2007), organización científica y social
venezolana constituida por profesionales del área de las ciencias naturales, con
experiencia en el cultivo de microalgas (cultivos en laboratorio y campo; optimización
de medios de cultivo, fotobiorreactores y obtención de productos de microalgas), con
énfasis en Arthrospira platensis desde el año 2000. Las investigaciones se
concentraron en la aplicación de la biotecnología de microalgas en temas como la
nutrición animal y humana. Recientemente con Botryococcus braunii, Nannochloropsis
sp., Tetraselmis chuii e Isochrysis galbana, entre otras especies, se ha explorado la
factibilidad de producir biocombustibles a partir de aceites sintetizados o acumulados
en sus compartimentos intracelulares.

23
3. Justificación
Este trabajo está enmarcado en el estudio ecológico, taxonómico y biogeográfico de
especies de microalgas eucarióticas y cianobacterias autóctonas con potencialidades
para su cultivo masivo en Venezuela y empleo en la acuicultura, biotecnología
alimentaria, generación de biocombustibles (biodiesel), biorremediación, producción de
energías limpias y trampas de gases de efecto invernadero. También se incorporan
especies cultivadas en Venezuela desde hace varios años, como Arthrospira platensis
y A. maxima de conocido perfil nutricional, médico y biorremediador, por lo que tienen
gran demanda mundial; más de 70 países las cultivan comercialmente (Henrikson
1994).
Las especies de los géneros Spirulina y Arthrospira han sido incorporadas con éxito
notable en programas sociales y alimentarios dentro de países del cuarto mundo con
altas tasas de desnutrición, como el África Ecuatorial. Conviene entonces incorporar el
estudio ecológico de estas cianobacterias para conocer sus tasas de crecimiento
óptimas y otros parámetros poblacionales con miras a cultivarlas a gran escala en el
país y aprovechar su biomasa con la firme intención de establecer programas
nutricionales similares en Venezuela y Latinoamérica, a sabiendas de los niveles
preocupantes de desnutrición que imperan en los países de esta región.
El estudio de los organismos en su ambiente natural brinda la oportunidad de conocer
las especies con las que cuenta Venezuela y adquirir un conocimiento integral de sus
ambientes naturales. Esta propuesta lleva implícito un propósito productivo de emular
las condiciones naturales para lograr sus cultivos en sistemas controlados, y una
finalidad de conservación a partir del conocimiento de los ambientes para manejo
sustentable de ecosistemas o su protección como santuarios de vida. En síntesis, la
meta de esta investigación es brindar puntos de partida a partir de un estudio
ecológico integral a expectativas y necesidades creadas en el país por la búsqueda de
fuentes alternativas de alimentos, medicinas, pigmentos y energías limpias, así como
perspectivas para la recuperación de ecosistemas acuáticos contaminados. Esta
propuesta está enmarcada en el Plan Nacional Simón Bolívar como nuevo modelo
sociopolítico para el país.

24
4. Hipótesis
1. Las poblaciones de Arthrospira platensis cultivadas en el laboratorio presentan
densodependencia, y un crecimiento poblacional que puede ser descrito con
modelos matemáticos.
2. La geografía y ecología variadas del país permiten esperar una gran diversidad de
especies de microalgas distribuidas en forma discontinua y gradientes, algunas de
ellas con gran potencial biotecnológico.

25
5. Objetivos
1. Establecer la dinámica de crecimiento poblacional de Arthrospira platensis con
optimización de medios de cultivo para fines productivos.
2. Caracterizar hábitats y comunidades fitoplanctónicas con especies de interés
biotecnológico en el país.
Del objetivo 1 se desprenden los objetivos específicos:
a. Caracterizar el crecimiento poblacional de A. platensis en condiciones
controladas y naturales.
b. Determinar los parámetros poblacionales.
c. Realizar análisis de perturbación en el modelo dinámico.
Del objetivo 2 derivan los objetivos específicos:
a. Identificar especies de microalgas de muestras colectadas en el campo.
b. Determinar riqueza, abundancia e índices de diversidad.
c. Establecer relaciones entre ambientes, variables físicas y químicas y la
composición de especies encontradas.

26
6. Metodología
El plan general de trabajo se dividió en dos niveles de organización: poblacional y
comunitario.
6.1 Ecología de poblaciones de especies cultivadas
Los estudios poblacionales se condujeron por aproximaciones experimentales y con
base en la teoría ecológica. Los mismos se centraron en cultivos de la cianobacteria
Arthrospira platensis (cepa Lefevre 1963/M-132-1, República Checa). La especie se ha
cultivado en el Laboratorio de Optimización Agrícola (LOA-IZET) por más de ocho
años. Arthrospira maxima (cepa cubana), es una especie congénere obtenida para el
cepario del laboratorio en tiempo más reciente. El cepario también cuenta con otras
especies de valor biotecnológico provenientes de otros ceparios del país y del
extranjero, así como colecciones de campo: Chaetoceros sp. (Bacillariophyta),
Botryococcus braunii (Chlorophyta), Chlamydomonas sp. (Chlorophyta), Chlorella
vulgaris (Chlorophyta), Dunaliella salina (Chlorophyta), D. viridis (Chlorophyta), D.
primolecta (Chlorophyta), Haematococcus pluvialis (Chlorophyta), Scenedesmus sp.
(Chlorophyta), Spirulina subsalsa (Cyanobacteria), Isochrysis galbana (Haptophyta),
Nannochloropsis sp. (Ochrophyta), Tetraselmis sp. (Prasinophyta), Porphyridium
cruentum (Rhodophyta) y Rhodosorus marinus (Rhodophyta).
Medios de cultivo. Se procedió a la preparación de un medio de cultivo estándar para
cianobacterias filamentosas alcalófilas de los géneros Spirulina y Arthrospira
denominado medio Spirulina o SAG (Schlösser 1994), que es una modificación del
medio Aiba y Ogawa (1977). Parra (2005), incorporó algunos cambios a este medio
nutritivo. La composición del medio Spirulina se presenta en la Tabla 2.
Tabla 2. Composición química del medio Spirulina (Schlösser 1994).
Componentes Solución Stock
(mL) Masa (g)
Concentración
en el medio
final
pH
Solución I 500 (2X) – – Alcalino
(>9)
NaHCO3 – 13,61 1,62×10-4
M –
Na2CO3 – 4,03 3,80×10-5
M –
K2HPO4* – 0,50 2,87×10-4
M –
Solución II 500 (2X) – – Neutro
(7)
NaNO3* – 5,00 2,94×10-5
M –
K2SO4 – 2,00 5,74×10-6
M –
NaCl – 2,00 1,71×10-5
M –

27
Tabla 2. Continuación.
Componentes Solución Stock
(mL) Masa (g)
Concentración
en el medio
final (1X)
pH
MgSO4.7H2O – 0,40 8,11×10-7
M – CaCl2.2H2O – 0,02 2,72×10
-7M –
FeSO4.7H2O – 0,02 3,60×10-8
M – Na2EDTA.2H2O – 0,16 2,15×10
-7M –
Traza metálica 1000 (1000X) – – Neutro
(7)
Na2EDTA.2H2O – 0,80 2,15×10-6
M – FeSO4.7H2O – 0,70 2,52×10
-6M –
ZnSO4.7H2O – 0,001 3,48×10-9
M – MnSO4.7H2O – 0,002 8,97×10
-9M –
H3BO3 – 0,01 1,62×10-7
M – Co(NO3)2.6 H2O – 0,001 3,44×10
-9M –
Na2MoO4.2 H2O – 0,001 4,13×10-9
M – CuSO4.5H2O – 0,00005 2,00×10
-11M –
*Parra (2005) reemplazó el K2HPO4 por KH2PO4, debido a la mayor solubilidad de la primera
en agua. También sustituyó el NaNO3 por KNO3 para obtener una relación Na:K más próxima a la fisiológica (4:1).
Spirulina subsalsa fue cultivada en medio Spirulina combinado con agua de mar
(50:50), debido al origen estuarino de la cepa (Morales, com. pers.). El resto de las
especies de microalgas del cepario se cultivaron en diferentes medios probados y
estandarizados como óptimos para sus crecimientos poblacionales: medio F/2
(Guillard 1975), para microalgas marinas como Nannochloropsis sp., Tetraselmis sp. e
I. galbana; medio Algal (Fábregas y col. 1985), el cual fue empleado tanto para
microalgas de agua dulce como Chlamydomonas sp., C. vulgaris y Scenedesmus sp.,
como para diatomeas marinas (Chaetoceros sp.), con adición de silicatos, rodofitas (P.
cruentum y R. marinus) y Tetraselmis sp., con adición de agua de mar, y clorofitas de
ambientes hipersalinos (Dunaliella spp.), con la incorporación de solución saturada de
cloruro de sodio (NaCl); medio Chu-13 (Chu 1942) modificado por Dayananda (2007)
para B. braunii; Haematococcus pluvialis fue cultivada en medio Bristol (Bold 1949).
Para especies del género Dunaliella también se probó el medio de Serpa y Calderón
(2006). Las composiciones de estos medios se presentan en el Apéndice 1.
La preparación de los medios se efectuó conforme los siguientes pasos generales:
1. Se preparó una solución madre 1000X (solución concentrada) de micronutrientes
(oligoelementos o traza metálica), según lo establecido en cada medio de cultivo.
Para ello, las sustancias requeridas como sales inorgánicas cristalizadas grado
analítico se pesaron en una balanza analítica digital AND (d = 1 mg).

28
2. Las sustancias constituyentes de la solución de macronutrientes, sales inorgánicas
cristalizadas grado analítico (salvo bicarbonato y carbonato de sodio, sales que
debido a sus altos requerimientos para el medio Spirulina se obtuvieron de sacos
de 25 ó 50 kg de agroquímicos grado técnico), también se pesaron en una balanza
analítica digital AND (d = 1 mg). Las sales de bicarbonato y carbonato de sodio se
pesaron en una balanza digital de 10 kg (d = 1 g).
3. Cada sal se disolvió en forma independiente con agitación magnética y calor, luego
se mezclaron todos los componentes de la solución madre y se enrasó al volumen
final, generalmente de un (1) litro, en un balón aforado o en un cilindro graduado
de 1 L. Algunas sales, como el sulfato ferroso heptahidratado (sal utilizada
generalmente como fuente de hierro para todos los medios), tuvieron que ser
disueltas con un quelante (EDTA disódico o citrato trisódico dihidratado). No se
añadieron vitaminas ni carbohidratos a ninguna solución de medio nutritivo.
4. La solución madre de micronutrientes se esterilizó en un autoclave analógico
vertical KALSTEIN durante 45 min hasta una temperatura tope de 119ºC
(temperatura óptima = 121ºC), y una presión de 10 PSI (presión óptima = 15 PSI).
Antes de autoclavar cualquier solución y material (pipetas Pasteur, fiolas, botellas
vacías, etc.), una cinta de papel autoadhesivo indicador (o testigo) se colocó a
cada recipiente.
5. A la solución de macronutrientes se añadió la alícuota correspondiente de solución
madre de micronutrientes estéril, para de este modo obtener el medio de cultivo
íntegro. Para trasvasar soluciones se tomó la previsión de destapar el envase
frente a la llama azul de un mechero FISHER.
6. Finalmente, la solución de medio de cultivo se autoclavó para luego enfriar y
proteger con papel envolvente.
Inóculo. Se obtuvieron cultivos monoalgales y monoclonales (poblaciones surgidas de
un filamento) de A. platensis (cepa Lefevre), por la virtud de estas cianobacterias de
reproducirse asexualmente por particiones de filamentos o tricomas en sitios críticos
que necrosan y se escinden formando hormogonios (filamentos reproductivos), sitios
denominados necridia o necridios. Esto garantizó uniformidad genética en las
poblaciones de estos microorganismos.
Para la obtención de cultivos monoclonales se cumplió la pauta siguiente:

29
1. De una cepa se aisló un filamento en una cápsula de Petri con solución buffer
alcalina (bicarbonato-carbonato de sodio, 4:1) bajo un estereoscopio (lupa).
2. Cada filamento se colocó en un tubo de ensayo con medio Spirulina esterilizado y
se colocó en cámara de crecimiento (T = 27ºC) con iluminación permanente. El
periodo de incubación fue de 15 días, tiempo en el cual el filamento inicial pudo
clonarse en varios filamentos vía escisión en necridios.
3. Los cultivos monoclonales se escalaron a fiolas de 250 mL y se mantuvieron sin
agitación en la cámara de crecimiento. En estos recipientes se mantuvieron para
ser empleados en medios sólidos y completar proceso de purificación de la cepa.
Para la siguiente etapa de purificación del cultivo monoalgal y monoclonal, con la
finalidad de controlar contaminación de bacterias y hongos, se emplearon los
siguientes pasos:
1. Se prepararon medios sólidos con disolución en caliente de agar no purificado en
medio Spirulina (1,5 g de agar por cada 100 mL de medio), de esta manera se
elaboró un medio de agar alcalino.
2. Las soluciones con agar se autoclavaron en cápsulas de Petri, para hacer placas y
en tubos de ensayo inclinados para obtener cuñas. Una vez obtenidos los medios
sólidos estériles, se procedió a encender la llama azul en un mechero FISHER y
limpiar el mesón con etanol absoluto. Este procedimiento se hizo con el fin de
tener las mayores condiciones de asepsia posibles.
3. Bajo estas condiciones de asepsia y llama azul, una vez que las soluciones
esterilizadas se enfriaron, se procedió a la siembra de las cepas vivas. Los medios
solidificados en placas y tubos fueron sembrados por medio de un asa metálica
pasada por llama azul y enfriada en algún punto del medio sólido, teniendo
cuidado de no inocular posteriormente en esa zona (figura 1).
Figura 1. Esquema ilustrado donde se muestra el escalamiento de microalgas de medio líquido a medio sólido con el empleo de la técnica del asa de estaño.

30
4. Finalmente las cepas en las soluciones nutritivas solidificadas se ubicaron en el
cepario o cámara fría bajo luz fría (tubos fluorescentes day-light) a una
temperatura media de 23ºC.
Luego de la obtención de cepas limpias, monoclonales y monoalgales, se procedió al
escalamiento a fiolas de 250 mL, y de éstas a fiolas de mayores volúmenes en forma
sistemática hasta llegar a los cultivos en fiolas de 2 L con agitación y sin fotoperiodo
(luz por 24 h diarias). Para ello se procedió del modo siguiente:
1. Soluciones líquidas esterilizadas se trasvasaron a fiolas de 250 mL cubiertas con
tapones de algodón y gasa. Todo este material previamente se autoclavó. Los
medios líquidos nuevos se inocularon con pequeños volúmenes de cepas
provenientes de cuñas y placas del cepario en cámara fría.
2. Las cepas cultivadas en medio líquido se colocaron en la cámara de crecimiento
bajo luz fría y temperatura media de 27ºC, con agitación y luz continuas. La
agitación en la cámara de crecimiento la proveyó un blower KAWAKE (¼ HP; 220
V), y se condujo a través de un sistema de tubos y mangueras hasta culminar en
pipetas Pasteur esterilizadas sumergidas en las soluciones con inóculo.
3. Los dos pasos anteriores se repitieron para los escalamientos sucesivos hasta
culminar en cultivos a escala mayor (figuras 2 y 3).
Figura 2. Esquema donde se muestra el escalamiento de poblaciones de microalgas a fiolas de diferentes volúmenes hasta alcanzar los cultivadores a gran escala.
Diseño de experimento. Los experimentos en laboratorio bajo condiciones
controladas se ejecutaron a escala menor (fiolas de 2 L); estas pruebas se ejecutaron
en la cámara de crecimiento del cepario del Laboratorio de Optimización Agrícola
(LOA). Los ensayos en campo se llevaron a cabo a escala mayor y con monitoreo de
las condiciones ambientales en la Planta Experimental “Ficotrón” ubicada en el
Instituto de Estudios Avanzados (IDEA), Sartenejas, Edo. Miranda. Se ensayó en

31
botellones de 5 L, tanques cilíndricos de 500 L y cultivadores tipo carrusel (en inglés
“race-ways”) de 2000 L (figura 3).
Figura 3. Imágenes donde se muestran las diferentes etapas de escalamiento del cultivo en medio líquido en el sistema integrado LOA-Ficotrón: (a) cultivos a escala pequeña en condiciones controladas (Cámara de Crecimiento, LOA - IZET); (b), (c) y (d) cultivos a cielo abierto en el Ficotrón, IDEA, en botellones de 5 L, tanques circulares de 500 L y cultivadores tipo carrusel de 2000 L, respectivamente.
Cada diseño de experimento se definió por el número de factores o variables
independientes a fijar y su número de niveles (Montgomery 2001). En todos los
experimentos se consideraron variables dependientes: (1) la densidad poblacional
(filamentos/L), por el método directo de conteo de filamentos, y (2) la absorbancia a
longitud de onda (λ) = 680 nm, para el método indirecto.
Los ensayos de laboratorio se organizaron en diseños factoriales 22 (dos factores,
sodio (Na) y potasio (K), con dos niveles, alta y baja concentración), para comparar
cuatro medios de cultivo (modificaciones del medio Spirulina) empleados para A.
platensis. Los ensayos no se replicaron en número de fiolas, puesto que se supone la
existencia de repeticiones del experimento en cada una de las posibles combinaciones
de los niveles del factor correspondiente (Montgomery 2001). Se hicieron ajustes
matemáticos para modificar la relación Na+:K+, sin variar en forma notable las
concentraciones de los demás iones. Con estas pruebas se determinó el medio de
cultivo óptimo para el crecimiento poblacional en condiciones controladas, a partir de
los dos cationes más importantes en el equilibrio electroquímico entre los
compartimentos intra y extracelulares, y contraiones de las sales inorgánicas. Las
interacciones entre factores se evaluaron con un ANOVA de dos vías (α = 0,05). La
Tabla 3 muestra el resumen del ensayo factorial 22.
Tabla 3. Diseño factorial 22 para ensayos de laboratorio. El experimento se simplificó a cuatro
medios de cultivo sin replicación.
K+ alto K+ bajo
Na+ alto + + + –
Na+ bajo – + – –
(a) (b) (c) (d)
32
En campo se continuó la investigación con diseños factoriales de mayor complejidad, a
medida que se añadieron factores. El primer experimento de campo se llevó a cabo en
botellones plásticos de 5 L y el diseño correspondió a un factorial fraccionado 26-1 de
resolución VI, montado con el programa Design Expert versión 6, donde las variables
independientes o factores fueron seis sales grado alimenticio a dos niveles, con cinco
puntos centrales. El programa produjo 37 tratamientos, 5 centrales y el resto
combinaciones aleatorias de los factores (Tabla 4). Las sales forman parte del medio
Spirulina (Schlösser 1994), pero en lugar de la sustitución de Parra (2005) de KNO3
por NaNO3, se empleó NH4NO3, esto debido a razones de costos. A esta escala de
experimentación se emplearon sales grado alimenticio y no reactivo, el NH4NO3 resultó
la fuente de nitrógeno disponible y muy empleada en producción de fertilizantes.
Tabla 4. Resumen del diseño factorial fraccionado 26-1
(salida del programa Design Expert)*.
Study Type: Factorial Runs: 37
Initial Design:
2 Level Factorial Blocks: No Blocks
Center
Points:
5
Design Model:
Reduced 3FI
Factor Name Units Type Low Actual
High Actual
Low Coded
High Coded
Mean Std. Dev.
A NaHCO3 mg/L Numeric 6805 13610 -1 1 10207,5 3164,3
B Na2CO3 mg/L Numeric 2015 4030 -1 1 3022,5 937,0 C K2HPO4 mg/L Numeric 250 500 -1 1 375 116,2
D NaCl mg/L Numeric 500 1000 -1 1 750 232,5
E K2SO4 mg/L Numeric 500 1000 -1 1 750 232,5
F NH4NO3 mg/L Numeric 1176 2352 -1 1 1764 546,8
*Para más detalle sobre este ensayo, ver Apéndice 2.
Las variables respuestas o dependientes aleatorias fueron: absorbancia a 680 nm,
número de filamentos por litro, y biomasa final (g/L). El medio óptimo para este
experimento se escogió por una técnica matemática multivariada denominada
superficie de respuesta (ver Apéndice 2 con salida del análisis en Design Expert, y
Apéndice 3 que muestra un marco teórico breve que explica la técnica).
Para este ensayo de campo, los medios se prepararon según esquema de la figura 4.
Los pasos se enumeran a continuación:
1. Se prepararon soluciones stocks por triplicado con agua destilada y se preservaron
en envases limpios. Los primeros ocho tratamientos se prepararon en dos
soluciones de 500 ml cada una: Solución I (Stocks A, B y C) y Solución II (stocks I,

33
D, E, F y II). Las preparaciones se hicieron en grupos de ocho debido a la
capacidad limitada del autoclave (16 botellas de vidrio de 1 L c/u).
2. Se autoclavaron las soluciones I y II de cada tratamiento.
3. Cada tratamiento se aforó a 4 L con agua filtrada.
4. Se tomó una muestra de 50 mL de cada tratamiento y se realizaron medidas de pH
y conductividad como método de comprobación de calidad del medio.
Figura 4. Esquema para la preparación de cada medio (tratamiento) del diseño factorial fraccionado 2
6-1.
La siguiente escala de volumen en el estudio, la constituyeron tanques cilíndricos de
500 L. El diseño experimental empleado para evaluar el crecimiento poblacional de A.
platensis en este tipo de cultivadores fue un factorial 32 con bloques completos
aleatorizados. Se estableció un sistema de 9 tanques cilíndricos de 500 litros de
polietileno de alta densidad, con sistema de bombeo y aireación (blower KAWAKE ¼
HP; 220 V) bajo invernadero (Ficotrón). El sistema de bombeo está compuesto de dos
bombas de pecera (cada bomba aproximadamente suministra 2 L•min-1). Cada medio
en tanque se inoculó con 10% de los volúmenes totales, dando como resultado una
concentración promedio de 83 mg/L. Las pérdidas por evaporación se repusieron con
agua filtrada. Cada 72 h se midió la absorbancia o densidad óptica (DO) en un
espectrofotómetro SHIMADZU UV-160 a λ = 680 nm. Como medio de cultivo se
empleó el optimizado en el diseño factorial fraccionado 26-1. La variable independiente
aleatoria fue la profundidad (cm), debido a que este factor físico resulta determinante
Stock A
1/2L Sol. I
de Trat. X
1/2L Sol. II
de Trat. X
4L Trat. X
Autoclavar I y II
Stock B Stock C Stock I Stock D Stock E Stock F Stock II

34
en las tasas de crecimiento y mortalidad de filamentos por la generación de puntos
oscuros donde la fotosíntesis se anula. Las variables respuestas a evaluar fueron la
absorbancia a 680 nm (medida cada tres días) y biomasa total (g/L), ésta última
determinada una vez realizada la cosecha al final del periodo de incubación. La figura
5 muestra un esquema de este sistema de tanques y su disposición espacial para el
experimento.
Figura 5. Esquema de la disposición espacial de los tanques cilíndricos en el Ficotrón para la ejecución del diseño de bloques completos aleatorizados.
Un Análisis de Varianza (acrónimo en inglés ANOVA) de dos vías (α = 0,05) se realizó
para determinar efectos significativos de tratamientos y bloques. La corrida de la
prueba se ejecutó en EXCEL versión 2007.
Finalmente, para piletas o cultivadores tipo carrusel de 2000 L no se realizó un diseño
experimental formal, pues sólo se dispuso de un sistema con agitación por paletas de
los ocho construidos en la planta. De esta forma, no se dispuso de “réplicas” para
comparar medias poblacionales de A. platensis con diferentes tratamientos. Sólo se
pudo recurrir a un muestreo sin norma o puntual cada tres días para valorar la
absorbancia a 680 nm y biomasa final (g/L), una vez realizada la cosecha.
Biomasa. Para la obtención de biomasa seca, en laboratorio se procedió a colectar la
biomasa húmeda en una malla de poliéster con 27 μm de apertura de poro, mientras
que en el campo (Ficotrón) la cosecha se hizo con filtros metálicos con diámetro de
poro de 55 μm. El pH del medio de cultivo de A. platensis se caracteriza por su
60 cm 60 cm
60 c
m6
0 c
m
ABC
1
2
3
N
S
BLO
QU
ES
F I L A S
35 cm
35 cm
35 cm 25 cm
25 cm
25 cm 15 cm
15 cm
15 cm
A1-25A2 25
A3 15
B1 35
B2 15
B3 25C1 15C2 35
C3 35
10º24`12`` N 66º53`13`` O

35
alcalinidad elevada (9,5 a 10), por lo que la biomasa fue neutralizada con agua filtrada
o agua acidulada para fines de aprovechamiento alimenticio; un pH-metro digital
HANNA permitió monitorear la reducción del pH hasta 7. Finalmente, la biomasa
húmeda y neutralizada fue extendida sobre una superficie de papel parafinado y
colocada en rejas metálicas, para ser secada en un deshidratador a 60ºC por 24 h. La
biomasa total o final se estableció una vez alcanzado el plateau y sucedidos algunos
días para la estabilización de la curva (de 15 a 30 días de incubación).
Determinación de la densidad poblacional. Se determinó el crecimiento poblacional
de A. platensis por dos métodos: (1) directo, por conteo de filamentos en cámaras
cilíndricas desmontables Utermöhl (1958) bajo un microscopio invertido LEICA DMIL, y
(2) indirecto, por absorbancia o densidad óptica en un espectrofotómetro digital
SHIMADZU UV-160 a longitudes de onda (λ) de 680 y 700 nm, correspondientes al
rojo y rojo lejano, respectivamente, en las cuales la clorofila a teóricamente tiene
absorción óptima de luz visible y umbral visible-infrarrojo.
Para el conteo de filamentos, se empleó el método de conteo de bandas (Utermöhl
1958) en una cámara de 25 mL. Previamente, las muestras se dejaron sedimentar en
la cámara por 24 h. Una vez hecho el cómputo de filamentos por banda, se determinó
la densidad ( D ) o número de filamentos por litro a partir de la ecuación:
1000/Lfilamentos
NVTL
SD (1)
Donde:
S = superficie total de la cámara de sedimentación (2r )
N = número total de filamentos contados
L = longitud de la banda recorrida (2 r )
T = ancho de la banda recorrida (según aumento del objetivo)
V = volumen total de la cámara (25 mL)
Para determinar el ancho de la banda, se calibró el microscopio con una escala
grabada en un ocular (aumento 10x) dividida en 100 líneas, la cual se superpuso a
otra escala de 2 mm de longitud grabada en un portaobjetos ubicado sobre el carro.
Para el aumento de cada objetivo se obtuvieron los siguientes factores de conversión:

36
10x: 3 líneas = 0,05 mm
20x: 10 líneas = 0,08 mm
32x: 20 líneas = 0,1 mm
Previo a cada ensayo con el método indirecto, en el espectrofotómetro se hizo un
barrido (en inglés “scan”) de absorbancias, en un intervalo de longitudes de onda de
400 a 700 nm, con factibilidad de expandir hacia el espectro UV por el límite inferior o
el infrarrojo por el límite superior; de esta manera se verificaron los máximos u óptimos
de absorción de las cepas de A. platensis.
La densidad óptica ( λDO ) es la absorción de un elemento óptico por unidad de
distancia, para una longitud de onda dada:
I
Ilog
lTlog
ll
ADO λ
λ0
1010
11 (2)
Donde:
Aλ = Absorción a longitud de onda λ
l = Distancia que la luz viaja por una muestra (i.e., el grosor de la muestra, o dicho de
otro modo, anchura o abertura de la cubeta, con exclusión de las paredes) medida en
centímetros (cm). Si l = 1 cm, entonces λDO = λA
T = Transmitancia por unidad
I0 = Intensidad del rayo de luz incidente
I = intensidad del rayo de luz transmitido
(http://es.wikipedia.org/wiki/Densidad_óptica)
Mientras más alta es la densidad óptica, más corta es la transmitancia. Esta última
medida está acotada entre 0 y 1, si se transforma a porcentaje (como generalmente se
registra en espectrofotómetros), entonces la transmitancia se lee entre 0 y 100%. Para
calibrar el espectrofotómetro, se empleó una solución “blanco”, la cual teóricamente
tiene 100% T o Aλ = 0, y la misma se restó a las absorbancias de las muestras.
Modelos para crecimiento poblacional y cosecha de biomasa. Los estudios
teóricos tienen valor predictivo y se verifican en experimentos de laboratorio y campo.
Los supuestos generales de los modelos determinísticos son los siguientes (Begon y
col. 1996):

37
1. Las poblaciones son aisladas o cerradas (E = I = 0; donde E = tasa de emigración,
I = tasa de inmigración).
2. Las dinámicas son coetáneas (cohortes o individuos de la misma edad).
3. Los individuos son unitarios, excluyendo los modulares (Ej.: colonias de filamentos
o células).
4. Todos los individuos se consideran unidades reproductivas.
5. En modelos discretos, el tiempo entre generaciones Δt se iguala a la unidad.
El modelo de crecimiento densodependiente de partida es el logístico continuo de una
población simple sin estructura:
K
NrN
dt
dN1 (3)
Siendo N(t) la densidad al tiempo t; r la tasa per cápita de crecimiento poblacional
cuando N 0, y K la capacidad de carga o densidad de equilibrio.
La solución de la ecuación (3) es:
rteNKN
KNtN
)()(
00
0 (4)
Siendo N0 = N(0).
El modelo de crecimiento logístico, en su forma discreta, se puede expresar como:
tttt bNrNNN 11 (5)
O bien,
ttt rbNrNN 11 (6)
Donde tN y 1tN son densidades poblacionales a tiempos t y t+1, respectivamente, y
b es una constante definida como un término de retroalimentación densodependiente.
Este modelo es iterativo y el sistema de ecuaciones tiene un arreglo matricial que
representa la estructura de la población en el tiempo (Caswell 2001).

38
Como una aproximación experimental a la tasa de crecimiento instantánea (r), se
determinó la tasa de crecimiento per cápita o de recambio en intervalos de tiempo (t,
t+n) en la fase inicial de crecimiento exponencial, r = ΔN/[N(t)Δt], donde, las tasas
instantánea y media son comparables a densidades bajas (May 1976).
El punto de equilibrio estable (capacidad de carga, KN ˆ ) también se determinó en
forma experimental, a partir del conteo de filamentos en el plateau de la curva de
crecimiento.
El tiempo de duplicación, tg (Begon y col. 1996), es el intervalo temporal en el que la
población inicial ( 0N ) se duplica suponiendo crecimiento exponencial. Esto implica
que N(t) al tiempo tg es dos veces el tamaño inicial de la población (N(t) = 2 0N ). La
definición matemática de tg surge de un despeje e integración de la ecuación
diferencial de crecimiento exponencial:
rNdt
dN
gtN
N
dtrN
dN
0
2 0
0
grtN
N
0
02ln
grt2ln
rt g
2ln (7)
Como se puede observar en la ecuación (7), el tiempo de duplicación es independiente
del tamaño poblacional inicial ( 0N ).
Con el medio escogido como óptimo en el factorial fraccionado 26-1, se reprodujeron
algunos cultivos de A. platensis en botellones de 5 L, y se hicieron conteos de
filamentos clasificados en diferentes tallas (el criterio escogido fue el número de
células por filamento). Las categorías de tallas escogidas fueron: (1) filamentos con
dos células (hormogonios), (2) filamentos con cuatro células y (3) filamentos con más
de cuatro células.
Finalmente, con empleo del mismo medio optimizado, se hizo un análisis de
estabilidad posterior a perturbaciones sobre poblaciones de A. platensis. Dichas
perturbaciones consistieron en extracciones de volúmenes (25, 50 y 75%) en la fase
de saturación o capacidad de carga (K) y dilución con medio optimizado hasta el
enrase del recipiente. Se graficaron las dinámicas de crecimiento hasta el equilibrio.

39
6.2 Ecología de comunidades fitoplanctónicas
Los estudios de comunidades fitoplanctónicas se llevaron a cabo en diversas
localidades del país. Se ubicaron especies nativas de cianobacterias (entre ellas
representantes de los géneros Arthrospira y Spirulina) y eucariotas en ecosistemas
como humedales herbáceos de la península de Paria (Edo. Sucre) y El Clavo
(Barlovento, Edo. Miranda), lagunas de inundación del río Orinoco (Edo. Monagas),
sabanas inundadas todo el año (módulos de Mantecal, Edo. Apure), ambientes
marinos costeros (Bahía de Mochima, Edo. Sucre, y Puerto Cabello, Edo. Carabobo),
lagunas hipersalinas (Las Cumaraguas, Edo. Falcón), algunos sistemas contaminados
con crudos, producto de la explotación petrolera comercial, como la fosa “El Caracol”
(Edo. Anzoátegui), y manaderos naturales de hidrocarburos como la laguna de Boca
Chica en la isla de Margarita, estado Nueva Esparta (Mata García 2003). La figura 6
señala la ubicación geográfica de los sitios visitados.
Figura 6. Ubicación de los sitios de muestreo (el mapa se encuentra en www.guiageo-ameri cas.com/mapas/venezuela.htm).
Salinas de Las Cumaraguas,
Paraguaná Laguna de
Bocachica, Isla de Margarita
Lagunas de inundación, Bajo Orinoco
Humedales de la península de Paria
Humedales de Barlovento
Puerto Cabello
Bahía de Mochima Fosa El Caracol,
Edo. Anzoátegui Módulos de Mantecal

40
Humedales de la península de Paria. La península de Paria está localizada en el
extremo noreste de la región Nororiental de Venezuela, en el estado Sucre, y abarca
un área de 1.078 km² (figura 7a). Limita al norte con el Mar Caribe y al sur con el golfo
de Paria. La Boca Dragón la separa de la Isla de Trinidad en su extremo este.
Geográficamente está localizada entre las coordenadas 10°27’00” N – 10°42’32” N y
62°32’00” W – 63°11’00” W. La geomorfología de la península de Paria es muy
accidentada. Se trata de un sistema montañoso menos macizo que la serranía del
Interior, con relieves poco prominentes, pendientes de 30 a 45% y alturas
relativamente bajas (500 a 1000 m). El sustrato geológico está constituido por rocas
del Cretáceo metamórfico (cuarcitas, esquistos, filitas), y los rasgos más importantes
de la evolución geomorfológica cuaternaria de los depósitos continentales están
influenciados por fenómenos tectónicos. Los suelos son en general poco profundos, de
textura entre franco-arenosa y franco-arcillosa. El clima es tropical húmedo en el norte
caribeño (zona de barlovento) y semi-árido o tropical de sabana en el sur (zona de
sotavento). La precipitación anual media de la península de Paria está comprendida
entre 881,30 y 2348,6 mm, con una clara tendencia de las lluvias a incrementarse en
dirección sur-norte. La temperatura media anual para los primeros 500 m de altura,
gradualmente disminuye de 27,3 a 23,8°C (MARN 1992).
La vegetación natural de la región comprende una variedad importante de formaciones
que notablemente cambian de norte a sur. El norte está dominado por la selva húmeda
tropical y el sur tiene una vegetación de sabana con formas arbóreas características
como el chaparro y el manteco. En las desembocaduras de los ríos que vierten sus
aguas en el Golfo de Paria o el Mar Caribe, existen comunidades de manglares. Hay
diversas zonas húmedas interiores que contienen una vegetación palustrina. El
entorno del área de estudio, es de tipo bosque bajo y denso, con actividad
agropecuaria y levemente intervenido (MARN 1992).
“Palmares III” fue el sitio escogido para el muestreo en los años 2008 y 2009 (Proyecto
Fonacit 2001001850). Es un humedal palustrino, según el sistema de clasificación de
Tabilo-Valdivieso (1999). Sus aguas son someras y semipermanentes y está ubicado
en un terreno bajo o estero. Su localización geográfica precisa es la vertiente sur de la
península, cerca de la población de Los Palmares, con latitud 10°34’58” N, longitud
62°52’10” W y altitud de 30 m.s.n.m. (figura 7b). El área del humedal está cubierta de
vegetación emergente, predominantemente de hábito herbáceo, no siendo evidente la
formación de espejos de agua. Unas pocas especies de fanerógamas forman zonas o
bandas monoespecíficas, aunque también existen algunos sectores con vegetación
mixta, y al sur limita con un sembradío de cocoteros (figura 7c).

41
Figura 7. (a) Mapa de Venezuela donde se destaca a la península de Paria en un recuadro, (b)
ubicación del área de estudio y (c) vista panorámica del humedal “Palmares III” desde una
carretera que lo bordea al norte, la vegetación herbácea cubre la totalidad de su superficie y
forma bandas monoespecíficas, evidenciadas por las tonalidades distintas del color verde, que
cubren toda su superficie sin formar espejos de agua (tomado de Torres y Zoppi de Roa 2010).

42
“Palmares III” tiene un régimen hídrico estacional o semipermanente, producto de una
estacionalidad marcada. Valores medios mensuales que sintetizan 47 años (1953-
2000) de determinaciones del régimen anual de lluvias en las cercanías del humedal,
registradas por la Estación Meteorológica de Irapa, establecen un promedio anual de
precipitaciones de 930,8 mm (MARN 2000). Se define que la estación seca va de
diciembre a marzo, abril es una transición de sequía a lluvia, la estación lluviosa
corresponde al intervalo de mayo a octubre y finalmente noviembre es la transición de
lluvia a sequía (figura 8).
Figura 8. Pluviodiagrama con precipitaciones medias de 47 años (1953-2000) del sur de la península de Paria (datos tomados de la Dirección de Meteorología del MARN 2000).
Las comunidades de plantas que forman zonas monoespecíficas son: (1) gramínea
(Brachiaria mutica (Forssk) Stapt in Pain, Poaceae), (2) leguminosa (Sesbania
exasperata H.B.K., Fabaceae), (3) junco (Cyperus articulatus L., Cyperaceae) y (4)
enea (Typha dominguensis (Pers.) Poir. Ex Steud., Typhaceae).
Humedal de Barlovento. Al este de Caracas, donde finaliza la Cordillera de la Costa,
se inicia la llanura de Barlovento, la cual geopolíticamente es una región del centro de
Venezuela, ubicada en el estado Miranda, que abarca los municipios Acevedo, Andrés
Bello, Brión, Buroz, Páez y Pedro Gual. Esta región se encuentra inserta en la
depresión de Barlovento, siendo una de las regiones naturales que conforman la
denominada región Norte Costera. Se ubica entre los paralelos 10° y 11° latitud norte y

43
los meridianos 65° y 67° longitud oeste. Al norte limita con el mar Caribe; al sur lo hace
con la serranía del Interior y el estado Guárico; al este delimita con la serranía del
Litoral de la cordillera de la Costa, el estado Vargas y otros municipios del estado
Miranda; al oeste culmina en el río Uchire y estado Anzoátegui
(http://es.wikipedia.org/wiki/Barlovento_(Venezuela)).
El área donde se realizó la investigación en 2009 (Ecología de Humedales, materia
electiva del postgrado en Ecología IZET-UCV), se denomina El Clavo, ubicado a 10
msnm, 10°15’34’’ N y 66°7’28’’ O (figura 9). La precipitación total anual es 2484 mm,
con distribución bimodal, con máximos en julio-agosto y noviembre-diciembre y la
mínima entre febrero-marzo, por lo cual la estación seca es corta (MARN 1996). De
acuerdo al sistema de clasificación de Holdridge (Ewel y Madriz 1968, cit. Gordon y
Feo 2007), la zona de vida corresponde a un bosque húmedo tropical. El humedal
herbáceo es inundado por el río Colorado, afluente del río Tuy, y está dominado por
Hymenachne amplexicaulis (Poaceae). En algunos sitios del humedal se encuentran
otras especies como: Montrichardia arborescens (L.) Schott, Mimosa sp., Polygonum
acuminatum Kunth, Ludwigia octovalvis (Jacq.) Raven, Ipomoea sp., Ludwigia
helminthorrhiza (Mart.) Hara, Pistia stratiotes L., Eichhornia crassipes (Mart.) Solms.,
Lemna sp., Salvinia auriculata Aubl., Azolla filiculoides Lam. y Utricularia sp. La textura
de los suelos del humedal es arcillosa (72% arcilla, 22,4% limo y 5,6% arena); el
porcentaje medio de materia orgánica total del suelo es 2,7%, y su pH medio 6,4. La
conductividad del agua varía entre 116 y 294,7 mS/cm, y el pH del agua 6-8 (Feo
2002; cit. Gordon y Feo 2007).
Figura 9. Vista satelital del área de estudio (humedal herbáceo de El Clavo). La línea blanca dibujada en el centro de la imagen señala el transecto levantado en la salida de campo (fuente: http://earth.google.com/).

44
Se muestrearon dos zonas contiguas de vegetación emergente monoespecífica, en un
sentido suroeste–noreste en la parte más estrecha del humedal (ver en la figura 9 la
línea blanca que señala el transecto). Las especies dominantes en cada zona de
vegetación fueron: (1) la hierba arbórea Heliconia marginata (Heliconiaceae), y (2) la
gramínea Hymenachne amplexicaulis. Las figuras 10a y 10b muestran vistas parciales
de las dos zonas de vegetación estudiadas en el humedal herbáceo de El Clavo.
Figura 10. Tomas fotográficas parciales de las dos zonas de vegetación emergente estudiadas: (a) vista de la amplia zona central de Hymenachne amplexicaulis, en primer plano la zona de Heliconia marginata que bordea todo el litoral sur; (b) zona de H. marginata (fotos: Carlos Lugo).
(a)
(b)

45
Lagunas de inundación del Orinoco bajo. El Orinoco es uno de los ríos más
importantes del mundo, no tanto por su longitud y caudal (2.140 km y algo más de
30.000 m³/s), ni por la extensión de su cuenca (989.000 km²); ni siquiera por las
peculiaridades que encierra, sino por su importancia histórica y económica y la
significación que ha tenido para Venezuela, país en el que se extiende la mayor parte
de su cuenca, con casi las dos terceras partes de la misma. Es probablemente el río
más caudaloso del mundo con relación a su cuenca, similar en extensión a la del
Danubio, pero con un caudal que triplica al de este último. En toda la extensión de la
cuenca del Orinoco los climas son isotermos, es decir, climas con escasas variaciones
de temperatura a lo largo del año (la diferencia entre la temperatura media de los
meses más y menos cálidos es de apenas 3°C), como corresponde a la zona
intertropical. Se distinguen de manera bastante nítida cinco grandes tipos de clima en
las zonas bajas (hasta los 800 msnm aproximadamente, según las consideraciones de
Antonio W. Goldbrunner, fundador de los estudios de Meteorología en Venezuela), de
los que destacan el clima de selva (Af en la clasificación de Köppen) y el de sabana
(Aw en la misma clasificación) (http://es.wikipedia.org/wiki/Cuenca_del_Orinoco). El
clima de sabana es el característico de la zona del Orinoco bajo.
La temporada de sequía se extiende de noviembre a abril y la de lluvia se ubica entre
mayo y octubre (Instituto Nacional de Canalizaciones 1999, cit. Rodríguez y
Betancourt 1999). Como sistema de inundación, el río Orinoco presenta en su orilla
bosques sometidos a inundaciones periódicas. Estos bosques crecen cercanos a las
lagunas, y en las áreas de inundación se desarrollan las macrofitas acuáticas (Blanco-
Belmonte 1990). Entre las especies de plantas acuáticas dominantes en estas
lagunas, se encuentran las flotantes libres Eichhornia crassipes (bora) y Paspalum
repens (tapón volador).
La figura 11 muestra el área de estudio a orillas del río Orinoco en la ribera sur del
estado Monagas, en las proximidades del puente Orinokia. Se destacan la ubicación y
una fotografía de la laguna Macapaima, uno de los ambientes estudiados en el periodo
seco del año 2010 (Proyecto PDVSA-IZET Orinokia). Algunos tramos son inundados
en forma directa y constituyen lagunas muy alargadas, paralelas y muy próximas al
curso principal; también se forman brazos y canales que comunican a unas lagunas
con otras. La laguna Macapaima es un sistema que se inunda en forma secundaria por
medio de un canal; estas lagunas suelen tener formas redondeadas y no se ubican
contiguas al río Orinoco, sino que más bien algo apartadas tierra adentro. En época de
sequía, estas lagunas redondas suelen quedar aisladas porque bajan los niveles tanto
del curso principal como de los canales.

46
Figura 11. Ubicación geográfica del área de estudio al sur de Monagas (Orinoco bajo), con detalle de la localización en el mapa y fotografía panorámica de Macapaima, uno de los cuerpos de agua visitados (foto: Rubén Torres).
Módulos inundables de Mantecal. Los módulos de Mantecal son un caso especial de
manejo agroecológico. En ellos, la regulación del caudal de agua se efectúa mediante
la construcción de diques-carreteras en las sabanas inundables de los Llanos bajos de
Venezuela (Cressa y col. 1993). Estos diques tienen una extensión de 3.600 ha con
una capacidad de 40 x 106 m3 (Schargel y González 1973, cit. Cressa y col. 1993).
Los módulos se encuentran en el centro-norte del estado Apure, próximos a Mantecal
y al curso del río Apure. Mantecal es una población satélite de la pequeña ciudad de
Bruzual, capital del Municipio Muñoz, jurisdicción a la cual el pueblo pertenece
(http://es.wikipedia.org/wiki/Mantecal). El estudio en el ecosistema artificial de los
módulos se llevó a cabo en las postrimerías del periodo lluvioso, transición hacia la
sequía, en noviembre de 2010 (Proyecto Mantecal, IZET).
En casi todo el territorio apureño prevalece una vegetación de sabana, con amplio
dominio del componente herbáceo, entre este último se incluyen abundantes
pastizales que alimentan a numerosas reses de ganado vacuno y bufalino de la zona.
También existen matorrales y arbustos, frecuentemente acompañados de enormes
palmas que agrupadas forman los paisajes de morichales y palmares comunes a todo

47
el llano. Posee también secciones intercaladas de selvas, llamadas "de galería", y en
menor grado, zonas de bosque tropical lluvioso y húmedo montano en las
estribaciones de los Andes. En esteros y márgenes de ríos prolifera la vegetación
acuática (http://es.wikipedia.org/wiki/Apure).
Fosas petroleras. Dentro del marco de estudios de ambientes acuáticos
continentales, se determinó la realización de muestreos de plancton en fosas
petroleras, que son ambientes residuales altamente contaminados derivado de su
empleo como sumideros de desechos de la actividad petrolera comercial. Interesan
además estos ambientes contaminados, por el hallazgo de especies fitoplanctónicas
que suponen ser un gran potencial para la biotecnología petrolera, en procura de la
derivación de biocombustibles (biodiesel).
Muestras compuestas de suelo arenoso, materia vegetal, hidrocarburo y agua, fueron
colectadas de la macro-fosa Caracol en la región de Caico Seco, ubicada en el
municipio Aragua del estado Anzoátegui, en el año 2009. Esta fosa fue escogida por
representar un área de exposición histórica a hidrocarburos que operó durante muchos
años como una de las principales y más grandes recolectoras de desechos petrolíferos
de la zona. La figura 12 muestra un acercamiento a la zona de descarga.
Figura 12. Imagen correspondiente a un sector de la orilla de la fosa El Caracol, (Municipio Aragua, Edo. Anzoátegui, 2009), donde se pueden apreciar los desechos petroleros que conforma parte del fondo de la misma (Foto: Olaf Ilzins, IDEA).

48
Igualmente se colectaron muestras de un pozo (P57X) ubicado al norte del estado
Bolívar, 2009. Las muestras provienen en forma específica de una pequeña charca
fangosa en la parte exterior del pozo mencionado, enmarcada en un lugar donde se
encuentra petróleo, ripios y sedimentos de perforación.
Laguna de Boca Chica. Se trata de una laguna costera hipersalina ubicada en la
proximidad de Punta Arenas, en el extremo suroccidental de la península de Macanao,
isla de Margarita, Edo. Nueva Esparta (figura 13). Se trata de un ambiente extremo
donde se han reportado emanaciones naturales de crudos. El sitio fue visitado en julio
de 2009 para obtener algunas muestras fitoplanctónicas.
Figura 13. Ubicación de la laguna de Boca Chica (círculo azul) en la península de Macanao, isla de Margarita, estado Nueva Esparta (mapa: http://www.disfrutevenezuela.com/Municipio-Peninsula-de-Macanao-Mapa.html).
Mata García (2003) cita en forma textual: “En lo que respecta a Margarita, en un
artículo del año 1919, Charles Caracristi afirmó haber descubierto un mene en Punta
Arenas (Península de Macanao). Aguerrevere (1936), expresa en el mismo orden de
ideas: “... en la región occidental de Margarita, cerca de la Laguna de Bocachica, hay
unas pequeñas manifestaciones de petróleo (...), no quiere esto decir que se haya
comprobado definitivamente que no hay depósitos de valor comercial en ese lugar”.
Lorenz (1951, cit. Mata García 2003), ofreció una explicación de las potencialidades
petrolíferas de Margarita: “La isla de Margarita propiamente ofrece pocas condiciones
favorables para la existencia de yacimientos petrolíferos, en vista de que: 1) el

49
cretácico, por su carácter metamórfico, cabe descartarlo como roca madre; 2) los
sedimentos terciarios no poseen espesores halagadores; 3) estructuras favorables son
inestables o están parcialmente erosionadas”.
Bahía de Mochima. Esta bahía profunda y semi-cerrada, de gran belleza paisajística,
forma parte del Parque Nacional Mochima, Edo. Sucre, en los límites con el Edo.
Anzoátegui, a orillas del mar Caribe. El Parque Nacional Mochima se encuentra
altitudinalmente ubicado desde 0 hasta 600 msnm, al noreste de Venezuela, entre las
ciudades de Barcelona, Puerto la Cruz y Cumaná, y se extiende a lo largo de la costa
en un área de 94.935 ha (http://www.mochima.org/).
El clima del Parque Nacional Mochima es semiárido. En la costa las temperaturas
oscilan entre 22 y 28°C, y existen dos periodos contrastantes: (1) una época seca que
se extiende de enero a mayo, con una media de lluvias de 3,5 mm/mes y (2) una
época lluviosa entre junio y diciembre, con una media de lluvias de 60-70 mm/mes. En
el parque existe una variada vegetación en donde predominan cactus, arbustos,
helechos y orquídeas, encontrándose árboles de mayor tamaño en la zona más alta
del parque. Parte de la flora la conforman los mangles y la hierba de vidrio (Salicornia
fruticosa, Sesuvium portulacastrum y Batis marítima). Se pueden conseguir cactáceas
como el guamacho y leguminosas como el cují y el dividive (http://www.mochima.org/).
En la población de Mochima, ubicada al sur de la bahía, se encuentra la Estación
Biológica Mochima (Sucre), que es una dependencia de la Fundación Instituto de
Estudios Avanzados (IDEA). El proyecto de microalgas (Proyecto BID-Fonacit
2006000537), cuya acometida impulsó la Asociación Civil Gente de Ciencias, está
embebido en un megaproyecto (Hidrocarburos Verdes) de esta institución
gubernamental. Por tal motivo, se procedió a la toma de muestras fitoplanctónicas y
zooplanctónicas en esa localidad marina costera, en agosto de 2009, con el interés
especial de colectar especies de microalgas marinas de gran potencial para la
obtención de biocombustibles.
Salinas de Las Cumaraguas. Las Cumaraguas es un sitio natural de salinas,
ubicadas al noreste de la península de Paraguaná, específicamente en el Municipio
Falcón. Su acceso es por vía terrestre. Presentan un espectáculo digno de ver en
horas del atardecer, cuando el tanino que contienen las aguas que irrigan ese sector,
les torna el color a rojizo. El estado Falcón cuenta con grandes potencialidades para el
desarrollo de la industria salinera. Existen cinco (05) salinas naturales y unas 220.000
ha de superficie aptas para la construcción de salinas artificiales repartidas a lo largo
del territorio falconiano. De las salinas del estado, sólo Las Cumaraguas se encuentra

50
bajo explotación industrial y privada, siendo el resto de las salinas explotadas de
manera artesanal (http://www.tuplaya.com/paginasfalcon/salinas/lascumaraguas.htm).
Las coordenadas astronómicas de las salinas de Las Cumaraguas son 12°06’ N y
69°54’ W (Guevara y col. 2005).
Ambientes hipersalinos como Las Cumaraguas y Boca Chica, ambos con condiciones
semi-áridas y precipitaciones escasas, son propicios para el desarrollo de especies
extremófilas que toleran grandes concentraciones de NaCl, como especies del género
Dunaliella (Chlorophyta), las cuales constituyen las fuentes vivientes más ricas en
carotenoides, contando hasta 14% de su masa seca (Guevara y col. 2005), lo que se
traduce en grandes posibilidades para su empleo en biotecnología alimentaria,
farmacéutica y pigmentos. Es por esto que en octubre de 2010, en el marco del
proyecto de microalgas para la exploración de especies nativas de gran valor
biotecnológico, se visitaron las salinas de Las Cumaraguas para obtener muestras de
agua con grandes densidades poblacionales de Dunaliella spp., para apuntar al inicio
de su cultivo en condiciones controladas de laboratorio y optimizar el medio nutritivo.
Puerto Cabello. El muelle de Puerto Cabello está localizado en la ciudad del mismo
nombre, norte del estado Carabobo, región Central de Venezuela, a orillas del mar
Caribe. Se trata de un ambiente marino costero muy perturbado por actividades
antrópicas relacionadas con dinámicas de las faenas portuarias, refineras y el vertido
de efluentes domésticos. La ciudad de Puerto Cabello tiene una población aproximada
de 203.633 habitantes y su puerto es el segundo en importancia para el país, debido a
su gran actividad de importación de materias primas para el sector industrial
venezolano, que normalmente se trasladan hacia Valencia, la capital del estado
Carabobo, y otras regiones del país. El Palito está a la entrada de Puerto Cabello, una
de las refinerías de petróleo más importantes del país. La ciudad también posee
relevancia en el área comercial y turística, pero su actividad por tradición ha sido la
portuaria (http://www.ipapc.gov.ve/).
La temperatura oscila entre 23 y 30°C, con una media anual de 27°C. El clima es
semiárido en la franja litoral con una precipitación anual de 463 mm. En tierra
predomina la vegetación xerófila, y en ambientes estuarinos y marinos costeros se
extiende el bosque de manglar, con el mangle rojo (Rhizophora mangle) como la
especie dominante (http://es.wikipedia.org/wiki/Puerto_Cabello). Se muestrearon
diversos punto en el muelle, en enero de 2011, en procura de evaluar la composición
fitoplanctónica y zooplanctónica del lugar y caracterizar su estado ambiental.

51
El trabajo de campo se ejecutó como se describe a continuación:
Se hicieron muestreos en ambientes parcelados, como humedales herbáceos y
litorales con vegetación, por el modelo de muestreo estratificado sin afijación (número
homogéneo de muestras), que sirvió de muestreo piloto para trabajos posteriores en
los mismos sitios con muestreo por conglomerados con afijación óptima, es decir,
mayor número de muestras se tomaron en lugares con varianzas altas y menor
número en zonas con varianzas bajas (Azorín 1970, Scheaffer y col. 1987). En cada
estrato se tomaron al azar cinco muestras de fitoplancton, para ello se levantaron
parcelas de 2 × 2 m y se escogieron pares ordenados con una tabla de números
aleatorios. Las muestras se colectaron con una botella de captación LaMotte de 1 L,
en forma sistemática desde el borde hasta el centro de la parcela, de este modo sólo
se perturbaron los puntos muestreados, sin alterar el resto. Las muestras de
fitoplancton se fijaron in situ con solución de lugol.
En aguas abiertas (espejos de agua y ambientes marinos costeros), el fitoplancton se
colectó en forma diferente. Tres puntos a lo largo de un transecto, con aleatorización
del tiempo en minutos para la escogencia del primer punto, permitieron tomar dos
muestras puntuales de fitoplancton en cada sitio con una botella LaMotte de 1 L.
Posteriormente, en forma continua se tomaron muestras muy grandes por medio de
dos arrastres horizontales de 5 min con red (øporo = 105 μm) realizados desde una
pequeña embarcación con motor fuera de borda a una velocidad de 2 nudos2. El
sistema de captura por botella permitió la retención de fitoplancton pequeño
(nanoplancton), pues constituye un tipo de trampa hermética que captura la muestra
con el agua, mientras que la red tomó en forma selectiva microalgas de mayor tamaño.
Como complemento al estudio comunitario, en los mismos ambientes se tomaron
algunas muestras de zooplancton, para conocer la microbiota animal asociada al
fitoplancton. Se tomaron dos muestras con botella LaMotte de 1 L en humedales
herbáceos y ambientes litorales, y dos barridos con red (øporo = 105 μm) en ambientes
abiertos (lagunas de inundación y ambientes marinos costeros). Las muestras se
fijaron in situ con solución de formalina al 10% V/V.
Finalmente, se midieron in situ algunas variables ambientales (pH, conductividad,
salinidad, O2 disuelto y temperatura) con un medidor digital HORIBA U-10, bajo la
2 1 nudo = 1 milla náutica por hora = 0,5144 metros por segundo (SI). Esta definición se basa en el acuerdo internacional sobre la longitud de la milla náutica, adoptado por EE. UU. (que utilizaba previamente una longitud de 1.852,249 m) y el Reino Unido (que utilizaba previamente una longitud de 1.853,184 m), entre otros países (http://es.wikipedia.org/wiki/Nudo_(unidad)).

52
lámina de agua. La radiación solar se determinó como luminancia (densidad de
fotones) con un luxímetro LUTRON-LX y la profundidad con una vara graduada.
Laboratorio. Una vez trasladadas las muestras, éstas se limpiaron y procesaron para
identificar las especies y determinar sus abundancias (conteos en cámara Bogorov
bajo estereoscopio o en cámara Utermöhl bajo microscopio invertido). Las especies
fitoplanctónicas se identificaron con ayuda de claves e ilustraciones: Ortega (1984),
Varela y col. (1986) y Matto y Parra (1995). Para el zooplancton la taxonomía se hizo a
partir de los trabajos de Edmonson (1959), Elster y Ohle (1978), Infante (1980),
Dussart (1984), Reid (1984), Zoppi de Roa y Vásquez (1991), Zoppi de Roa (1993),
Velho y Lansac-Tôha (1996) y Velho y col. (1996).
Para algunas muestras de agua tomadas en un humedal de Barlovento, se hicieron
algunos análisis de nutrientes, principalmente nitrógeno (método de Kjeldahl (1883)) y
fósforo (método de Murphy-Riley (1962)). Las muestras de agua se fijaron en campo
con ácido sulfúrico. De seis puntos muestreados en la zona de H. marginata, las
muestras tomadas en los puntos 1, 2 y 3 estaban muy próximas, por lo que fueron
unidas y se obtuvo una muestra combinada, lo que permitió tener una muestra más
representativa. Los métodos químicos están representados en las figuras 14 y 15.
Para muestras de Paria, algunos nutrientes se determinaron por absorción atómica.
Figura 14. Marcha analítica simplificada para la determinación de nitrógeno en muestras de agua con el método Kjeldahl (1883) (continúa en la página siguiente).
Filtración de la muestra
Muestra de campo fijada con ácido sulfúrico
Destrucción de la materia orgánica con ácido sulfúrico concentrado
Equipo de destilación Kjeldahl

53
Figura 14. Continuación (Fotos de equipo Kjeldahl y sucedáneas tomadas de una presentación digital del curso de Ecología de Humedales, Postgrado en Ecología, IZET, 2009).
Figura 15. Marcha analítica simplificada para la determinación del fósforo por el método colorimétrico de Murphy – Riley (1962). Las dos imágenes inferiores fueron tomadas de una presentación digital del curso de Ecología de Humedales, Postgrado en Ecología, IZET, 2009.
Destrucción de la materia orgánica con ácido sulfúrico concentrado
Muestra de campo fijada con ácido sulfúrico
Filtración de la muestra
Medición y curva de calibración

54
Índices de diversidad. Para el análisis comunitario se cumplieron las pautas
siguientes:
1. Los cómputos del fitoplancton quedaron expresados según el nivel de organización
o forma de vida en células, colonias y filamentos por litro, para el caso de muestras
obtenidas con botella de captación, o en células, colonias y filamentos por metro
cúbico para el caso de muestras tomadas con red. En forma análoga para el
zooplancton, los organismos totales quedaron expresados en individuos por litro
(Ind./L) o individuos por metro cúbico (Ind./m3), según que el método de captura
haya sido por empleo de botella de captación o red, respectivamente. Para la
extrapolación al volumen de arrastre o barrido con red, V, se calculó la distancia, d,
recorrida en bote a partir del tiempo recorrido y la velocidad de la embarcación, la
cual fue aproximadamente constante, resultando un movimiento rectilíneo
uniforme. Esta distancia se multiplicó por el área de la boca de la red, A, que es
circular (A = πr2), obteniéndose el volumen, V, de un cilindro, V = dπr
2.
2. La riqueza (S) y la equidad ( 'J ) (Pielou 1975) son dos parámetros comunitarios de
relevancia para caracterizar la diversidad, por lo que fueron calculados. La riqueza
es la sumatoria de las especies presentes en la comunidad y es el índice de
diversidad más sencillo que se conoce. Por su parte, la ecuación que describe a la
equidad es la siguiente:
SHHHJ /ln'=/'=' max (8)
Donde 'H es el índice de diversidad de Shannon-Wiener (Shannon y Weaver 1949) y
SH ln=max es la diversidad máxima. El uso del índice de Shannon-Wiener se basa en
suponer que los individuos se muestrean al azar en una población infinita y que todas
las especies están representadas en la muestra. Este índice se define
matemáticamente como:
i
S
i
i ppH ln-='1=
∑ (9)
Siendo pi la fracción de abundancia de la especie i.
3. Se empleó el índice de Simpson, D (Simpson 1949), el cual es un índice de
dominancia y representa la probabilidad de que dos individuos obtenidos al azar de

55
una muestra pertenezcan a la misma especie. Por lo tanto, valores mayores
indican menor diversidad. Su representación matemática es:
S
i
ii
NN
nnD
1= 1
1= (10)
El cual normalmente es calculado como:
S
i
ipD1=
2= (11)
Cuando se utiliza el recíproco de este índice (1/D), los valores son interpretados como
el número de especies esperado de una muestra con una determinada distribución de
individuos en especies. El valor 1/D aumenta cuando la muestra es más equitativa, por
lo cual 1/D tiene un significado biológico más claro que el índice original.
Otro índice de dominancia empleado es el de Berger-Parker (1970):
N
Nd máx= (12)
Donde:
máxN = número total de individuos de la especie más abundante
N = número total de individuos en la comunidad
Se determinó el índice de similitud o distancia Jaccard (1901), el cual es definido como
la razón entre el tamaño de la intersección y el tamaño de la unión de dos conjuntos
de muestras, por lo que Del Pino y col. (2006) lo clasifican como un índice de
diversidad beta (β), donde se considera la tasa o grado de cambio en la composición
de especies entre diferentes comunidades en un paisaje.
El programa ejecutable libre PAST se utilizó para la determinación de los índices
anteriormente descritos.
Análisis de Datos. El manejo de la información obtenida se hizo con un Análisis de
Datos (Andrienko y Andrienko 2006), que describe el patrón de distribución espacial y
temporal de las especies asociadas a diferentes ambientes. El análisis de datos se
llevó a cabo en tres fases:
1. Descriptiva (análisis de homogeneidad, asimetría y curtosis de las variables,
evaluación de valores extremos). Se establecieron estadísticos robustos para

56
variables con notables asimetrías y valores extremos. Se eliminaron del análisis
especies zooplanctónicas con desviaciones estándares menores a 0,4 Ind./L (Peña
y Rodríguez 2003).
2. Análisis de Componentes Principales (ACP) iterativo para escoger las variables
estadísticamente importantes representadas en gráficos biplot tridimensionales
(Fluir 1988). En este análisis se utilizaron las densidades poblacionales de las
especies fitoplanctónicas (células, colonias y filamentos por litro) y zooplanctónicas
(Ind./L) como variables dependientes aleatorias. Con esta metodología se
seleccionaron las especies que se consideraron más importantes desde el punto
de vista estadístico y que posteriormente se emplearon en el Análisis de
Agrupamiento.
3. Análisis de Agrupamiento o “Cluster Analysis” (Anderberg 1973), con empleo de la
Distancia Euclídea como medida de disimilitud, el método de Ward’s como método
de agrupamiento y las variables dependientes aleatorias escogidas con el ACP:
densidades poblacionales de las especies zooplanctónicas (Ind./L). Se calcularon
los centroides para establecer las especies con mayor ponderación. Un Análisis
Multivariado de Varianza (acrónimo en inglés MANOVA) se utilizó para establecer
diferencias significativas entre grupos (α = 0,05).
Los análisis descriptivos multivariados se realizaron en el software MVSP 3.0. Los
programas JMP versión 3.2.1 (1997) y SPAD versión 3.0 (1999) sirvieron para hacer
Análisis de Datos en general, y MINITAB release 13.20 (2000) fue otra herramienta útil
para evaluar la fase descriptiva.
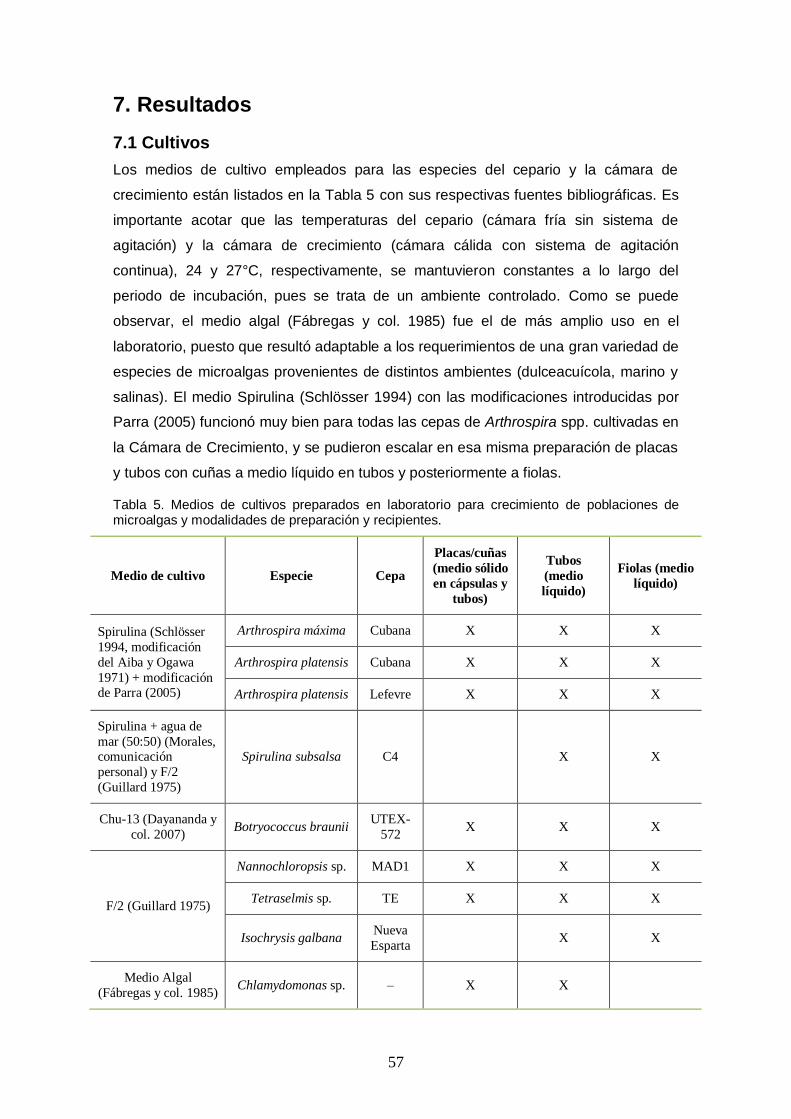
57
7. Resultados
7.1 Cultivos
Los medios de cultivo empleados para las especies del cepario y la cámara de
crecimiento están listados en la Tabla 5 con sus respectivas fuentes bibliográficas. Es
importante acotar que las temperaturas del cepario (cámara fría sin sistema de
agitación) y la cámara de crecimiento (cámara cálida con sistema de agitación
continua), 24 y 27°C, respectivamente, se mantuvieron constantes a lo largo del
periodo de incubación, pues se trata de un ambiente controlado. Como se puede
observar, el medio algal (Fábregas y col. 1985) fue el de más amplio uso en el
laboratorio, puesto que resultó adaptable a los requerimientos de una gran variedad de
especies de microalgas provenientes de distintos ambientes (dulceacuícola, marino y
salinas). El medio Spirulina (Schlösser 1994) con las modificaciones introducidas por
Parra (2005) funcionó muy bien para todas las cepas de Arthrospira spp. cultivadas en
la Cámara de Crecimiento, y se pudieron escalar en esa misma preparación de placas
y tubos con cuñas a medio líquido en tubos y posteriormente a fiolas.
Tabla 5. Medios de cultivos preparados en laboratorio para crecimiento de poblaciones de microalgas y modalidades de preparación y recipientes.
Medio de cultivo Especie Cepa
Placas/cuñas
(medio sólido
en cápsulas y
tubos)
Tubos
(medio
líquido)
Fiolas (medio
líquido)
Spirulina (Schlösser
1994, modificación
del Aiba y Ogawa
1971) + modificación de Parra (2005)
Arthrospira máxima Cubana X X X
Arthrospira platensis Cubana X X X
Arthrospira platensis Lefevre X X X
Spirulina + agua de
mar (50:50) (Morales, comunicación
personal) y F/2
(Guillard 1975)
Spirulina subsalsa C4 X X
Chu-13 (Dayananda y
col. 2007) Botryococcus braunii
UTEX-
572 X X X
F/2 (Guillard 1975)
Nannochloropsis sp. MAD1 X X X
Tetraselmis sp. TE X X X
Isochrysis galbana Nueva
Esparta X X
Medio Algal
(Fábregas y col. 1985) Chlamydomonas sp. – X X
58
Tabla 5. Continuación.
Medio de cultivo Especie Cepa
Placas/cuñas
(medio sólido
en cápsulas y
tubos)
Tubos
(medio
líquido)
Fiolas (medio
líquido)
Medio Algal
(Fábregas y col. 1985)
Chlorella vulgaris – X X
Scenedesmus sp. – X X X
Medio Algal con
adición de silicatos Chaetoceros sp. – X
Medio Algal con
adición de agua de
mar
Porphyridium
cruentum C1 X X
Rhodosorus marinus C2 X X
Tetraselmis sp. TE X X
Medio Algal con
adición de solución
saturada de NaCl
(grado alimenticio)
Dunaliella salina – X X
Dunaliella viridis – X
Dunaliella
primolecta DPN X
Serpa y Calderón
(2006)
Dunaliella salina – X X
Dunaliella viridis – X
Dunaliella
primolecta DPN X
Bristol (Bold 1949) Haematococcus
pluvialis HPOC8 X
Una evaluación microscópica de las diferentes cepas, cultivadas en la cámara fría sin
agitación y la cámara cálida o de crecimiento rápido con agitación, pudo comprobar
que los cultivos estaban en buen estado (monoalgales y poco detrito), donde las cepas
cubanas de A. platensis y A. maxima mostraron filamentos helicoidales, si bien en
poblaciones de la segunda se evidenciaron algunos filamentos lineales. Por su parte,
la cepa Lefevre de A. platensis se caracterizó por mostrar una población integrada en
forma absoluta por filamentos lineales en todas las “réplicas” ubicadas en la Cámara
de Crecimiento. La figura 16 muestra un conjunto de fotografías tomadas con una
cámara digital PAX-CAM acoplada al microscopio invertido y al computador, lo que
permitió visualizar los filamentos con alta resolución y detectar el estado de pureza de
los cultivos. Durante todo el periodo de mantenimiento, las observaciones al
microscopio no detectaron invasiones de otras especies de microalgas.

59
Figura 16. Algunas imágenes tomadas bajo microscopio invertido de las poblaciones de las diferentes cepas de Arthrospira spp. en la Cámara de Crecimiento (LOA-IZET).

60
Fig. 16. Continuación.
La cianobacteria Spirulina subsalsa se cultivó en medio Spirulina + agua de mar
(50:50), puesto que la cepa existente en la colección proviene de un ecosistema
estuarino. Los cultivos fueron viables y las poblaciones crecieron de modo rápido. Sin
embargo, una observación que se hizo recurrente en estos cultivos, es que una vez
que las poblaciones eran grandes, se iniciaba la formación de grandes colonias de
filamentos muy adosados entre sí (grandes flóculos), y que además segregaban una
sustancia mucilaginosa abundante y viscosa que envolvía a las colonias y, al cabo de
pocos días, dicha sustancia mezclada con sal precipitada del medio se endurecía y
conformaba una especie de cascarón. La consecuencia de esto es que las colonias de
filamentos se fijaban a las paredes del recipiente, y no podían ser desprendidas, aún
con la agitación constante del aire presurizado enviado del blower. Como fórmula
alternativa, S. subsalsa fue cultivada en medio F/2. Esta modificación de medio derivó
en un resultado positivo consistente en la no formación de las capas cementantes
evidenciadas en el medio Spirulina + agua de mar (50:50), pero en contraposición a
ello las poblaciones no incrementaron su tamaño en la forma que lo hacían en el
medio anterior, las mismas se mantuvieron en flóculos pequeños y dispersos con una
coloración verde menos intensa que el caso citado.

61
Otras especies de la colección. En términos generales, los medios funcionaron,
salvo el medio Algal para Dunaliella primolecta y el medio Serpa y Calderón (2006)
para todas las especies del género Dunaliella, puesto que las cepas no pudieron
adecuarse a los mismos bajo las condiciones de la Cámara de Crecimiento. Algunas
de las fiolas con cultivos de D. salina y D. viridis en medio Algal exhibieron
crecimientos poblacionales notables, dada la coloración verde que adquirieron y no
incolora como en D. primolecta. Algunas de las cepas que han crecido exitosamente
en fiolas de 250 mL se trasladaron a fiolas de 2000 mL, y han podido replicarse. No
obstante, dos cultivos en fiolas de 250 mL de D. viridis en buen estado de crecimiento
al ser unidas y llevadas a una fiola de 2000 mL con medio nuevo no pudieron
adecuarse y permanecieron en estado de latencia.
Una especie que ha crecido muy bien en fiolas pequeñas y se ha trasvasado a fiolas
grandes es la clorofita dulceacuícola Haematococcus pluvialis. Esta microalga ha
permanecido en una fase de crecimiento rápido con poblaciones sanas y libres de
contaminación por otras especies de microalgas. El color que siempre se ha
observado en los cultivos de esta especie es un verde oliva, el cual es típico de una
población con una estructura de células jóvenes, y no la coloración roja púrpura
predominante en una población de células maduras en condiciones de estrés.
Los cultivos de las especies Botryococcus braunii, Nannochloropsis sp. y Tetraselmis
sp., especies de particular interés en la producción de biocombustibles, se
mantuvieron en muy buen estado. La primera especie creció bien en medio Chu-13
(Dayananda y col. 2007) y las otras dos hicieron lo propio en el medio F/2 (Guillard
1975). Las tres especies pudieron ser escaladas a fiolas de 2000 mL en la cámara
cálida, y el crecimiento poblacional fue óptimo. Como observación curiosa, uno de los
cultivos de B. braunii en fiolas sedimentó, las células formaron grandes colonias y al
parecer produjeron una sustancia cementante que las adhirió al fondo y paredes del
recipiente de vidrio, a pesar de que el sistema de la cámara cálida contó con agitación
fuerte por medio de aireación presurizada. La figura 17 muestra una serie de
fotografías donde se evidencia la condición monoalgal de los cultivos de estas
especies, y sólo algunos animales (rotíferos) y protozoarios (ciliados) han sido
observados.
Finalmente, como se presentó en la Tabla 5, algunas especies cultivadas, como
Chlamydomonas sp., Chlorella vulgaris y Chaetoceros sp., no pudieron ser escaladas
a fiolas, bien sea porque sus tamaños poblacionales eran pequeños y permanecieron
en tubos de ensayo dentro de la cámara fría, o algunas fueron trasvasadas y no
pudieron crecer a mayores volúmenes (efecto de dilución).
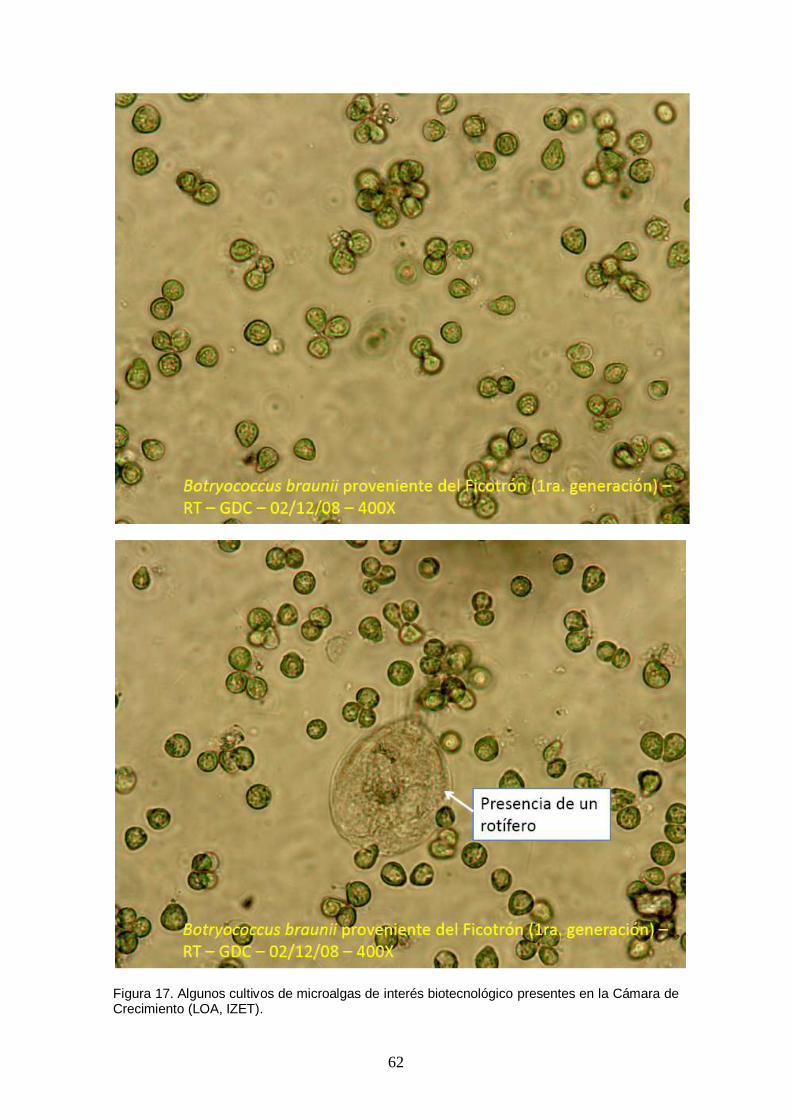
62
Figura 17. Algunos cultivos de microalgas de interés biotecnológico presentes en la Cámara de Crecimiento (LOA, IZET).

63
Figura 17. Continuación.

64
Figura 17. Continuación.
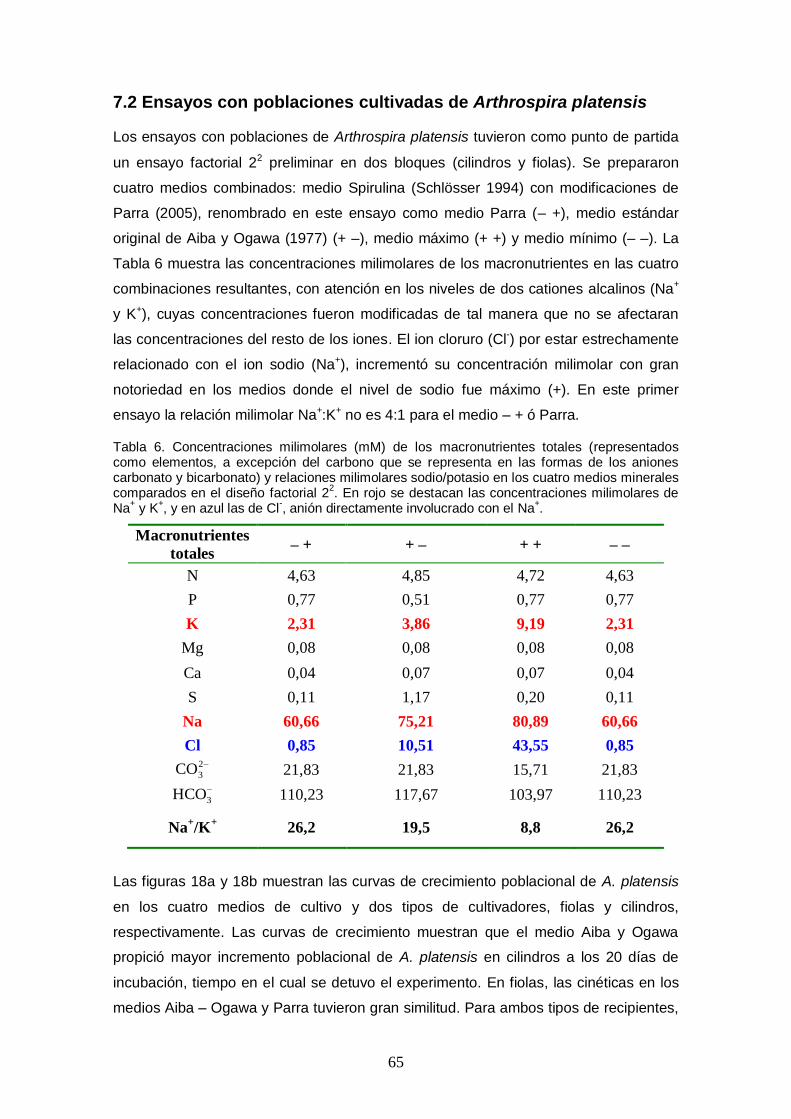
65
7.2 Ensayos con poblaciones cultivadas de Arthrospira platensis
Los ensayos con poblaciones de Arthrospira platensis tuvieron como punto de partida
un ensayo factorial 22 preliminar en dos bloques (cilindros y fiolas). Se prepararon
cuatro medios combinados: medio Spirulina (Schlösser 1994) con modificaciones de
Parra (2005), renombrado en este ensayo como medio Parra (– +), medio estándar
original de Aiba y Ogawa (1977) (+ –), medio máximo (+ +) y medio mínimo (– –). La
Tabla 6 muestra las concentraciones milimolares de los macronutrientes en las cuatro
combinaciones resultantes, con atención en los niveles de dos cationes alcalinos (Na+
y K+), cuyas concentraciones fueron modificadas de tal manera que no se afectaran
las concentraciones del resto de los iones. El ion cloruro (Cl-) por estar estrechamente
relacionado con el ion sodio (Na+), incrementó su concentración milimolar con gran
notoriedad en los medios donde el nivel de sodio fue máximo (+). En este primer
ensayo la relación milimolar Na+:K+ no es 4:1 para el medio – + ó Parra.
Tabla 6. Concentraciones milimolares (mM) de los macronutrientes totales (representados como elementos, a excepción del carbono que se representa en las formas de los aniones carbonato y bicarbonato) y relaciones milimolares sodio/potasio en los cuatro medios minerales comparados en el diseño factorial 2
2. En rojo se destacan las concentraciones milimolares de
Na+ y K
+, y en azul las de Cl
-, anión directamente involucrado con el Na
+.
Macronutrientes
totales – + + – + + – –
N 4,63 4,85 4,72 4,63
P 0,77 0,51 0,77 0,77
K 2,31 3,86 9,19 2,31
Mg 0,08 0,08 0,08 0,08
Ca 0,04 0,07 0,07 0,04
S 0,11 1,17 0,20 0,11
Na 60,66 75,21 80,89 60,66
Cl 0,85 10,51 43,55 0,85 2
3CO 21,83 21,83 15,71 21,83
3HCO 110,23 117,67 103,97 110,23
Na+/K
+ 26,2 19,5 8,8 26,2
Las figuras 18a y 18b muestran las curvas de crecimiento poblacional de A. platensis
en los cuatro medios de cultivo y dos tipos de cultivadores, fiolas y cilindros,
respectivamente. Las curvas de crecimiento muestran que el medio Aiba y Ogawa
propició mayor incremento poblacional de A. platensis en cilindros a los 20 días de
incubación, tiempo en el cual se detuvo el experimento. En fiolas, las cinéticas en los
medios Aiba – Ogawa y Parra tuvieron gran similitud. Para ambos tipos de recipientes,

66
el medio + + sucedió a los anteriores. Puede observarse que el medio – – se
caracterizó por presentar un período de latencia y adecuación muy prolongado, hasta
que la población empezó a crecer abruptamente a partir del día 15. No obstante, entre
este tratamiento y los demás no hubo diferencias significativas (α = 0,05).
Figura 18. Curvas de crecimiento poblacional de Arthrospira platensis en (a) fiolas y (b) cilindros. La densidad poblacional está expresada en términos de absorbancia a 680 nm.
La figura 19 presenta una comparación gráfica de la producción de biomasa seca en
los cuatro medios propuestos para el ensayo factorial. Como se puede evidenciar el
medio Aiba y Ogawa una vez más resultó ser el más productivo en cilindros y fiolas. El
medio Parra tuvo en fiolas una producción de biomasa que casi iguala a la del Aiba y
Ogawa, pero comparativamente baja en cilindros. El medio + + tuvo una producción
similar en ambos tipos de recipientes, y la cosecha del medio – – tuvo una producción
muy baja en los dos envases, siendo ligeramente superior en cilindro. En este ensayo
se apreció que hubo diferencias significativas entre fiolas y cilindros para los
tratamientos – – y Parra, así como entre el medio – – y los medios restantes para
ambos recipientes. La dispersión de datos (desviaciones estándares) fue pequeña.
0
0,5
1
1,5
2
2,5
3
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
T iempo (días )
A6
80
Aiba y Ogawa Máximo (+ +)Mínimo (– –) Parra
0,000
0,500
1,000
1,500
2,000
2,500
1 2 3 4 5 6 7 8 9 1011121314151617181920
A6
80
(a)
(b)

67
Figura 19. Producción de biomasa seca en fiolas y cilindros de 2 L para cada medio mineral. La biomasa está expresada en gramos por dos litros. Intervalos de confianza: media desviación estándar.
Los valores de pH de los cuatro medios ensayados caracterizaron el ambiente químico
donde crecieron los filamentos de A. platensis durante el periodo de incubación. Los
medios Aiba y Ogawa, Parra y + + exhibieron un gran paralelismo a lo largo de los
días del ensayo con una tendencia hacia el incremento. En contraste, el medio – –
mostró una tendencia inversa, al iniciar con un pH más alto que los otros tres medios y
después decrecer, hasta que en los últimos 5 días experimentó un nuevo aumento, el
cual coincidió con el comienzo del crecimiento rápido de A. platensis en ese medio,
luego de una latencia y adecuación prolongadas. Durante el lapso en el que el pH bajó
para el medio mínimo, aparentemente desfavorable para A. platensis, prosperó otra
especie de cianobacteria oportunista, Microcystis aeruginosa, la cual desapareció del
medio una vez el pH volvió a subir y A. platensis volvió a dominar (figura 20).
Figura 20. Valores comparativos de pH mostrados por los diferentes medios de cultivo. En la figura se señala con un óvalo rojo el lapso de disminución de pH del medio – – y la contaminación del mismo por la cianobacteria Microcystis aeruginosa.
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
2
Mínimo (– –) Máximo (+ +) Parra Aiba y Ogawa
Bio
ma
sa (
g/2
L)
Fiola Cilindro
8,60
8,80
9,00
9,20
9,40
9,60
9,80
10,00
10,20
10,40
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Tiempo (días)
pH
Mínimo (– –) Máximo (+ +) Parra Aiba y Ogawa
Contaminación del medio por Microcystis aeruginosa

68
La conductividad eléctrica se mantuvo constante en todos los medios a lo largo del
periodo de incubación, y resultó similar para los medios Parra y Aiba - Ogawa,
mientras que para el medio máximo (+ +), los valores de conductividad resultaron
ligeramente mayores por su condición de medio enriquecido. Por su parte, el medio
mínimo (– –) tuvo valores muy inferiores y paralelos a los registrados para los medios
anteriores (figura 21).
Figura 21. Valores comparativos de conductividad (S/cm) mostrados por los diferentes medios de cultivo.
En la figura 22 se observa la relación entre la absorbancia de la masa celular y el peso
seco de la misma. La curva muestra una tendencia lineal que fue evaluada con una
regresión lineal simple, con un coeficiente de regresión R2 = 0,9935, que evidencia una
correlación fuerte. Se calculó el peso seco de la masa celular presente en 1000 ml de
cultivo, en base a la linealidad de la fase de crecimiento rápido, con la obtención del
valor medio de 586 mg (0,586 g/L) de masa seca de A. platensis en ese volumen.
Figura 22. Curva de calibración A680 vs. Peso seco (mg). Las variables tienen una relación lineal (R
2 = 0,9935).
y = 0,0011x + 0,0578 R² = 0,9935
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
0 200 400 600 800 1000 1200
Ab
so
rban
cia
(λ =
680 n
m)
Peso seco (mg)
0,00
5,00
10,00
15,00
20,00
25,00
30,00
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Tiempo (días)
Co
nd
. (S
/cm
)
Mínimo (– –) Máximo (+ +) Parra Aiba y Ogawa

69
El experimento factorial 22 se replicó en un nuevo conjunto de fiolas, donde hubo un
reajuste de las concentraciones de los medios nutritivos del ensayo anterior (Tabla 7).
En este nuevo ensayo, la nomenclatura de los medios varió, el medio 1 corresponde al
anteriormente mencionado medio Parra, el medio 2 es el Aiba y Ogawa, el medio 3
corresponde al medio máximo (+ +) y por último el medio 4 es el mínimo (– –). Se
logró un mejor arreglo de las combinaciones milimolares de los macronutrientes, para
obtener en el medio 1, con la modificación de Parra (2005), una relación Na+:K+ bien
aproximada a la relación fisiológica normal de estos dos cationes en células vivas, lo
que no se consiguió en el primer perfil nutricional.
Tabla 7. Concentraciones milimolares totales de los macronutrientes que integran cada uno de los medios de cultivo preparados. En rojo se destaca la relación 4:1 de K y Na en el medio 1.
Nutriente Medio 1 Medio 2 Medio 3 Medio 4
N 29,44 29,40 29,86 29,90
P 2,87 2,87 3,33 3,30
K+ 63,75 17,22 18,07 17,22
Mg2+
0,81 0,81 0,81 0,81
Ca2+
0,27 0,27 0,27 0,26
SO42-
6,60 6,60 1,34 0,86
Na+ 238,07 284,60 282,62 238,06
Cl- 17,65 17,65 75,83 1,40
CO32-
38,70 38,70 27,85 38,70
HCO3- 162,02 162,02 143,16 163,02
Na+/K
+ 3,73 4 16,5 15,6 13,8
La figura 23 muestra las curvas de crecimiento poblacional de A. platensis en los
cuatro medios de cultivo. Como en el ensayo anterior, los medios 1 y 2 presentaron
cinéticas similares, aunque la pendiente del segundo supera un poco a la del primero.
La curva del medio 3 tiene la tercera pendiente más alta, mientras que en la curva de
crecimiento correspondiente al medio 4 se aprecia que la fase de latencia es muy
prolongada y la población rápidamente crece los últimos 5 días del ensayo. La fase de

70
0,000
0,500
1,000
1,500
2,000
2,500
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
Tiempo (días)
A6
80
Medio 1 Medio 2 Medio 3 Medio 4
crecimiento rápido de todas las curvas presentó una fuerte tendencia lineal. En
ninguno de los casos se alcanzó el plateau o capacidad de carga.
Figura 23. Crecimiento poblacional de Arthrospira platensis en cuatro medios de cultivo. En la fase de crecimiento rápido A680 y el tiempo presentaron correlaciones lineales fuertes, como se evidencia en los valores del coeficiente de determinación (R
2).
La figura 24 exhibe las curvas de crecimiento para el segundo ensayo, ahora con
densidades poblacionales. Se evidencian tendencias similares a las observadas con
valores de absorbancias, la pendiente del medio 2 supera en forma ligera a las de 1 y
3, y en el medio 4 hubo un crecimiento muy lento y desfasado de los otros, por la
prolongada fase de latencia y adecuación. Las capacidades de carga no fueron
alcanzadas y no se encontraron diferencias significativas (α = 0,05) entre los medios.
Figura 24. Crecimiento poblacional de Arthrospira platensis en cuatro medios de cultivo (filamentos por litro) en la Cámara de Crecimiento.
0
50000
100000
150000
200000
250000
300000
350000
400000
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
Den
sid
ad (f
ilam
ento
s/L)
Tiempo (días)
Medio 1 Medio 2 Medio 3 Medio 4
A680 = 0,1471t + 0,0124 R2 = 0,9986
A680 = 0,1426t + 0,0404 R
2 = 0,9968
A680 = 0,1612t + 0,0136 R2 = 0,9969
A680 = 0,1619t + 0,0473 R2 = 0,9991

71
La Tabla 8 resume los principales resultados obtenidos en el ensayo. Las tasas de
crecimiento per cápita (r) se calcularon entre los días 7 y 10. Los mayores valores de
r, capacidad de carga (K) y producción de biomasa correspondieron al medio 1. La
población que creció en el medio 3 presenta una tasa per cápita ligeramente menor a
los medios 1 y 2. Los medios 1, 2 y 3 tuvieron valores de pH inicial y final muy
próximos con una tendencia al aumento, el medio 4 mostró valores similares.
Tabla 8. Tasa de crecimiento per cápita de Arthrospira platensis en la fase de crecimiento rápido, biomasa seca producida en cada medio de cultivo y pH inicial y final.
Medio Parámetros poblacionales Biomasa seca (g/L) pH
r (días-1
) K (filamentos/L) Inicial Final
1 0,50 412.900 0,840 9,12 10,15
2 0,48 390.300 0,797 9,17 10,16
3 0,30 321.800 0,717 9,19 10,04
4 0,01 – 0,243 9,70 9,86
En la siguiente etapa de escalamiento, se realizó el primer diseño multifactorial a cielo
abierto en botellones de 5 L. Los ensayos produjeron, en periodos de incubación
uniformes (28 días), curvas de crecimiento logísticas con amortiguaciones monótonas
y oscilaciones amortiguadas sostenidas y crecientes (figura 25).
La inmensa mayoría de los tratamientos siguieron la dinámica de una típica curva
sigmoidea, que gráficamente representa el modelo logístico de crecimiento
poblacional. Las curvas se caracterizaron por presentar una fase inicial de crecimiento
lento (latencia y adecuación), donde incluso en algunos cultivos se evidenciaron
decrecimientos entre los días 1 y 4. A esta fase de crecimiento lento o estacionario le
siguió otra de crecimiento rápido o exponencial, la cual duró tres (3) días en la mayoría
de los tratamientos. Finalmente, después del punto de inflexión de la curva el
crecimiento empezó a amortiguarse hasta alcanzar el estado de saturación (plateau),
donde la mayoría de las poblaciones se estabilizaron una vez fue alcanzada la
capacidad de carga del ambiente (medio de cultivo).
Varios tratamientos alcanzaron un equilibrio estable con atenuación monótona
(“monotonic damping”) en el valor de la capacidad de carga (K), mientras que algunas
poblaciones alcanzaron dicha capacidad de carga con oscilaciones, unas
amortiguadas y otras sostenidas, lo que puede atribuirse a algún retraso temporal en
el efecto de la densidad. Algunos tratamientos como el 6, 22, 26, 28 y 29 no
alcanzaron el plateau y los cultivos continuaron un crecimiento indefinido, en la

72
mayoría de los casos con marcadas oscilaciones. Por su parte, la curva del
tratamiento 19 alcanzó la capacidad de carga a una absorbancia muy inferior a los
medios más productivos, pero claramente muestra un patrón de oscilaciones
amortiguadas.
Figura 25. Curvas de crecimiento poblacional de Arthrospira platensis obtenidas en un ensayo
factorial fraccionado 26-1
con combinaciones aleatorias de niveles mínimos y centrales de cinco macronutrientes (tratamientos). La densidad poblacional fue medida indirectamente con la absorbancia a una longitud de onda de 680 nm, correspondiente al rojo dentro del espectro visible, pico de absorción de la clorofila a.
Tratamiento 1
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A6
80
nm
Tratamiento 2
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A6
80
nm
Tratamiento 3
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A6
80
nm
Tratamiento 4
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A6
80
nm
Tratamiento 5
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A6
80
nm
Tratamiento 6
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
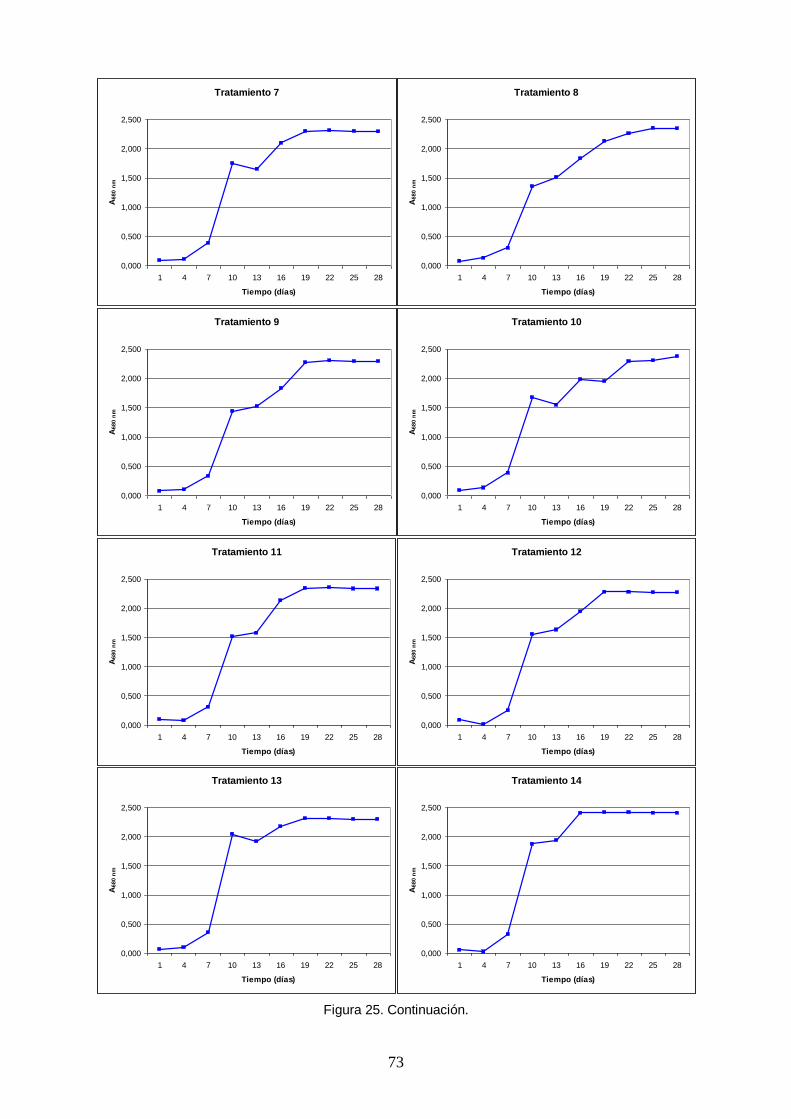
73
Figura 25. Continuación.
Tratamiento 7
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 8
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 9
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 10
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 11
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 12
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 13
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A6
80
nm
Tratamiento 14
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A6
80
nm

74
Figura 25. Continuación.
Tratamiento 17
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 18
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 19
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 20
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 21
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 22
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A6
80
nm
Tratamiento 15
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 16
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A6
80
nm
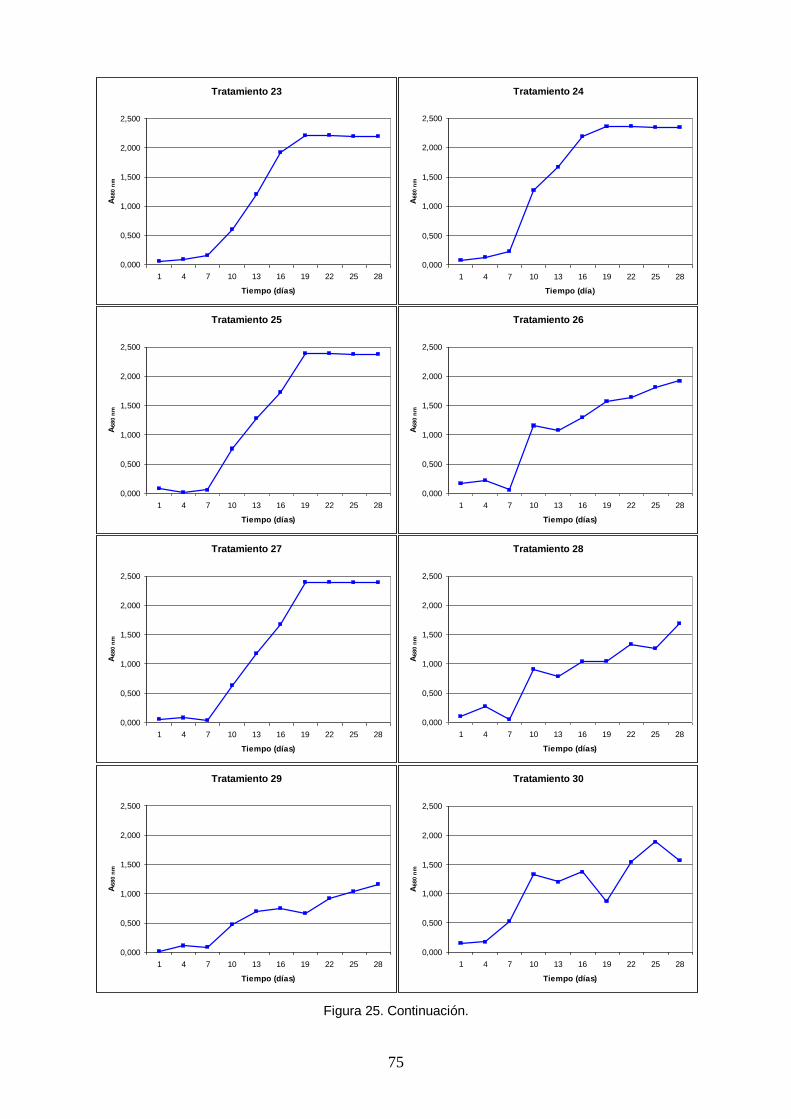
75
Figura 25. Continuación.
Tratamiento 23
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 24
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (día)
A680 n
m
Tratamiento 25
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 26
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 27
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 28
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 29
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 30
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
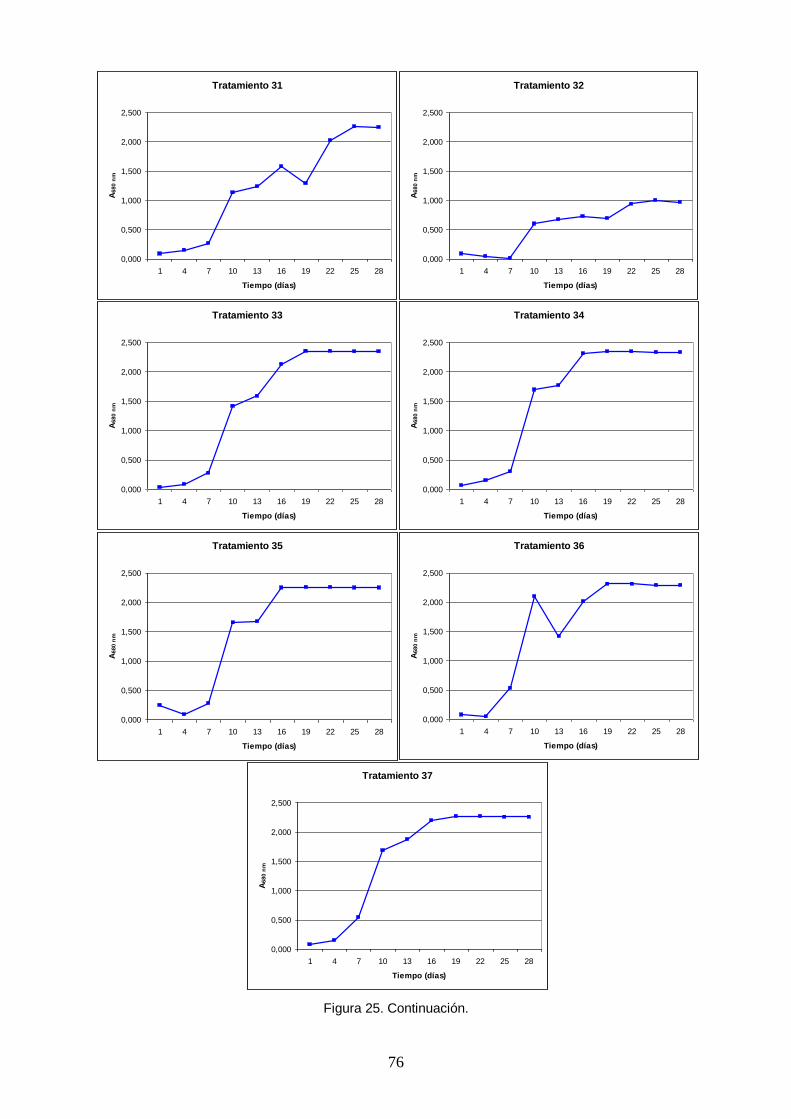
76
Figura 25. Continuación.
Tratamiento 31
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 32
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 33
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A6
80
nm
Tratamiento 34
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A6
80
nm
Tratamiento 35
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 36
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m
Tratamiento 37
0,000
0,500
1,000
1,500
2,000
2,500
1 4 7 10 13 16 19 22 25 28
Tiempo (días)
A680 n
m

77
La tasa per cápita de crecimiento, ΔN/[N(t)Δt], presenta un incremento notable en
algunas curvas para los primeros días. Casi todas las curvas coincidieron en el día 10
como tiempo tope del crecimiento aparentemente exponencial, donde es válido
aproximar la tasa per cápita de crecimiento al valor r. Posteriormente a este lapso de
tiempo, el crecimiento disminuye progresivamente hasta llegar al equilibrio en K. La
población decreció hasta cero (ΔN/Δt = 0), en todas los casos donde la capacidad de
carga, K, se alcanzó. Para algunos tratamientos se observaron decrecimientos en los
primeros tres días (ΔN/Δt < 0), antes del inicio del crecimiento rápido, principalmente
en las curvas correspondientes a los tratamientos 2, 3, 4, 12, 18, 25, 32 y 35, lo que
revela fases de latencia y adaptación marcadas por parte de las poblaciones de A.
platensis. La Tabla 9 resume las tasas de crecimiento per cápita (días-1) para tres
intervalos de tiempo en fase de crecimiento rápido, así como las capacidades de carga
(filamentos/L) para el método de conteo de filamentos.
Tabla 9. Tasas de crecimiento per cápita y capacidades de carga en cultivos de Arthrospira platensis para el ensayo factorial fraccionado 2
6-1. En rojo se destacan los medios con valores
mayores para uno o ambos parámetros poblacionales obtenidos de forma experimental.
Tratamiento Tasa de crecimiento per cápita (días-1
) Capacidad de carga, K
(filamentos/L)
Días 7 – 10 Días 10 – 13 Días 13 – 16
1 0,22 0,21 0,11 762.000
2 0,12 0,16 0,06 812.000
3 0,44 0,35 0,22 658.500
4 0,20 0,114 0,08 785.000
5 0,20 0,17 0,10 –
6 0,15 0,12 0,05 –
7 0,46 0,42 0,37 630.500
8 0,42 0,40 0,25 654.700
9 0,41 0,32 0,20 616.870
10 0,45 0,40 0,27 –
11 0,30 0,25 0,17 523.540
12 0,29 0,22 0,11 722.520
13 0,68 0,41 0,28 522.410
14 0,82 0,75 0,52 739.750
15 0,54 0,40 0,29 740.165
16 0,85 0,81 0,70 1.782.960
17 0,65 0,50 0,32 789.875
18 0,72 0,54 0,35 1.425.000
19 0,37 0,30 0,21 374.100
20 0,18 0,19 0,10 730.589
21 0,21 0,20 0,12 798.500

78
Tabla 9. Continuación.
Tratamiento Tasa de crecimiento per cápita (días-1
) Capacidad de carga,
K (filamentos/L)
Días 7 – 10 Días 10 – 13 Días 13 – 16
22 0,18 0,17 0,09 –
23 0,23 0,15 0,10 851.450
24 0,37 0,30 0,16 730.215
25 0,41 0,35 0,22 831.230
26 0,16 0,12 0,08 –
27 0,45 0,39 0,30 730.257
28 0,27 0,18 0,14 –
29 0,84 0,71 0,69 –
30 0,20 0,21 0,11 –
31 0,25 0,22 0,12 –
32 0,19 0,11 0,07 –
33 0,82 0,74 0,64 1.932.680
34 0,51 0,45 0,40 1.747.890
35 0,10 0,11 0,05 301500
36 0,75 0,65 0,51 2.520.000
37 0,70 0,51 0,32 2.050.000
La figura 26 describe en orden creciente los tiempos de duplicación (tg) para
poblaciones inoculadas en 37 botellones con los tratamientos preparados. Para la
obtención de estos valores se utilizó la tasa de recambio entre los días 7 y 10, en la
fase de crecimiento rápido, como valor de la tasa de crecimiento poblacional, dado que
al día 10 las poblaciones duplicaron sus tamaños. Algunos de los tratamientos con las
mejores dinámicas de crecimiento poblacional, como los tratamientos 14, 16, 18, 29,
33, 36 y 37 estuvieron entre los medios con tiempos generacionales más cortos (de
0,8 a 1 día), y algunos con dinámicas menos eficientes tuvieron tiempos
generacionales más prolongados, como 2, 6 y 32. El tratamiento 2 tuvo un tiempo de
duplicación muy prolongado (6,5 días), a pesar de ostentar una de las capacidades de
carga más elevadas, pero el gran retraso inicial producto de una latencia y adecuación
prolongadas, desplazó con mucho el inicio de la fase crecimiento rápido, alcanzando el
plateau aproximadamente a los 18 días, cuando poblaciones en otros medios lo
habían logrado de 10 a 15 días. Algo similar se puede decir del medio 35 que tuvo el
valor de tg más alto de todos los tratamientos. El medio 16 presentó el tiempo de
duplicación más corto (0,8 días), esto debido a una tasa de crecimiento per cápita muy
alta en la fase exponencial, lo que permitió a la población salir prontamente de una
breve fase de latencia y alcanzar el plateau un poco antes de los diez (10) días.
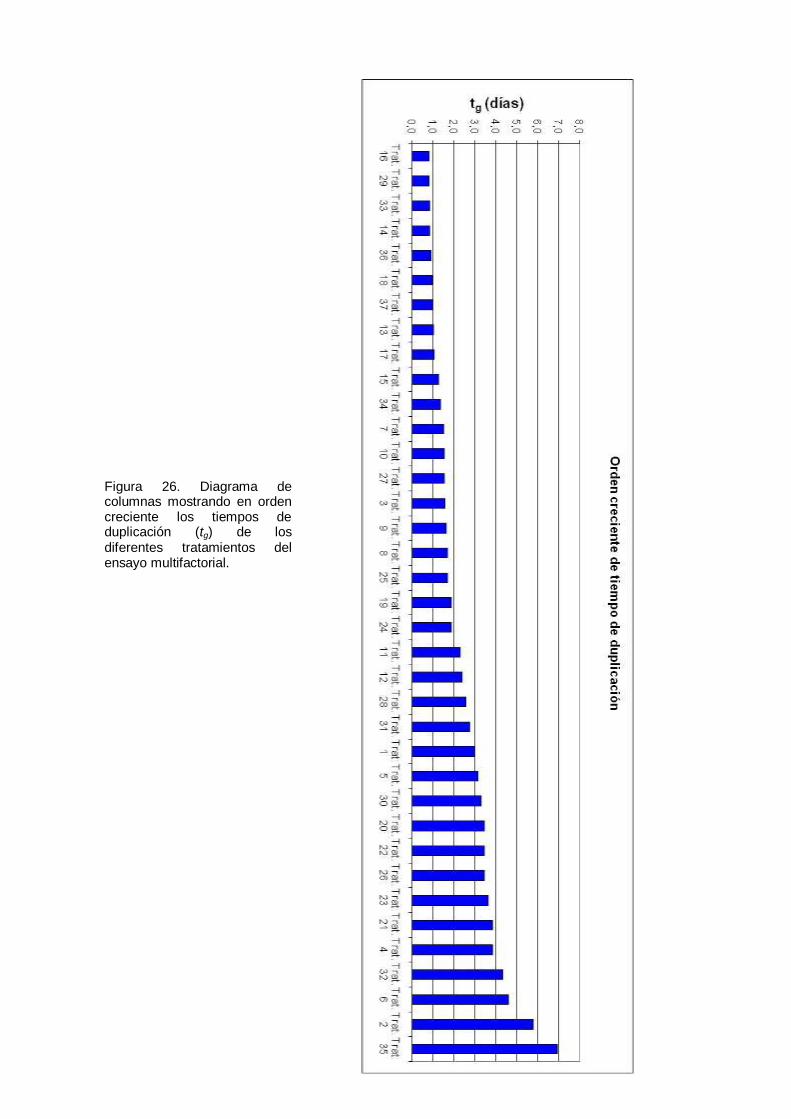
79
Figura 26. Diagrama de columnas mostrando en orden creciente los tiempos de duplicación (tg) de los diferentes tratamientos del ensayo multifactorial.

80
El medio más productivo en biomasa fue el tratamiento 36 (aprox. 2,3 g/L), mientras
que el tratamiento 29 resultó el menos rendidor. Varios de los medios (16, 18, 33 y 36)
que mostraron los tg más bajos, también se constituyeron en los más productivos en
biomasa seca (figura 27). Un tiempo de duplicación corto implica que la producción de
biomasa será elevada en un término de tiempo breve, a la vista están los resultados
que indican que los tratamientos con tiempos generacionales menores alcanzaron la
capacidad de carga en un número de días inferior, alrededor de 10 días, y entonces
los 20 días restantes del ensayo permitieron generar un rendimiento superior de
biomasa para fines de producción masiva a escala mayor.
El análisis de varianza para las diferentes combinaciones aleatorias y niveles centrales
de cinco macronutrientes, para la variable respuesta densidad óptica o absorbancia,
determinó diferencias significativas (α = 0,05) para las sales que constituyen la
solución I del medio Spirulina. Esto repercutió en forma determinante en las dinámicas
poblacionales tan variables que se observaron en cada uno de los medios nutritivos.
De este análisis estadístico se desprende que las fuentes de carbono (NaHCO3 y
Na2CO3) y fósforo (KH2PO4), así como el efecto del modelo o diseño empleado,
resultan concluyentes para evidenciar diferencias significativas en los crecimientos de
las poblaciones de A. platensis, donde hubo combinaciones aleatorias de esos
componentes químicos esenciales para el desarrollo y replicación de los filamentos. El
nitrógeno, otro nutriente relevante en la nutrición mineral de A. platensis, no tuvo
efecto significativo en las cinéticas evidenciadas (Tabla 10).
Tabla 10. Análisis de varianza para el diseño factorial fraccionado 26-1
(salida del programa Design Expert). Los números destacados en rojo son valores de p<0,05.
Fuente Suma de
Cuadrados
Grados de
libertad
Cuadrado
Medio F p
Modelo 4,18 16 0,26 4,88 0,0007
NaHCO3 1,19 1 1,19 22,32 0,0001
Na2CO3 0,43 1 0,43 8,12 0,0102
KH2PO4 0,25 1 0,25 4,76 0,0418
NaCl 8,32E+00 1 8,32E+00 0,16 0,6978
K2SO4 0,18 1 0,18 3,42 0,0799
KNO3 0,12 1 0,12 2,21 0,1532
A partir de estas determinaciones, se escogió el tratamiento 36 como el óptimo para
ser probado en ensayos a escala mayor. Esta decisión deriva del análisis de las
interacciones de la biomasa con las concentraciones milimolares de los nutrientes más
importantes en la ejecución del experimento, siendo el 36 uno de los tratamientos
centrales (ver Apéndice 2), con mejores valores de parámetros y biomasa.

81
Figura 27. Biomasa seca (g/L) cosechada en los diferentes tratamientos del ensayo factorial 2
6-1.
Los valores se ordenan en forma creciente.
Ord
en
cre
cie
nte
de
bio
ma
sa
se
ca
0,0
00
0,5
00
1,0
00
1,5
00
2,0
00
2,5
00
29
19
30
25
28
32
611
10
117
12
31
26
415
322
227
514
820
24
35
13
916
23
33
18
34
37
21
736
Tra
tam
ien
to
BS (g/L)
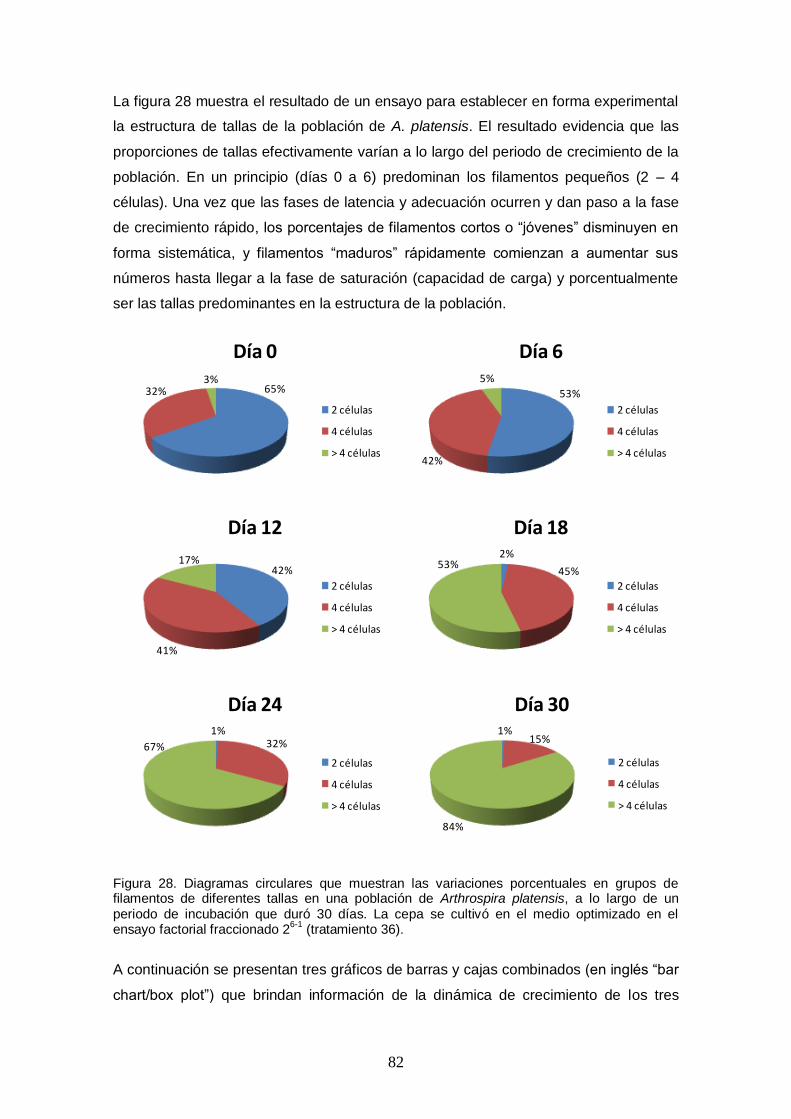
82
La figura 28 muestra el resultado de un ensayo para establecer en forma experimental
la estructura de tallas de la población de A. platensis. El resultado evidencia que las
proporciones de tallas efectivamente varían a lo largo del periodo de crecimiento de la
población. En un principio (días 0 a 6) predominan los filamentos pequeños (2 – 4
células). Una vez que las fases de latencia y adecuación ocurren y dan paso a la fase
de crecimiento rápido, los porcentajes de filamentos cortos o “jóvenes” disminuyen en
forma sistemática, y filamentos “maduros” rápidamente comienzan a aumentar sus
números hasta llegar a la fase de saturación (capacidad de carga) y porcentualmente
ser las tallas predominantes en la estructura de la población.
Figura 28. Diagramas circulares que muestran las variaciones porcentuales en grupos de filamentos de diferentes tallas en una población de Arthrospira platensis, a lo largo de un periodo de incubación que duró 30 días. La cepa se cultivó en el medio optimizado en el ensayo factorial fraccionado 2
6-1 (tratamiento 36).
A continuación se presentan tres gráficos de barras y cajas combinados (en inglés “bar
chart/box plot”) que brindan información de la dinámica de crecimiento de los tres
65%32%3%
Día 0
2 células
4 células
> 4 células
53%
42%
5%
Día 6
2 células
4 células
> 4 células
42%
41%
17%
Día 12
2 células
4 células
> 4 células
2%
45%53%
Día 18
2 células
4 células
> 4 células
1%32%67%
Día 24
2 células
4 células
> 4 células
1%15%
84%
Día 30
2 células
4 células
> 4 células

83
componentes poblacionales de filamentos discriminados por sus tallas y la dispersión
de los datos (desviaciones estándares), respectivamente. Se verifica que la cohorte de
filamentos con dos células tiene un crecimiento poblacional vigoroso y sostenido hasta
el día 12 cuando alcanza un máximo, y desde este punto crítico empieza a declinar de
forma rápida hasta valores ínfimos al final del periodo cuando la población se
estabiliza y es madura. Los filamentos con cuatro células tienen una dinámica similar
(patrón unimodal) a la exhibida en el perfil de los filamentos más cortos de dos células,
pero con un desfase, alcanzando el máximo el día 18 y luego, como en el caso
anterior, declina la población a niveles basales hacia postrimerías del tiempo de
crecimiento asintótico. El tercer conjunto de tallas, correspondiente a filamentos con
más de cuatro células, posee una dinámica tardía respecto a los dos primeros, puesto
que la población se incrementa luego de que se han formado muchos filamentos
cortos que empiezan a crecer radialmente y sumar células al filamento, de este modo
la población de filamentos grandes empieza a ser ampliamente dominante al final del
periodo de crecimiento y se estabiliza con ligeras fluctuaciones en el estado de
saturación. Los datos medios, que son los que se grafican, presentaron poca
dispersión, lo que puede mostrarse como una aproximación experimental a una
estructura de tallas de la población de A. platensis (figura 29).
Figura 29. Dinámicas de crecimiento poblacional de los tres componentes de tallas de filamentos de Arthrospira platensis: (a) filamentos con 2 células, (b) filamentos con cuatro células y (c) filamentos con más de cuatro células. La letra “Y” en la ordenada es la densidad (filamentos/L) y en la abscisa el tiempo está dividido en intervalos de tres días. Salida: PAST.
0 3 6 9 12 15 18 21 24 27 30
0
3E05
6E05
9E05
1,2E06
1,5E06
1,8E06
2,1E06
2,4E06
2,7E06
Y
0 3 6 9 12 15 18 21 24 27 30
0
8E04
1,6E05
2,4E05
3,2E05
4E05
4,8E05
5,6E05
6,4E05
7,2E05
Y
0 3 6 9 12 15 18 21 24 27 30
0
2E05
4E05
6E05
8E05
1E06
1,2E06
1,4E06
1,6E06
1,8E06
Y
(a) (b)
(c)

84
Por último, para poner a prueba la idoneidad del modelo logístico como modelo
dinámico para A. platensis, se hizo una prueba sencilla de extracción de volúmenes a
partir de la saturación o capacidad de carga (K), y reemplazar el volumen retirado con
medio de cultivo optimizado (tratamiento 36). Esta aproximación experimental permitió
medir en cuanto tiempo la población retornó a K en las siguientes situaciones: (1)
retiro de un volumen por encima del punto de inflexión (N = K/2), correspondiente a
25% del volumen total del cultivo; (2) retirar la mitad de la población (50%), para llevar
a la población hasta el punto de inflexión; (3) extracción del 75% del volumen total, que
implica cosechar la mayor parte de la población en una concentración muy por debajo
del punto de inflexión. Se puede observar que en el primer caso, a pesar de tratarse
de una remoción cercana al punto de saturación, su pendiente es inferior a la obtenida
para una remoción de 50%, lo que evidencia que el segundo tratamiento reduce a la
población justo a la fase de crecimiento rápido, mientras que con un 25% de remoción
las condiciones son similares o próximas a la saturación. El tratamiento con una
reducción de 75% de la población total llevó a los filamentos remanentes a un tamaño
basal, y la recuperación evidentemente se hizo muy lenta y, al momento de que en los
otros tratamientos las poblaciones alcanzaron y se estabilizaron en la capacidad de
carga, en el tercer caso la población aún no salía de la fase de latencia. El tiempo en
el que la población en el primer tratamiento (25%) retornó a K fue de cinco días, y con
una remoción de 50% la población tardó igual número de días.
Figura 30. Dinámicas de crecimiento poblacional de Arthrospira platensis a partir de tres extracciones de volúmenes en fase de saturación. De izquierda a derecha se muestran las curvas de crecimiento a partir de 25%, 50% y 75% de extracción.
0,000
0,500
1,000
1,500
2,000
2,500
3,000
1 3 5 7 9 11 13 15 1 3 5 7 9 11 13 15 1 3 5 7 9 11 13 15
A6
80
Tiempo (días)
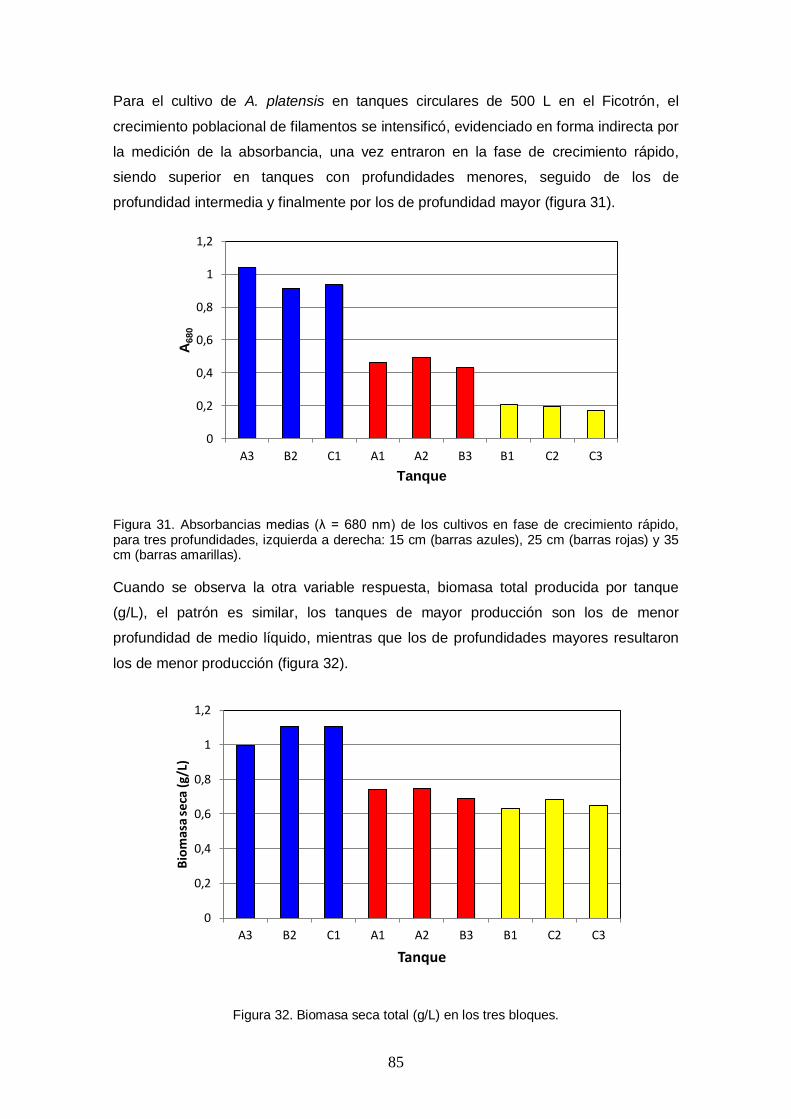
85
Para el cultivo de A. platensis en tanques circulares de 500 L en el Ficotrón, el
crecimiento poblacional de filamentos se intensificó, evidenciado en forma indirecta por
la medición de la absorbancia, una vez entraron en la fase de crecimiento rápido,
siendo superior en tanques con profundidades menores, seguido de los de
profundidad intermedia y finalmente por los de profundidad mayor (figura 31).
Figura 31. Absorbancias medias (λ = 680 nm) de los cultivos en fase de crecimiento rápido, para tres profundidades, izquierda a derecha: 15 cm (barras azules), 25 cm (barras rojas) y 35 cm (barras amarillas).
Cuando se observa la otra variable respuesta, biomasa total producida por tanque
(g/L), el patrón es similar, los tanques de mayor producción son los de menor
profundidad de medio líquido, mientras que los de profundidades mayores resultaron
los de menor producción (figura 32).
Figura 32. Biomasa seca total (g/L) en los tres bloques.
0
0,2
0,4
0,6
0,8
1
1,2
A3 B2 C1 A1 A2 B3 B1 C2 C3
A680
Tanque
0
0,2
0,4
0,6
0,8
1
1,2
A3 B2 C1 A1 A2 B3 B1 C2 C3
Bio
mas
a se
ca (
g/L)
Tanque

86
Es indiscutible pensar que la profundidad de la lámina de agua en los tanques
circulares afecta de manera diferencial la tasa de crecimiento y producción de
biomasa. El análisis de varianza determinó que hay diferencias significativas (α = 0,05)
entre los tratamientos (profundidades). Por otro lado, el efecto de los bloques en el
experimento parece no ser relevante (Tabla 11).
Tabla 11. Resumen del Análisis de Varianza (salida de MICROSOFT EXCEL).
Fuente de Variación Suma de
Cuadrados
Grados de
libertad
Media de
cuadrados Fo
Tratamientos (profundidad) 4197,31 2 2098,66 11,82
Bloques (columnas) 556,03 2 278,01 1,57
Error 709,95 4 177,49
Total 5463,29 8
F0,05;2;4=6,94
Para cultivadores tipo carrusel (2500 L de capacidad) se tiene información limitada de
su dinámica y producción de biomasa. La tasa de crecimiento per cápita media del
cultivo en carrusel, como aproximación a la tasa instantánea, fue 0,25 días-1 y la
biomasa seca total 0,5 g/L, en la única cosecha realizada. Se puede advertir cómo
disminuyen la tasa de crecimiento per cápita media y la producción de biomasa desde
botellones hasta el carrusel en condiciones no controladas y con fotoperiodo (sólo el
intervalo de horas con luz natural).
7.3 Comunidades fitoplanctónicas
El estudio de comunidades fitoplanctónicas ofrece la oportunidad de explorar los
ambientes acuáticos variados del país, así como colectar numerosas especies de
cianobacterias y microalgas eucarióticas con gran potencial biotecnológico.
Ambientes dulceacuícolas. La Tabla 12 presenta la composición y abundancia de la
biota fitoplanctónica de un humedal herbáceo de la península de Paria (Palmares III).
El fitoplancton estuvo principalmente representado, en riqueza y abundancia, por
diatomeas (Bacillariophyta) y algas verde-azules (Cyanobacteria). La zona de
Brachiaria mutica (Poaceae) fue la comunidad vegetal con mayor número de especies
(24), mientras que el ecotono Cyperus articulatus (Cyperaceae) - Typha dominguensis
(Typhaceae) fue la vegetación con riqueza menor (8).
Las especies de diatomeas cuantitativamente importantes fueron: Gyrosigma
attenuatum, muy común en B. mutica y el ecotono T. dominguensis - Sesbania

87
exasperata (Fabaceae); tres especies del género Navicula con densidades
poblacionales grandes en zonas diversas, en especial N. platalea con una densidad
notable en el ecotono S. exasperata - C. articulatus; Nitzschia valens, abundante en B.
mutica, S. exasperata y C. articulatus. Otras diatomeas, como Pinnularia gibba y
Pleurosigma sp., fueron numerosas en B. mutica, pero se hicieron raras o ausentes en
las demás zonas de vegetación. Oscillatoria sp. fue la única cianobacteria abundante y
dominante, con densidades grandes en B. mutica, S. exasperata y los ecotonos B.
mutica - S. exasperata y T. dominguensis - S. exasperata. Otra cianobacteria,
Anabaena sp., fue muy importante en el parche de B. mutica. También se encontraron
ejemplares de Spirulina subsalsa, Arthrospira sp y Chlorella sp. Las algas verdes
(Chlorophyta) y euglenofitas (Euglenophyta) escasearon en todas las comunidades
vegetales. Se halló un género de dinoflagelado (Pyrrophyta) con algunos individuos de
la especie Protoperidinium sp. en el ecotono S. exasperata - C. articulatus.
Tabla 12. Composición de especies y abundancia (células/litro) de los taxa fitoplanctónicos presentes en las zonas de vegetación en noviembre de 2008. Bm (Brachiaria mutica), EcBmSe (ecotono B. mutica - Sesbania exasperata), Se (S. exasperata), EcSeCa (ecotono S. exasperata - Cyperus articulatus), Ca (C. articulatus), EcCaTd (ecotono C. articulatus-Typha dominguensis), Td (T. dominguensis) y EcTdSe (ecotono T. dominguensis - S. exasperata).
TAXA Bm EcBmSe Se EcSeCa Ca EcCaTd Td EcTdSe Se
Bacillariophyta (15 especies)
Cocconeis striata 0 0 0 0 4 0 0 0 0
Eunotia monodon 0 0 8 0 8 4 8 0 8
Fragilaria crotonensis 5 2 2 1 1 0 0 1 2
Frustulia sp. 8 4 4 2 1 4 2 0 0
Gyrosigma attenuatum 264 80 24 0 40 20 4 236 24
Gomphonema brasiliense 0 0 0 0 4 0 0 0 0
Navicula fulva 72 12 32 32 52 20 0 0 32
N. oblonga 48 14 40 20 12 4 0 4 40
N. platalea 40 8 64 336 64 2 0 0 64
Nitzschia valens 80 50 64 24 96 40 24 0 64
N. obtusa 0 0 0 0 8 4 0 0 0
Pinnularia gibba 68 8 4 0 8 0 0 0 4
Pleurosigma sp. 48 20 0 0 0 0 4 0 0
Surirella sp. 0 0 0 0 8 0 4 0 0
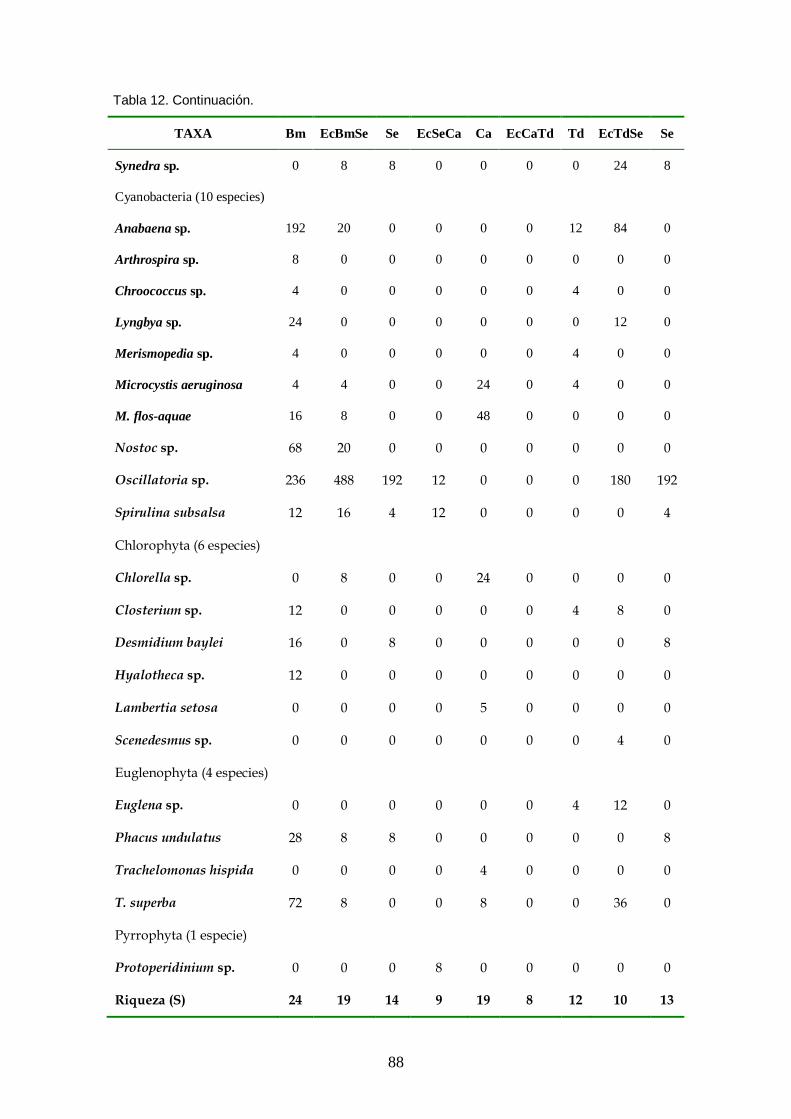
88
Tabla 12. Continuación.
TAXA Bm EcBmSe Se EcSeCa Ca EcCaTd Td EcTdSe Se
Synedra sp. 0 8 8 0 0 0 0 24 8
Cyanobacteria (10 especies)
Anabaena sp. 192 20 0 0 0 0 12 84 0
Arthrospira sp. 8 0 0 0 0 0 0 0 0
Chroococcus sp. 4 0 0 0 0 0 4 0 0
Lyngbya sp. 24 0 0 0 0 0 0 12 0
Merismopedia sp. 4 0 0 0 0 0 4 0 0
Microcystis aeruginosa 4 4 0 0 24 0 4 0 0
M. flos-aquae 16 8 0 0 48 0 0 0 0
Nostoc sp. 68 20 0 0 0 0 0 0 0
Oscillatoria sp. 236 488 192 12 0 0 0 180 192
Spirulina subsalsa 12 16 4 12 0 0 0 0 4
Chlorophyta (6 especies)
Chlorella sp. 0 8 0 0 24 0 0 0 0
Closterium sp. 12 0 0 0 0 0 4 8 0
Desmidium baylei 16 0 8 0 0 0 0 0 8
Hyalotheca sp. 12 0 0 0 0 0 0 0 0
Lambertia setosa 0 0 0 0 5 0 0 0 0
Scenedesmus sp. 0 0 0 0 0 0 0 4 0
Euglenophyta (4 especies)
Euglena sp. 0 0 0 0 0 0 4 12 0
Phacus undulatus 28 8 8 0 0 0 0 0 8
Trachelomonas hispida 0 0 0 0 4 0 0 0 0
T. superba 72 8 0 0 8 0 0 36 0
Pyrrophyta (1 especie)
Protoperidinium sp. 0 0 0 8 0 0 0 0 0
Riqueza (S) 24 19 14 9 19 8 12 10 13

89
Las comunidades fitoplanctónicas de Palmares III no sólo variaron en forma cualitativa
(composición de especies) en los diferentes parches de vegetación emergente, los
índices de diversidad revelan que también cambiaron en forma cuantitativa a lo largo
de los ambientes parcelados. Para 2008, la diversidad, expresada en el índice
Shannon-Wiener (H´), relativamente fue alta en los parches de B. mutica (Bm) y C.
articulatus (Ca) debido a que la riqueza y equidad, sus dos componentes, también
fueron elevadas; en el caso de T. dominguensis (Td), con el tercer valor más alto, el
ambiente mostró la equidad más prominente del humedal en esa época. Los ecotonos
de vegetación o ambientes de transición fueron menos diversos que los parches
monoespecíficos contiguos. En el ecotono S. exasperata – C. articulatus (EcSeCa) la
equidad se constituyó en la más baja, subsecuentemente los índices de dominancia de
Simpson y Berger-Parker tuvieron valores sobresalientes (Tabla 13).
Tabla 13. Algunos índices de diversidad empleados para caracterizar la comunidad
fitoplanctónica de Palmares III en sus diferentes parches de vegetación monoespecífica y ecotonos (noviembre 2008). Salida: PAST.
Índice Bm EcBmSe Se EcSeCa Ca EcCaTd Td EcTdSe
Riqueza (S) 24 19 14 9 19 8 12 11
Equidad (J´) 0,80 0,54 0,72 0,46 0,81 0,78 0,88 0,66
Shannon-Wiener (H´) 2,55 1,60 1,89 1,00 2,38 1,62 2,20 1,59
Dominancia de Simpson (D) 0,11 0,40 0,23 0,58 0,12 0,26 0,15 0,27
Berger-Parker (d) 0,20 0,62 0,42 0,75 0,23 0,41 0,31 0,39
La Tabla 14 muestra los valores del índice de distancia o similitud Jaccard, para el
conjunto de ambientes parcelados (asociaciones de plantas emergentes) del humedal
Palmares III en noviembre de 2008. En términos generales, se evidencian similitudes
notables entre algunos parches monoespecíficos con ecotonos vecinos,
principalmente en el gradiente B. mutica (Bm) ecotono B. mutica – S. exasperata
(EcBmSe) S. exasperata (Se). Es curioso que el ecotono S. exasperata – C.
articulatus (EcSeCa) sea tan diferente del parche monoespecífico de C. articulatus
(Ca), cuando la otra vegetación monoespecífica que converge en esa área del
humedal guarda una similitud notable con la misma zona ecotonal o mixta entre ambas
especies vegetales. Las formaciones de vegetación espacialmente más apartadas
entre sí en el humedal tuvieron similitudes moderadas a bajas. Los ambientes más
disímiles fueron el ecotono S. exasperata – C. articulatus y la zona monoespecífica de
T. dominguensis (Td).
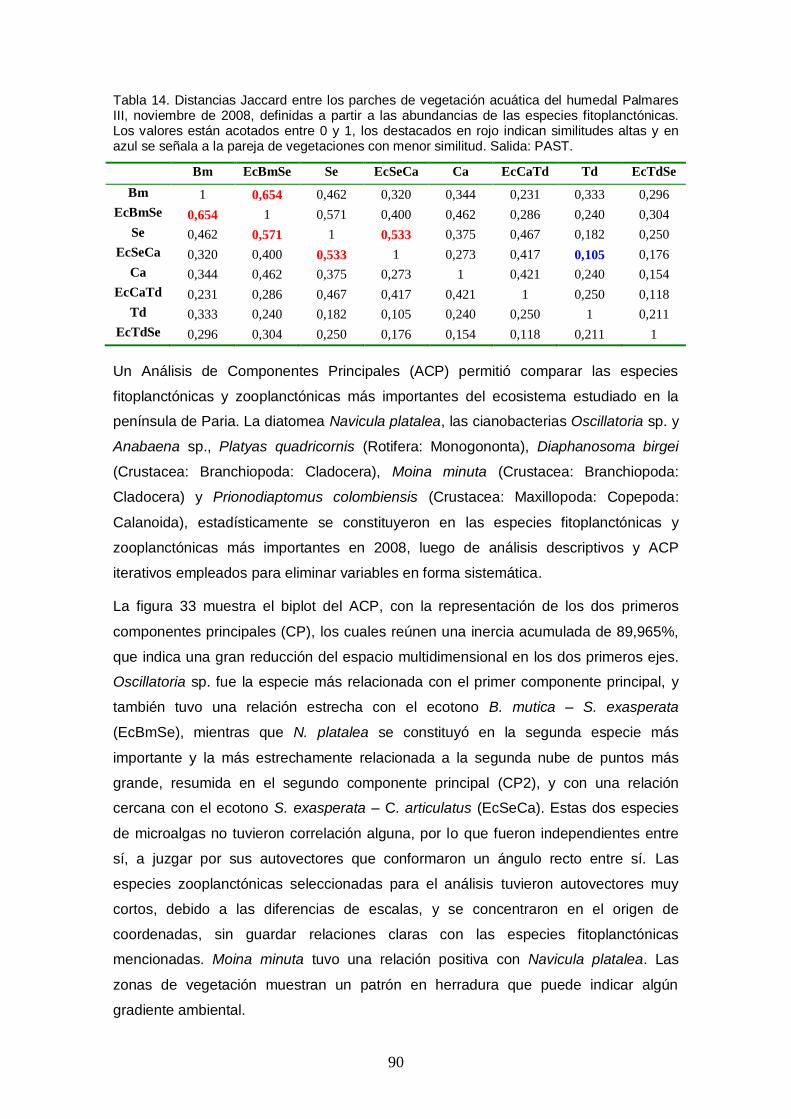
90
Tabla 14. Distancias Jaccard entre los parches de vegetación acuática del humedal Palmares III, noviembre de 2008, definidas a partir a las abundancias de las especies fitoplanctónicas. Los valores están acotados entre 0 y 1, los destacados en rojo indican similitudes altas y en azul se señala a la pareja de vegetaciones con menor similitud. Salida: PAST.
Bm EcBmSe Se EcSeCa Ca EcCaTd Td EcTdSe
Bm 1 0,654 0,462 0,320 0,344 0,231 0,333 0,296
EcBmSe 0,654 1 0,571 0,400 0,462 0,286 0,240 0,304
Se 0,462 0,571 1 0,533 0,375 0,467 0,182 0,250
EcSeCa 0,320 0,400 0,533 1 0,273 0,417 0,105 0,176
Ca 0,344 0,462 0,375 0,273 1 0,421 0,240 0,154
EcCaTd 0,231 0,286 0,467 0,417 0,421 1 0,250 0,118
Td 0,333 0,240 0,182 0,105 0,240 0,250 1 0,211
EcTdSe 0,296 0,304 0,250 0,176 0,154 0,118 0,211 1
Un Análisis de Componentes Principales (ACP) permitió comparar las especies
fitoplanctónicas y zooplanctónicas más importantes del ecosistema estudiado en la
península de Paria. La diatomea Navicula platalea, las cianobacterias Oscillatoria sp. y
Anabaena sp., Platyas quadricornis (Rotifera: Monogononta), Diaphanosoma birgei
(Crustacea: Branchiopoda: Cladocera), Moina minuta (Crustacea: Branchiopoda:
Cladocera) y Prionodiaptomus colombiensis (Crustacea: Maxillopoda: Copepoda:
Calanoida), estadísticamente se constituyeron en las especies fitoplanctónicas y
zooplanctónicas más importantes en 2008, luego de análisis descriptivos y ACP
iterativos empleados para eliminar variables en forma sistemática.
La figura 33 muestra el biplot del ACP, con la representación de los dos primeros
componentes principales (CP), los cuales reúnen una inercia acumulada de 89,965%,
que indica una gran reducción del espacio multidimensional en los dos primeros ejes.
Oscillatoria sp. fue la especie más relacionada con el primer componente principal, y
también tuvo una relación estrecha con el ecotono B. mutica – S. exasperata
(EcBmSe), mientras que N. platalea se constituyó en la segunda especie más
importante y la más estrechamente relacionada a la segunda nube de puntos más
grande, resumida en el segundo componente principal (CP2), y con una relación
cercana con el ecotono S. exasperata – C. articulatus (EcSeCa). Estas dos especies
de microalgas no tuvieron correlación alguna, por lo que fueron independientes entre
sí, a juzgar por sus autovectores que conformaron un ángulo recto entre sí. Las
especies zooplanctónicas seleccionadas para el análisis tuvieron autovectores muy
cortos, debido a las diferencias de escalas, y se concentraron en el origen de
coordenadas, sin guardar relaciones claras con las especies fitoplanctónicas
mencionadas. Moina minuta tuvo una relación positiva con Navicula platalea. Las
zonas de vegetación muestran un patrón en herradura que puede indicar algún
gradiente ambiental.

91
Figura 33. Biplot de los dos primeros componentes principales del ACP para las variables (especies) y casos (parches de vegetación) escogidas para caracterizar el ecosistema del humedal Palmares III en noviembre de 2008. Los dos primeros componentes principales acumularon 89,965% de la inercia total del sistema. Salida: MVSP 3.0.
El dendrograma (figura 34) del análisis de agrupamiento derivado de las especies
fitoplanctónicas y zooplanctónicas que destacaron en el ACP, muestra que se
formaron tres (3) grupos: (1) ecotono B. mutica – S. exasperata discriminado por
Oscillatoria sp. con el valor de centroide más alto; (2) ecotono S. exasperata – C.
articulatus (EcSeCa) al cual está asociada N. platalea y (3) B. mutica, S. exasperata y
ecotono T. dominguensis – S. exasperata, C. articulatus, ecotono C. articulatus – T.
dominguensis y T. dominguensis, con asociación de M. minuta. La vegetación
monoespecífica de T. dominguensis ecológicamente es muy parecida al ecotono que
esta especie forma con C. articulatus. La mayor parte de los otros ambientes poseen
disimilitudes de mediana magnitud en la escala Euclidiana. Los dos ecotonos que
conforman el segundo y tercer grupo, respectivamente, son ambientes muy diferentes
entre sí y a cualquier otra comunidad de plantas acuáticas que existen en el humedal.
CP 2
CP 1
Bm
EcBmSe
Se
EcSeCa
Ca
EcCaTdTd
EcTdSe
-28
-56
-84
28
56
84
112
140
-33.21-66.41-99.62 33.21 66.41 99.62 132.82 166.03
Navicula platalea
Anabaena sp.
Oscillatoria sp.
Platyas quadricornis
Diaphanosoma birgei
Moina minuta
Prionodiaptomus colombiensis
Vector scaling: 146,94

92
Figura 34. Dendrograma del Análisis de Agrupamiento o “Cluster Analysis” derivado del conjunto de especies fitoplanctónicas y zooplanctónicas más importantes colectadas en el ACP. Salida: MVSP 3.0.
En cuanto al ambiente abiótico, las variables fisicoquímicas mostraron una cierta
uniformidad, sólo algunas oscilaciones en el oxígeno disuelto. Los valores de salinidad
resultaron importantes para establecer que el humedal es un ambiente con
característica salobre. Se adicionan datos de algunos iones importantes para la
caracterización química del agua del humedal, principalmente metales alcalinos con
concentraciones importantes de sodio y muy bajas de potasio, que robustecen la idea
del carácter salobre del ambiente, y alcalino-térreos (cantidades notables de calcio y
magnesio), y aniones como sulfatos y cloruros en concentraciones altas (Tabla 15).
Tabla 15. Variables fisicoquímicas y concentraciones de cationes y aniones del agua, determinados en las zonas de vegetaciones monoespecíficas estudiadas en el humedal de Palmares en noviembre de 2008.
Parámetro/ ion B. mutica S. exasperata C. articulatus T. dominguensis
pH 7,3 6,9 7,1 7,3 Conductividad (mS/cm) 2,3 2,3 2,7 3,5 O2 (mg/L) 1,4 2,3 3,2 4,5 T (˚C) 25,1 25,4 24,6 25,8 Salinidad (‰) 0,11 0,11 0,12 0,13 Profundidad (cm) 23,8 21,8 19,0 18,6 Ca2+ (mg/L) 93,0 94,0 118,0 112,0 Mg2+ (mg/L) 64,0 55,0 58,0 60,0 Na+ (mg/L) 255,0 285,0 258,0 325,0 K+ (mg/L) 7,8 7,1 9,8 4,6 Cl- (mg/L) 540,0 500,0 460,0 660,0 SO4
2- (mg/L) 237,0 257,0 343,0 298,0

93
En agosto de 2009, un segundo muestreo en el mismo humedal de Paria, presentó
características fisonómicas cambiantes en la vegetación, debido a un incendio en
sequía que diezmó buena parte de la cobertura herbácea original, la cual cubría 100%
de su superficie, sin formar espejos de agua. El fitoplancton estuvo representado casi
en su totalidad por cianobacterias y euglenofitas, con predominio de las primeras en el
ecotono B. mutica - C. articulatus y abundancia de las segundas en B. mutica y T.
dominguensis. En términos generales, la riqueza fue muy inferior a la observada en
noviembre. Hubo un dominio alternado de una especie de euglenofita (Trachelomonas
superba) y una cianobacteria (Oscillatoria sp.). En esta época nuevamente se
encontraron representantes del género Arthrospira. Las diatomeas, en general,
escasearon en todos los ambientes, la única especie resaltante de este grupo fue N.
fulva en la zona B. mutica y el ecotono B. mutica - C. articulatus. Se observaron muy
pocas clorofitas. No se encontró fitoplancton en la zona de C. articulatus (Tabla 16).
Tabla 16. Composición de especies y abundancias (células/litro) de los taxa fitoplanctónicos presentes en las zonas de vegetación para agosto de 2009. EcBm1Ca y EcCaBm2 (ecotonos B. mutica - C. articulatus); EcBm2Td (ecotono B. mutica - T. dominguensis).
TAXA Bm1 EcBm1Ca Ca EcCaBm2 Bm2 EcBm2Td Td
Bacillariophyta (4 especies) Gyrosigma attenuatum 0 0 0 0 0 0 40 Navicula fulva 100 100 0 100 100 23 20 Nitzschia valens 60 20 0 20 60 5 0 Pinnularia gibba 20 20 0 20 20 0 0 Cyanobacteria (5 especies) Arthrospira sp. 20 0 0 0 20 0 0 Chroococcus sp. 0 20 0 20 0 0 0 Lyngbya sp. 20 0 0 0 0 0 0 Microcystis aeruginosa 0 20 0 20 0 0 0 Oscillatoria sp. 0 7560 0 7560 0 0 0 Chlorophyta (1 especie) Closterium sp. 20 0 0 0 20 0 0 Euglenophyta (2 especies) Trachelomonas hispida 0 20 0 20 0 0 0 T. superba 10.000 300 0 300 10.000 450 7680
Riqueza (S) 7 8 0 7 8 3 3
La Tabla 17 muestra los índices de diversidad empleados para caracterizar la
estructura comunitaria fitoplanctónica de Palmares III en agosto de 2009. Las riquezas
fueron muy bajas en todos los ambientes, por lo que la equidad pasó a tener
preponderancia o peso mayor en el valor del índice Shannon-Wiener. Hubo zonas de
vegetación como los dos parches de B. mutica (Bm), observados en esa época, con
equidades muy altas que produjeron los índices de diversidad más altos para el
humedal. El resto de los ambientes exhibieron valores de equidad muy bajos con la
subsecuente disminución de la diversidad y aumento de los índices de dominancia de
Simpson y Berger-Parker.

94
Tabla 17. Algunos índices de diversidad empleados para caracterizar la comunidad fitoplanctónica de Palmares III en sus diferentes parches de vegetación monoespecífica y ecotonos (agosto 2009). Salida: PAST.
Índice Bm1 EcBm1Ca Ca Bm2 EcBm2Td Td
Riqueza (S) 7 8 0 6 3 3
Equidad (J´) 0,85 0,15 0 0,83 0,23 0,05
Shannon-Wiener (H´) 1,65 0,31 0 1,49 0,25 0,05
Dominancia de Simpson (D) 0,76 0,12 0 0,72 0,11 0,02
Berger-Parker (d) 0,40 0,94 0 0,43 0,94 0,99
El índice de similitud o distancia Jaccard señala que la mayor similitud entre bandas de
vegetación, diferentes a ellas mismas, se encontró entre los dos parches de B. mutica
(Bm1 y Bm2). La segunda pareja de ambientes parcelados con mayor similitud
ecológica es la establecida entre B. mutica (Bm2) y su ecotono con T. dominguensis
(EcBm2Td). Los demás ambientes difirieron en forma notable y en distintos grados
(Tabla 18).
Tabla 18. Distancias Jaccard entre los parches de vegetación acuática del humedal Palmares III, agosto de 2009, definidas a partir a las abundancias de las especies fitoplanctónicas. Los valores están acotados entre 0 y 1, los destacados en rojo indican similitudes altas y en azul se señala a la pareja de vegetaciones con menor similitud. Salida: PAST.
Bm1 EcBm1Ca Bm2 EcBm2Td Td
Bm1 1 0,364 0,857 0,429 0,250
EcBm1Ca 0,364 1 0,400 0,375 0,222
Bm2 0,857 0,400 1 0,500 0,286
EcBm2Td 0,429 0,375 0,500 1 0,500
Td 0,250 0,222 0,286 0,500 1
Las variables físicas y químicas exhibieron cierta uniformidad en agosto de 2002. Se
encontraron incrementos leves de la salinidad, conductividad, temperatura y
profundidad hacia la zona de T. dominguensis. La concentración de oxígeno en el
agua también mostró un aumento hacia esa vegetación, aunque más notable que los
otros factores abióticos. El pH se mostró más alto en el parche de C. articulatus, sin
embargo en todas las comunidades se mantuvo alrededor de la neutralidad. En cuanto
a la intensidad de luz, se obtuvieron diferencias grandes entre la cantidad de luz que
llegó al techo vegetal y la que las plantas dejaron filtrar hasta la lámina de agua. La
zona de C. articulatus generó la disminución mayor de densidad fotónica (luminancia),
al sólo permitir que se filtraran 2 lux hasta la superficie del agua, de los 70 lux que
llegaron al dosel (Tabla 19).

95
Tabla 19. Variables ambientales medidas en agosto de 2009.
ZONA pH Conductividad
(mS/cm)
Temperatura
(°C)
Salinidad
(‰)
Profundidad
(cm)
Oxígeno
(mg/L)
Luminancia (lux)
Sobre el
techo
Bajo el
techo
B. mutica 6,93 2,30 26,92 0,11 18,75 0,88 56 8
Ecotono B.
mutica-C.
articulatus
6,92 2,33 26,90 0,11 15,00 0,74 53 14
C. articulatus 7,50 2,39 25,55 0,11 17,50 0,44 70 2
Ecotono B.
mutica-T.
dominguensis
7,41 3,05 31,47 0,14 16,00 2,68 65 19
T.
dominguensis 7,31 3,10 30,78 0,15 29,00 3,02 60 38
En la depresión de Barlovento, Edo. Miranda, específicamente en un sector
denominado El Clavo, se muestrearon dos zonas contiguas de vegetación emergente
monoespecífica: Heliconia marginata (Heliconiaceae) e Hymenachne amplexicaulis
(Poaceae), en sentido suroeste – noreste en la parte más estrecha de un humedal
herbáceo que ocupa la parte más baja (estero) de una zona boscosa con pendientes
pronunciadas. Este estudio se realizó en enero de 2009 a comienzo de sequía, pero
aún con algunas lluvias copiosas. En una zona pequeña de espejo de agua, el
fitoplancton tuvo una abundancia importante, aunque con riqueza baja, y dominio
amplio de dos especies: la cianobacteria Lyngbya lutea (500 filamentos/L) y en
segundo término la clorofita Spirogyra ternata (100 filamentos/L). Asterococcus
limneticus fue común en la zona de H. amplexicaulis. En las dos zonas con vegetación
emergente, las densidades poblacionales de las dos primeras especies y de algunas
otras cianobacterias, clorofitas y diatomeas fueron muy inferiores al espejo (Tabla 20).
Tabla 20. Grupos (Taxa) fitoplanctónicos identificados en los diferentes ambientes de vegetación en El Clavo, Barlovento, Edo. Miranda (enero de 2009).
Zona de
muestreo Muestra División Especie
Nivel de
organización
Abundancia
(células/L;
colonias/L;
filamentos/L)
Heliconia
marginata
He1
Cyanobacteria Microcoleus
chthonoplastes Filamentoso 1
Chlorophyta Closterium
littorale Unicelular 1
He2
Cyanobacteria Microcoleus
chthonoplastes Filamentoso 1
Chlorophyta
Closterium
ehrenbergii Unicelular 1
Closterium lineatum
Unicelular 2
He3 Chlorophyta Asterococcus
limneticus Colonial 1

96
Tabla 20. Continuación.
Zona de
muestreo Muestra División Especie
Nivel de
organización
Abundancia
(células/L;
colonias/L;
filamentos/L)
Espejo de agua
(dominado por Lemna sp. y
algunas otras
flotantes libres)
E1
(muestra
única)
Cyanobacteria Lyngbya lutea Filamentoso 500
Bacillariophyta
(diatomeas) Pinnularia sp. Unicelular 1
Chlorophyta
Asterococcus limneticus
Colonial 19
Micrasterias
sol Unicelular 2
Closterium ehrenbergii
Unicelular 6
Closterium
lineatum Unicelular 1
Spirogyra ternata
Filamentoso 100
Hymenachne
amplexicaulis
Hy1
Cyanobacteria Lyngbya lutea Filamentoso 100
Chlorophyta
Asterococcus limneticus
Colonial 16
Closterium
cornu Unicelular 1
Closterium
lineatum Unicelular 6
Closterium parvulum
Unicelular 1
Spirogyra
ternata Filamentoso 10
Hy2
Cyanobacteria Lyngbya lutea Filamentoso 100
Chlorophyta
Asterococcus
limneticus Colonial 86
Micrasterias
sol Unicelular 1
Closterium
ehrenbergii Unicelular 3
Closterium
lineatum Unicelular 2
Spirogyra
ternata Filamentoso 11

97
Tabla 20. Continuación.
Zona de
muestreo Muestra División Especie
Nivel de
organización
Abundancia
(células/L;
colonias/L;
filamentos/L)
Hymenachne
amplexicaulis
Hy2 Chlorophyta Volvox aureus Colonial
(cenobio) 1
Hy3
Cyanobacteria Lyngbya lutea Filamentoso 100
Chlorophyta
Asterococcus
limneticus Colonial 47
Closterium
cornu Unicelular 1
Closterium
ehrenbergii Unicelular 5
Closterium lineatum
Unicelular 3
Closterium
parvulum Unicelular 1
Spirogyra
ternata Filamentoso 9
Los índices de diversidad establecen que el ambiente más diverso es el parche de H.
marginata, la cual estuvo soportada en una equidad máxima, pese a su menor riqueza.
Los índices de dominancia fueron elevados en espejo y H. amplexicaulis debido a las
grandes abundancias de unas pocas especies (Tabla 21).
Tabla 21. Algunos índices de diversidad empleados para caracterizar la comunidad fitoplanctónica del humedal de El Clavo (enero 2009). Salida: PAST.
Índice Heliconia marginata Espejo de agua Hymenachne amplexicaulis
Riqueza (S) 5 8 9
Equidad (J´) 1,000 0,338 0,502
Shannon-Wiener (H´) 1,609 0,702 1,103
Dominancia de Simpson (D) 0,200 0,650 0,424
Berger-Parker (d) 0,200 0,790 0,556
El índice de similitud de Jaccard suministró información probabilística de los ambientes
a partir de su composición en especies. Las dos zonas con vegetación emergente
fueron más parecidas entre sí que respecto al espejo de agua (Tabla 22).
Tabla 22. Índice de similitud de Jaccard para las tres zonas estudiadas en el Clavo, Edo. Miranda (enero de 2009). El análisis se hizo en PAST.
Heliconia hirsuta Espejo de agua Hymenachne
amplexicaulis
Heliconia marginata 1 0,54054 0,64706
Espejo 0,54054 1 0,51429
Hymenachne amplexicaulis 0,64706 0,51429 1
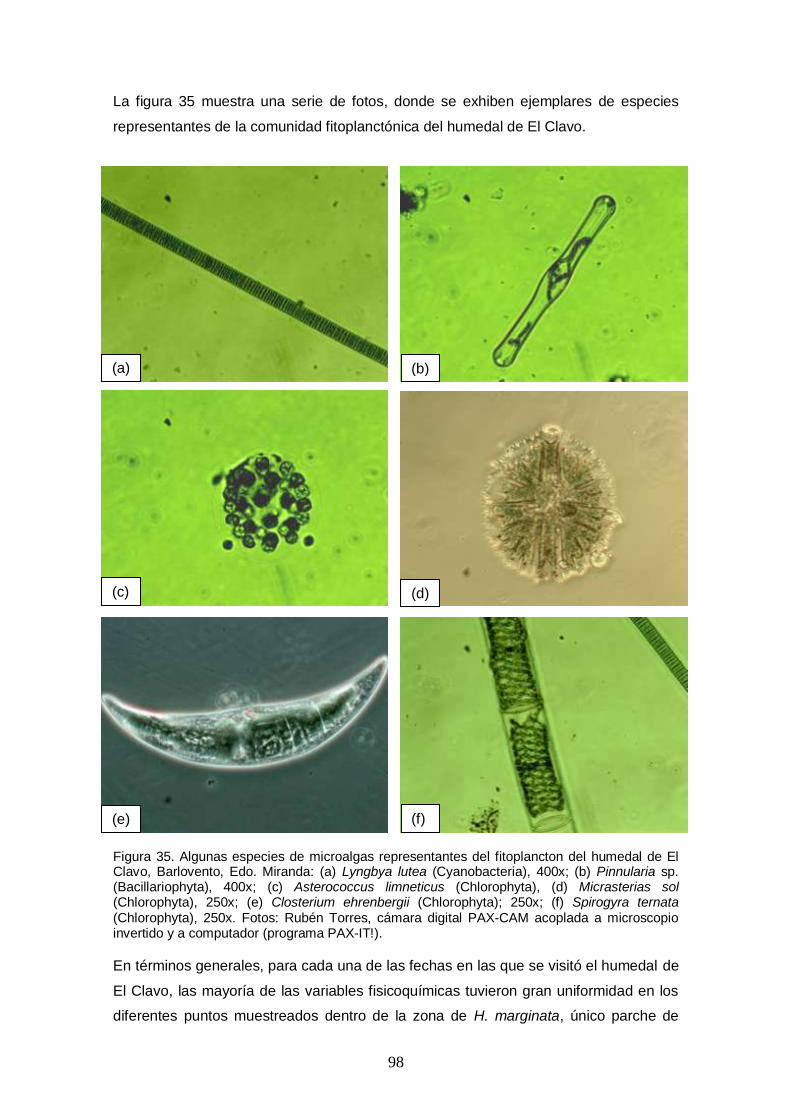
98
La figura 35 muestra una serie de fotos, donde se exhiben ejemplares de especies
representantes de la comunidad fitoplanctónica del humedal de El Clavo.
Figura 35. Algunas especies de microalgas representantes del fitoplancton del humedal de El Clavo, Barlovento, Edo. Miranda: (a) Lyngbya lutea (Cyanobacteria), 400x; (b) Pinnularia sp. (Bacillariophyta), 400x; (c) Asterococcus limneticus (Chlorophyta), (d) Micrasterias sol (Chlorophyta), 250x; (e) Closterium ehrenbergii (Chlorophyta); 250x; (f) Spirogyra ternata (Chlorophyta), 250x. Fotos: Rubén Torres, cámara digital PAX-CAM acoplada a microscopio invertido y a computador (programa PAX-IT!).
En términos generales, para cada una de las fechas en las que se visitó el humedal de
El Clavo, las mayoría de las variables fisicoquímicas tuvieron gran uniformidad en los
diferentes puntos muestreados dentro de la zona de H. marginata, único parche de
(a) (b)
(c) (d)
(e) (f)

99
vegetación donde se tomaron esas mediciones dentro del ámbito del estudio. La
profundidad de la lámina de agua disminuyó rápidamente de enero a febrero conforme
la estación seca se aproximó, señal de que la época fue una transición de lluvia a
sequía y los valores de oxígeno y conductividad variaron (Tabla 23).
Tabla 23. Valores medios desviaciones estándares de las variables fisicoquímicas tomadas en la zona de Heliconia marginata del humedal herbáceo de El Clavo, Edo. Miranda en enero - febrero de 2009.
Salida Profundidad
(cm)
Temperatura
(°C) pH %O2 [O2]mg/L CE (μS)
CE
específica
(μS/cm)
Salinidad
(‰)
05/01/2008 69,2 15,4 – 6,28 0,1 2,2 1,3 0,3 0,2 87,1 2,1 86,8 2,1 <<1
26/01/2008 13,7 7,0 24,7 0,6 5,92 0,2 8,7 6,4 0,7 0,5 108,36,1
107,25,8
0,1 0
30/01/2008 11,3 4,6 22,5 0,3 5,58 0,3 10,29,3
0,9 0,8 99,436,5
94,434,6
0,1 0
07/02/2008 12,7 6,4 24,7 0,9 6,41 0,1 6,5 1,9 0,5 0,1 104,08,4
102,86,9
0,1 0
14/02/2008 13,7 5,7 24,6 0,3 6,17 0,4 3,9 0,6 0,7 0,3 133,861,2
132,560,4
0,1 0
01/03/2008 11,3 4,6 23,1 0 5,73 0,2 10,83,2
1,0 0,4 154,461,5
149,457,9
0,1 0
Las mediciones de conductividad eléctrica (CE) y concentración de oxígeno (O2)
fluctuaron entre los puntos a lo largo del seguimiento. Dichas fluctuaciones
principalmente fueron notables en el oxígeno, pero ambas variables mostraron
tendencias al incremento de sus valores a lo largo del tiempo, conforme la profundidad
de la lámina de agua disminuyó (figura 36).
Figura 36. Variaciones temporales de (a) conductividad eléctrica y (b) oxígeno disuelto a medida que la lámina de agua disminuyó en el parche de Heliconia marginata (enero – febrero 2009).
(b)
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1 2 3 4 5 6
Tiempo (días)
[O2] m
g/L
0,0
10,0
20,0
30,0
40,0
50,0
60,0
70,0
80,0
Pro
f. (
cm
)
[O2]mg/L Profundidad (cm)
0,0
20,0
40,0
60,0
80,0
100,0
120,0
140,0
160,0
180,0
1 2 3 4 5 6
Tiempo (días)
CE
(μ
S)
0,0
10,0
20,0
30,0
40,0
50,0
60,0
70,0
80,0
Pro
f. (
cm
)
CE (μS) Profundidad (cm)
(a)

100
0
1
2
3
4
5
6
7
1,2,3 4,5 6
Muestra
Co
nc.
N (
mg
/L)
Los nutrientes determinados para el estudio en El Clavo fueron dos de los elementos
más importantes y limitantes para las plantas y el fitoplancton: el nitrógeno (N) y el
fósforo (P). La determinación de N total por método Kjeldahl (1883) resultó en una
concentración de 2,94 g/L de N para una muestra combinada 1-2-3, 6,44 g/L para otra
muestra combinada 4-5 y finalmente una concentración de 2,31 g/L para la muestra 6.
Llama la atención que en la muestra combinada 4-5 la concentración de N duplica a la
determinada en la muestra combinada 1-2-3 y casi triplica al contenido de la muestra
6, siendo todas muestras provenientes de la misma zona de H. marginata y tomadas
en la misma fecha. El punto de quiebre en la determinación de N lo determinó la
cantidad de ácido clorhídrico (HCl) empleado para cada titulación, ya que se necesitó
verter un volumen doble de HCl en la bureta para hacer el viraje a la coloración final de
la muestra combinada 4-5; la relación del volumen de HCl con la concentración de N
total es lineal o directa (figura 37).
Figura 37. Concentraciones de nitrógeno (N) en las muestras de agua colectadas en la zona de Heliconia marginata en la primera salida de campo al humedal de El Clavo (05/01/2009).
Las concentraciones de fósforo total (P) en fracciones de las mismas muestras
anteriormente mencionadas, se determinaron a partir de la curva de calibración
resultante de la medición de patrones de distintas concentraciones a las que se midió
su absorbancia a λ = 640 nm. En la muestra combinada 1-2-3 la concentración de P
fue 0,01 mg/L; en la muestra combinada 4-5 la concentración de P arrojó un resultado
físicamente ilógico, pues se trata de una concentración negativa (-4,61 mg/L), producto
de una transmitancia de 100% que al aplicar el factor de conversión a absorbancia
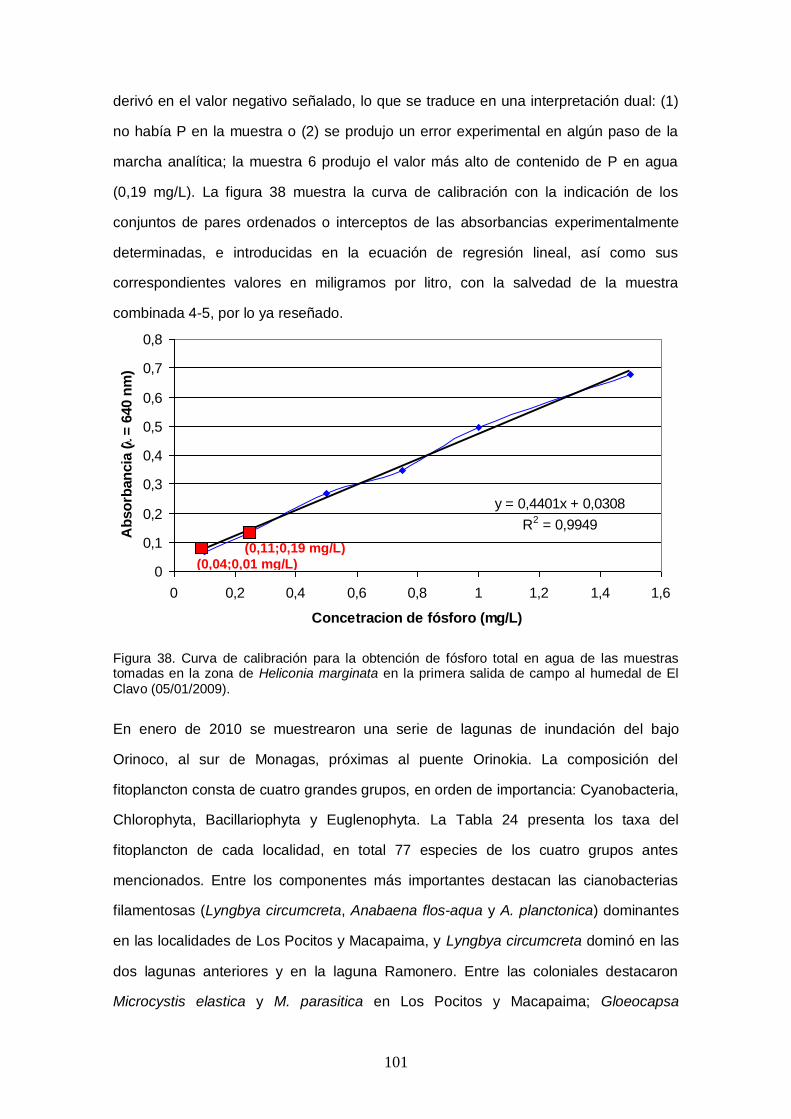
101
y = 0,4401x + 0,0308
R2 = 0,9949
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6
Concetracion de fósforo (mg/L)
Ab
so
rban
cia
(λ =
640 n
m)
derivó en el valor negativo señalado, lo que se traduce en una interpretación dual: (1)
no había P en la muestra o (2) se produjo un error experimental en algún paso de la
marcha analítica; la muestra 6 produjo el valor más alto de contenido de P en agua
(0,19 mg/L). La figura 38 muestra la curva de calibración con la indicación de los
conjuntos de pares ordenados o interceptos de las absorbancias experimentalmente
determinadas, e introducidas en la ecuación de regresión lineal, así como sus
correspondientes valores en miligramos por litro, con la salvedad de la muestra
combinada 4-5, por lo ya reseñado.
Figura 38. Curva de calibración para la obtención de fósforo total en agua de las muestras tomadas en la zona de Heliconia marginata en la primera salida de campo al humedal de El Clavo (05/01/2009).
En enero de 2010 se muestrearon una serie de lagunas de inundación del bajo
Orinoco, al sur de Monagas, próximas al puente Orinokia. La composición del
fitoplancton consta de cuatro grandes grupos, en orden de importancia: Cyanobacteria,
Chlorophyta, Bacillariophyta y Euglenophyta. La Tabla 24 presenta los taxa del
fitoplancton de cada localidad, en total 77 especies de los cuatro grupos antes
mencionados. Entre los componentes más importantes destacan las cianobacterias
filamentosas (Lyngbya circumcreta, Anabaena flos-aqua y A. planctonica) dominantes
en las localidades de Los Pocitos y Macapaima, y Lyngbya circumcreta dominó en las
dos lagunas anteriores y en la laguna Ramonero. Entre las coloniales destacaron
Microcystis elastica y M. parasitica en Los Pocitos y Macapaima; Gloeocapsa
(0,11;0,19 mg/L)
(0,04;0,01 mg/L)
102
aeruginosa fue importante en Macapaima. La cianobacteria unicelular Chroococcus sp.
destacó por su abundancia en Bañador. En Macapaima y Palital se encontraron
individuos de Spirulina subsalsa. De los grupos restantes, resaltan la diatomea
Achnanthes sp. en La Redonda, y la euglenofita Trachelomonas sp. en Guarampo. A
pesar de que ninguna especie de clorofita resultó numéricamente dominante, el grupo
fue el primero en número de especies y tercero en abundancia.
Tabla 24. Lista de especies y abundancia (cel./L) del fitoplancton en aguas libres de las lagunas estudiadas en el bajo Orinoco, sur de Monagas.
Grupos/Estaciones
Los
Pocit
os
Maca
paim
a
Pali
tal
Bañ
ad
or
La R
ed
on
da
Gu
aram
po
Las
Palo
meta
s
Ram
on
ero
Cyanobacteria
Unicelulares
Chroococcus sp. 886044
Filamentosas
Anabaena planctonica 1272000 1172640 439880 128822 56538 515560
A. flos-aqua 1600968 1549560 12564 40846 34052
A. spiroides 400000 78000 37704
Lyngbya sp. 4282
L. circumcreta 3000656 1900830 515288 2698000
L. limnetica 905628 161238 153958 15710 180084
Oscillatoria sp. 282690 15710 18852 96324
Spirulina subsalsa 52350 4188
Total filamentosas 7179252 5197308 21034 1203386 147674 15710 332946 3247612
Coloniales
Aphanocapsa biformis 234528 288972
A. pulchra 213588 188460 345821
Gloeocapsa aeruginosa 196836 586320 18846 97402 105090
Microcystis inserta 169614 41880
M. minima 200541
M. elastica 586320 339228 2094 223082
M. parasitica 335040 586320
M. robusta 196836 83760 14658 116254 3142 2094 4084 40787
Aphanocapsa biformis 234528 288972
A. pulchra 213588 188460 345821
Gloeocapsa aeruginosa 196836 586320 18846 97402 105090

103
Tabla 24. Continuación.
Grupos/Estaciones
Los
Pocit
os
Maca
paim
a
Pali
tal
Bañ
ad
or
La R
ed
on
da
Gu
aram
po
Las
Palo
meta
s
Ram
on
ero
A. pulchra 213588 188460 345821
G. aeruginosa 196836 586320 18846 97402 105090
Microcystis inserta 169614 41880
M. minima 200541
M. elastica 586320 339228 2094 223082
M. parasitica 335040 586320
M. robusta 196836 83760 14658 116254 3142 2094 4084 40787
Merismopedia sp. 12564 12568
M. punctata 10470 282690 3142 3142 35598
Nostoc sp. 113076 2094
Total coloniales 2056308 2399724 48162 452448 6284 2094 39682 692239
Total Cianobacterias 9235560 7597032 69196 1655834 153958 17804 372628 3939851
Chlorophyta
Actinastrum
hantzschii
15710
Artrodesmus sp. 33504
Bambusina sp. 20940
Closterium sp. 10470 41880
C. acutum 3142
C. aplanatum 197946
C. cetaceum 20940
C. kützingii 15710
C. ehrenbergii 4188
C. macilentum 113076 9426
Coelastrum cambricum
37692 12564
Cosmarium sp. 4188
C. bioculatum 3142
C. denticulatum 6284
C. logiense 2094
C. margaritiferum 6284
Euastrum ansatum 2094
E. elegans 2094
E. pulchellum 16752
Gonatozygon sp. 12568
104
Tabla 24. Continuación.
Grupos/Estaciones
Los
Pocit
os
Maca
paim
a
Pali
tal
Bañ
ad
or
La R
ed
on
da
Gu
aram
po
Las
Palo
meta
s
Ram
on
ero
G. pilosum 23034
Kirchneriella lunaris 2094 37692
Pediastrum duplex 41880
P. tetras 2094 9426 6284 25128
Scenedesmus sp. 167520 12564 9426
S. armatus 2094 6284
S. bimorfus 6284
S. brasiliensis 41880 16752
S. intermedius
S. quadricauda 73290 50272
Selenastrum gracile 4188
Spirogyra sp. 12564 3142
S. gracile 18846 6284 3142
Staurastrum sp. 31410 12568
S. brasiliense 10470
S. leptocladum 3142
S. noticum 3142 3142
S. tetracerum 2094
Staurodesmus sp. 8376
Tetraedrum
arthrodesmiformis
14658
T. minimum 6284
Ulothrix zonata 4188
Xanthidium sp. 2094
Total Chlorophyta 0 544440 211494 122538 270212 50258 77478 0
Bacillariophyta
Achnanthes sp. 249186 97402 628400 297348
A. lanceolata 3142
A. heteromorpha 12568
Actinella guianensis 3142
Gomphonema sp. 4188 12568
G. tenellum 12564
Hydrosera sp. 4188
Melosira granulata 10470 226152 21994 6284 50272 108888 138943
Pinnularia sp. 8376 3142 9426

105
Tabla 24. Continuación.
Grupos/Estaciones
Los
Pocit
os
Maca
paim
a
Pali
tal
Bañ
ad
or
La R
ed
on
da
Gu
aram
po
Las
Palo
meta
s
Ram
on
ero
P. dactylus 4188
P. hemiptera 12564
P. pectinata 9426
P. singularis 33504 8376
Tryblionella rustica 2094 3142 230340
Total
Bacillariophyta
10470 226152 330852 122538 678672 59698 644952 138943
Euglenophyta
Trachelomonas sp. 2094 593838 60726
Total Euglenophyta 2094 593838 60726
Total Fitoplancton 9246030 1449048 1553748 4141156 3010036 4739076 3815268 4257734
Los índices de diversidad señalan que las lagunas con mayor variedad fueron
Macapaima y Palital, puesto que tuvieron las mayores riquezas y equidades, junto con
Los Pocitos, el tercer ambiente más diverso y que presentó la misma equidad que
Macapaima. En las lagunas donde la equidad disminuyó, los índices de dominancia
aumentaron, además las riquezas también resultaron bajas en los mismos, lo que trajo
como consecuencia valores de diversidad relativamente bajos (Tabla 25).
Tabla 25. Algunos índices de diversidad empleados para caracterizar la comunidad fitoplanctónica de lagunas de inundación del bajo Orinoco (sur de Edo. Monagas), para enero de 2010. Salida: PAST.
Índice Los
Pocitos Macapaima Palital Bañador
La
Redonda Guarampo
Las
Palometas Ramonero
Riqueza (S) 24 35 43 29 25 9 14 14
Equidad (J´) 0,82 0,82 0,75 0,73 0,48 0,34 0,80 0,68
Shannon y Wiener
(H´) 2,59 2,92 2,82 2,45 1,54 0,74 2,12 1,80
Dominancia de
Simpson (D) 0,12 0,08 0,15 0,13 0,37 0,68 0,15 0,29
Berger-Parker (d) 0,26 0,17 0,37 0,27 0,57 0,82 0,26 0,52
Por su parte, el índice de similitud de Jaccard revela que los ambientes de mayor
similitud en el número y abundancias de especies comunes fueron Macapaima y Los
Pocitos. Los otros ambientes mostraron pocas similitudes entre sí, por lo que cada una
de esas lagunas tuvo una composición de especies diferente (Tabla 26).
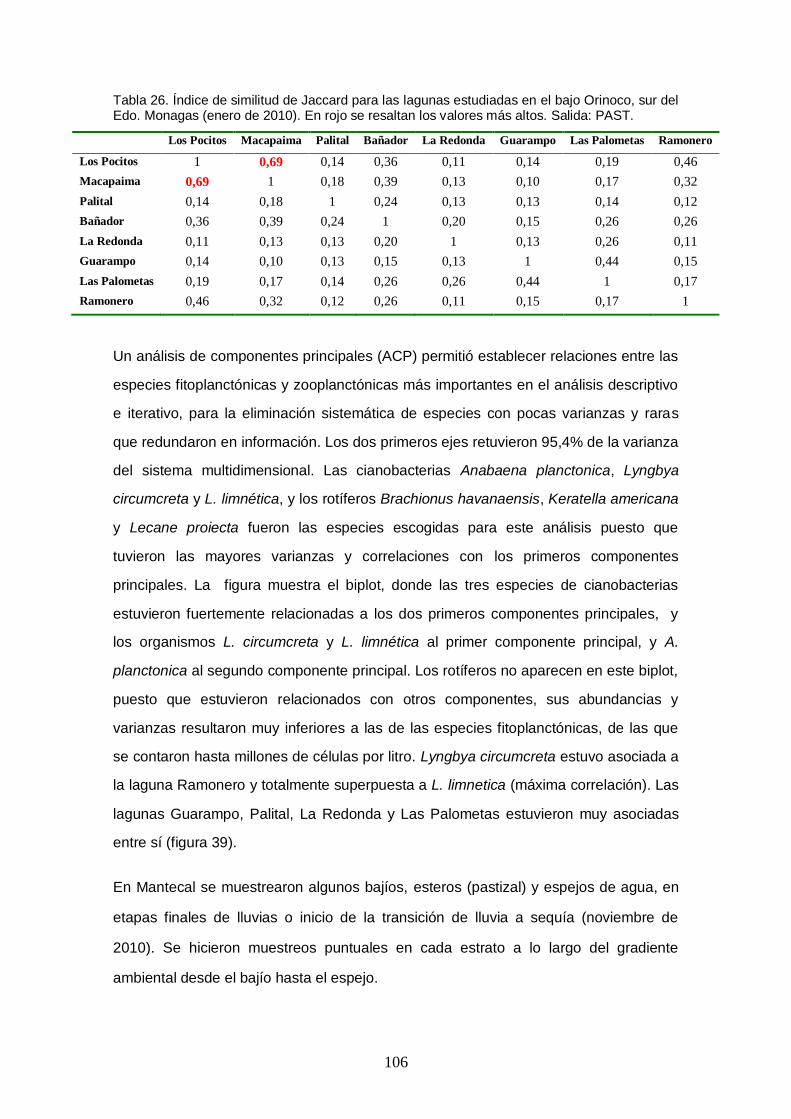
106
Tabla 26. Índice de similitud de Jaccard para las lagunas estudiadas en el bajo Orinoco, sur del Edo. Monagas (enero de 2010). En rojo se resaltan los valores más altos. Salida: PAST.
Los Pocitos Macapaima Palital Bañador La Redonda Guarampo Las Palometas Ramonero
Los Pocitos 1 0,69 0,14 0,36 0,11 0,14 0,19 0,46
Macapaima 0,69 1 0,18 0,39 0,13 0,10 0,17 0,32
Palital 0,14 0,18 1 0,24 0,13 0,13 0,14 0,12
Bañador 0,36 0,39 0,24 1 0,20 0,15 0,26 0,26
La Redonda 0,11 0,13 0,13 0,20 1 0,13 0,26 0,11
Guarampo 0,14 0,10 0,13 0,15 0,13 1 0,44 0,15
Las Palometas 0,19 0,17 0,14 0,26 0,26 0,44 1 0,17
Ramonero 0,46 0,32 0,12 0,26 0,11 0,15 0,17 1
Un análisis de componentes principales (ACP) permitió establecer relaciones entre las
especies fitoplanctónicas y zooplanctónicas más importantes en el análisis descriptivo
e iterativo, para la eliminación sistemática de especies con pocas varianzas y raras
que redundaron en información. Los dos primeros ejes retuvieron 95,4% de la varianza
del sistema multidimensional. Las cianobacterias Anabaena planctonica, Lyngbya
circumcreta y L. limnética, y los rotíferos Brachionus havanaensis, Keratella americana
y Lecane proiecta fueron las especies escogidas para este análisis puesto que
tuvieron las mayores varianzas y correlaciones con los primeros componentes
principales. La figura muestra el biplot, donde las tres especies de cianobacterias
estuvieron fuertemente relacionadas a los dos primeros componentes principales, y
los organismos L. circumcreta y L. limnética al primer componente principal, y A.
planctonica al segundo componente principal. Los rotíferos no aparecen en este biplot,
puesto que estuvieron relacionados con otros componentes, sus abundancias y
varianzas resultaron muy inferiores a las de las especies fitoplanctónicas, de las que
se contaron hasta millones de células por litro. Lyngbya circumcreta estuvo asociada a
la laguna Ramonero y totalmente superpuesta a L. limnetica (máxima correlación). Las
lagunas Guarampo, Palital, La Redonda y Las Palometas estuvieron muy asociadas
entre sí (figura 39).
En Mantecal se muestrearon algunos bajíos, esteros (pastizal) y espejos de agua, en
etapas finales de lluvias o inicio de la transición de lluvia a sequía (noviembre de
2010). Se hicieron muestreos puntuales en cada estrato a lo largo del gradiente
ambiental desde el bajío hasta el espejo.
107
Figura 39. Biplot de los dos primeros componentes principales del ACP para los ambientes lagunares del Orinoco bajo, sur de Monagas. Los dos primeros componentes principales retuvieron 95,4% de la inercia total del sistema multidimensional original. Salida: MVSP 3.0.
La Tabla 27 muestra la composición de especies fitoplanctónicas en tres ambientes de
los módulos inundables de Mantecal. Dominaron las clorofitas, seguidas de las
cianobacterias y en menores proporciones las euglenofitas y diatomeas. Las especies
más abundantes fueron las clorofitas Desmidium baylei y Scenedesmus brasiliensis en
bajío, Euastrum pectinatum en bajío y pastizal y Cosmarium subtumidum en pastizal.
La cianobacteria Microcystis aeruginosa fue el principal representante de este grupo
en las muestras, y también se hallaron filamentos de Arthrospira sp. El espejo de agua
fue el ambiente menos rico en especies y también exhibió las menores densidades
poblacionales.
Tabla 27. Composición y abundancia (células por litro) de la comunidad fitoplanctónica de tres ambientes de los módulos inundables de Mantecal (noviembre 2010).
Taxa Bajío Pastizal Espejo
Cyanobacteria
Anabaena planctonica 5416 745 45
Arthrospira sp. 1285 4545
Chroococcus sp. 2569 458 89
CP 2
CP 1
Los PocitosMacapaima
Palital
Bañador
La Redonda
GuarampoLas Palometas
Ramonero
-193789.5
-387578.9
193789.5
387578.9
581368.4
775157.9
968947.3
-193789.5-387578.9 193789.5 387578.9 581368.4 775157.9 968947.3
Anabaena planctonica
Lyngbya circumcretaLecane proiectaBrachionus havanaensisKeratella americanaLyngbya limnetica
Vector scaling: 807456,00

108
Tabla 27. Continuación.
Cylindrospermum muscicola 1526
Lyngbya circumcreta 1045
Merismopedia glauca 748 1256
Microcystis aeruginosa 14025 225
M. flos-aqua 4188
Nostoc sp. 458 236
Oscillatoria tenuis 4326 4589
Chlorophyta
Cosmarium sp. 457 2546
Cosmarium subtumidum 4458 14565
Closterium ehrenbergii 784 6821
Desmidium baylei 45126 4895
Euastrum crassum 1546 74
Euastrum pectinatum 12546 15878
Hyalotheca sp. 3012 485 152
Micrasterias sol 1458 5963 58
Scenedesmus brasiliensis 15689 856
Scenedesmus quadricauda 4589 589
Staurastrum paradoxum 2589
Staurastrum triangularis 451 5846
Euglenophyta
Trachelomonas superba 745 1265 2548
Phacus orbicularis 512
Bacillariophyta
Achnanthes lanceolata 452
Gomphonema sp. 74 859 412
Una observación de los índices de diversidad permite fácilmente advertir que en el
pastizal se conjugan una riqueza y equidad elevadas que se combinan para la
obtención de un valor de diversidad Shannon-Wiener relativamente alto, si se compara
con los otros ambientes contiguos. El bajío fue el segundo lugar más diverso, donde
riqueza y equidad disminuyeron un poco. El espejo fue menos rico y equitativo que los
otros dos ambientes, y los índices de dominancia, especialmente el de Berger-Parker,
subieron en forma notable (Tabla 28).
Tabla 28. Algunos índices de diversidad empleados para caracterizar la comunidad fitoplanctónica entres ambientes de los módulos inundables de Mantecal (Edo. Apure), en noviembre de 2010. Salida: PAST.
Índice Bajío Pastizal Espejo
Riqueza (S) 19 22 8
Equidad (J´) 0,744 0,803 0,611
Shannon y Wiener (H´) 2,189 2,483 1,271
Dominancia de Simpson (D) 0,181 0,114 0,387
Berger-Parker (d) 0,365 0,208 0,553

109
El índice de Jaccard muestra que el bajío y el pastizal tuvieron una gran similitud, y
que el espejo fue un ambiente totalmente diferente a ambos (Tabla 29).
Tabla 29. Índice de similitud de Jaccard para comparación de los tres ambientes estudiados en Mantecal (Edo. Apure). Salida: PAST.
Bajío Pastizal Espejo
Bajío 1 0,640 0,285
Pastizal 0,640 1 0,304
Espejo 0,286 0,304 1
Fosas petroleras y lagunas con emanaciones naturales de crudos. La figura 40
muestra un conjunto de fotos con observaciones de especies de microalgas de una
muestra de pozo (P57X) del Edo. Bolívar, 2009. La muestra proviene en forma
específica de una pequeña charca fangosa en la parte exterior del pozo mencionado,
enmarcada en un lugar donde se encuentra petróleo, ripios y sedimentos de
perforación. Se evidenciaron muchas cianobacterias filamentosas (principalmente
Oscillatoria tenuis); también hubo una cantidad de euglenofitas y diatomeas pennadas,
y algunas clorofitas (principalmente Scenedesmus spp.).
Figura 40. Imágenes que muestran la composición de especies fitoplanctónicas de una charca fangosa, específicamente proveniente del borde exterior de un área de pozos petroleros en el norte del Edo. Bolívar, 2009.

110
Figura 40. Continuación.

111
Figura 40. Continuación.

112
Figura 40. Continuación.
113
Figura 40. Continuación.
La Fosa El Caracol en la región de Caico Seco, ubicada en el municipio Aragua del
estado Anzoátegui, fue escogida para la búsqueda de especies de microalgas nativas
con gran potencialidad para la producción de biodiesel, por representar un área de
exposición histórica a hidrocarburos que operó durante muchos años como una de las
principales y más grandes recolectoras de desechos petrolíferos de la zona. Se
tomaron muestras de agua libre en tres puntos denominados: Zona I, Zona II y Zona
III.
En la Zona I se identificaron dos géneros de cianobacterias (Chroococcus y
Anabaena), un género de clorofita (Chlamydomonas) y una especie de diatomea
(Tabellaria flocculosa). Para la Zona II se reportaron dos géneros de clorofitas
(Chlamydomonas y Chlorella), un género de cianobacteria (Anabaena, aunque una
especie diferente a la encontrada en la Zona I) y uno de diatomea (Navicula).
Finalmente en la Zona III, a diferencia del resto de las zonas, en esta localidad de
estudio se identificaron cinco géneros de cianobacterias (Lyngbya, Merismopedia,
Aphanocapsa, Synechocystis y Chroococcus) además de una clorofita (Chlorella sp.).
Salinas y lagunas hipersalinas. La laguna de Boca Chica en la isla de Margarita,
Edo. Nueva Esparta, es un ambiente natural extremófilo, con una salinidad elevada (>
40 ppm), que se presenta como una laguna costera en la costa suroeste de la

114
península de Macanao, donde ocurren emanaciones naturales de crudo. Un análisis
de muestras tomadas en este lugar, mayo de 2009, refiere que en las mismas se
encontraron ejemplares de Dunaliella primolecta (Chlorophyta) y Chaetoceros sp.
(Bacillariophyta) en densidades muy bajas, 125 y 145 células por litro,
respectivamente.
La salina de Las Cumaraguas, norte de la península de Paraguaná, Edo. Falcón, es
otro ambiente extremófilo donde se ubica una de las poblaciones naturales de
Dunaliella salina más importantes del Caribe sur. Las muestras colectadas exhibieron
abundancias de 14.000 a 75.000 células por litro, en fase de crecimiento, y se inició el
cultivo de la especie en la Cámara de Crecimiento con la metodología expuesta y
resultados explicados en la sección de cultivos.
Ambientes marinos costeros. En Mochima y Puerto Cabello, entre los años 2009 y
2010, se colectaron muestras de fitoplancton marino donde las diatomeas centrales
(Bacillariophyta) y dinoflagelados dominaron en riqueza y abundancia los ambientes.
En la bahía de Mochima, se encontraron individuos de las diatomea Chaetoceros sp. y
Nitzschia valens, así como la haptofita Isochrysis galbana, en densidades bajas (100 a
250 células por litro). En el muelle de Puerto Cabello se hallaron blooms (o
afloramientos) de la diatomea Coscinodiscus sp. (>17.000.000 de células por litro), el
dinoflagelado colonial Alexandrium catenella (>15.000.000 de células por litro) y la
cianobacteria Anabaena sp. (>5.000.000 de células por litro).
Síntesis de los ambientes. La figura 41 muestra una síntesis gráfica de todas las
regiones muestreadas y las composiciones porcentuales del fitoplancton en cada
lugar. Se muestra que en los ambientes de aguas continentales las cianobacterias,
diatomeas y clorofitas fueron los grupos más representativos. Las euglenofitas
constituyeron otros representantes de agua dulce encontrados en porcentajes muy
inferiores, pero en la fosa petrolera de Bolívar se evidencia una gran importancia
numérica de este taxón, junto a cianobacterias, clorofitas y diatomeas en porcentajes
menores. Las clorofitas del género Dunaliella resultaron muy importantes en
ambientes hipersalinos, con una dominancia absoluta en las salinas de Las
Cumaraguas. En medios marítimos, las diatomeas, pirrofitas (o dinoflagelados) y las
haptofitas, éstas en Mochima representadas por Isochrysis galbana, resultaron los
taxones predominantes. Con esta representación gráfica de las divisiones de
cianobacterias y microalgas eucariotas, se comprueban las grandes variaciones
porcentuales de los taxones mayores del fitoplancton, como consecuencia de la
variedad de ambientes acuáticos representados en el país.

115
Figura 41. Composiciones porcentuales de las divisiones de organismos procariotas y eucariotas integrantes de la comunidad del fitoplancton en las regiones estudiadas en el país a lo largo del estudio.
0%10%20%30%40%50%60%70%80%90%
100%
Co
mp
osi
ció
n p
orc
en
tual
Haptofitas
Pirrofitas
Euglenofitas
Diatomeas
Clorofitas
Cianobacterias

116
8. Discusión
8.1 Cultivos
Los medios preparados para mantenimiento y experimentos se caracterizaron por sus
componentes exclusivamente minerales (medios minerales), es decir, sólo se
emplearon sales inorgánicas sin la incorporación de fuentes orgánicas para los
macronutrientes (Ej.: urea como fuente de nitrógeno) y micronutrientes. La única
incorporación orgánica a los medios minerales fue el EDTA disódico, el cual no
funcionó como nutriente, sino como agente quelante que “secuestró” iones Hierro II
(Fe2+) y calcio (Ca2+). Por este mecanismo ambos cationes permanecieron como
formas solubles en agua. En el caso específico del Fe2+, la acción del quelante evitó
que se oxidara a Hierro III (Fe3+) y reaccionar con el fósforo del medio, para formar un
precipitado de fosfato férrico, lo que trae como consecuencia la no disponibilidad de
este nutriente para las microalgas. De forma homóloga, la mediación del quelante
impidió que el ion calcio hiciese precipitar a los aniones fosfato y sulfato.
Por otro lado, no hubo necesidad de suplementar con vitaminas y carbohidratos
(cultivos mixotróficos) a ningún medio nutritivo, a pesar de formar parte de sus
formulaciones. Ciferri (1983) y Duerr y col. (1997) mencionan que cultivos mixotróficos
producen una cantidad de biomasa elevada en comparación con la autotrofía (medios
minerales para microalgas, incluidas fuentes inorgánicas de carbono, como los
empleados en esta investigación) y heterotrofía. Dicha aseveración se puede deber al
efecto energético de la incorporación de carbohidratos a los medios, así como la
actuación eficiente de las vitaminas como coenzimas en muchos procesos catabólicos.
Además, estos aditivos orgánicos aumentan el valor nutricional de la biomasa de
microalgas. No obstante, adiciones de carbohidratos simples como glucosa, sacarosa
o fructosa, también producen crecimientos rápidos y descontrolados de bacterias, las
cuales pueden ser patógenas y mortales para los cultivos. Otros microorganismos
favorecidos por la adición de carbohidratos y vitaminas son las microalgas invasoras,
las cuales suelen poseer tasas de crecimiento per cápita (r) superiores a las de
especies de valor biotecnológico, como Arthrospira spp. y Spirulina spp., las que
pueden ser desplazadas por competencia. El medio altamente básico (pH>9,5) donde
crece A. platensis es un escudo o primera línea de defensa para estas especies
primitivas, las cuales evolucionaron en esos ambientes agrestes e inhóspitos para casi
cualquier otra forma de vida (Ciferri 1983).
Para el medio Schlösser (1994, modificación del medio Aiba y Ogawa (1977)), un
aporte importante realizado por Parra (2005) fue la sustitución en algunas sales, del

117
catión sodio (Na+) por el catión potasio (K+), para generar una relación Na+ : K+ 4 a 1
(la proporción en el medio original es de 10 a 1). Este ajuste tuvo por finalidad una
mayor aproximación a las condiciones imperantes en los medios naturales de A.
platensis en los lagos carbonatados de África (L’évêque 1987).
La razón de la adaptabilidad de la cepa Lefevre de A. platensis a las condiciones
naturales del país puede derivar de las características del lugar de origen de esa
variedad, África tropical (específicamente borde sur del desierto del Sahara). Las
similitudes climáticas en temperatura y radiación solar altas durante la mayor parte del
año son adecuadas para esa cepa silvestre, más que para otras que sólo se han
cultivado en invernaderos y condiciones controladas de manera persistente. Los
ambientes naturales de A. platensis son lagos y lagunas carbonatados, cerrados,
altamente alcalinos con valores de temperatura y radiación solar elevados. Estos
cuerpos de agua están confinados a regiones desérticas tropicales y subtropicales en
el interior de los continentes y, en general, son ambientes naturales muy hostiles, por
lo que a menudo los centros urbanos están apartados de ellos (Jones y Grant 1999).
La composición de los medios de cultivo afecta la tasa de crecimiento y producción de
biomasa de los microorganismos, y define el costo de producción del mismo (Raoof y
col. 2006). Por esa razón, para esta investigación fue de gran importancia obtener
medios de cultivo óptimos que permitiesen maximizar la producción y minimizar los
costos. Por ejemplo, el empleo de sales grado técnico y alimenticio, en lugar de sales
grado reactivo o analítico de precios muy elevados debido a sus altos grados de
pureza, ha permitido la viabilidad económica de los experimentos. Por otro lado, en
numerosas oportunidades sales de alta pureza han demostrado ser menos nutritivas
que las de grado técnico o alimenticio. Estas impurezas constituyen valores nutritivos
agregados que aumentan la calidad nutricional de los medios preparados.
8.2 Ensayos con poblaciones de Arthrospira platensis
Los ensayos factoriales 22 permitieron llegar a una primera aproximación del proceso
de optimización de los medios de cultivos, con el propósito firme de conocer y procurar
tasas de crecimientos poblacionales y rendimientos de biomasa óptimos para la meta
de aprovechamiento biotecnológico de A. platensis. Los medios 1 y 2 presentaron una
dinámica similar, lo cual indica que el intervalo de variación de las concentraciones de
Na+ y K+ no afectaron la tasa de crecimiento poblacional, por lo que puede explorarse
la posibilidad de disminuir las concentraciones de estos iones hasta un mínimo que
permita mantener esta tasa de crecimiento. La tasa de crecimiento ligeramente menor
del medio 3 indica que un incremento del Cl- pudo afectar en forma desfavorable el

118
crecimiento de A. platensis. En el caso de la curva de crecimiento del medio 4, con
una fase de latencia y adecuación muy prolongada, la misma señala que una
disminución pronunciada del Cl- en conjunto con el SO42- incrementan de modo
apreciable dicha adaptación al medio. Sin embargo, la baja concentración de estos
iones no afecta la tasa de crecimiento una vez alcanzada la fase exponencial.
En los ensayos se lograron ajustes del pH del medio de cultivo a un valor de 10,5, una
unidad por encima de lo establecido por Melack (1979), lo que significó una mejor
aproximación al pH del medio natural de A. platensis. Los cambios incorporados por
Parra (2005), y reajustes posteriores en el segundo ensayo factorial 22, determinaron
que el pH del medio se mantuviera estable por un tiempo prolongado y con pocas
variaciones en la conductividad, de este modo siendo alcanzadas las condiciones
óptimas para el crecimiento de A. platensis. Estas condiciones de laboratorio se
corresponden con los trabajos de Wood y Talling (1988), Njuguna (1988), Melack
(1988) y Jones y Grant (1999), donde se reportan alcalinidades extremas y
osmolaridades elevadas de los ambientes originarios de esta cepa.
A un pH de 8,5, el medio de cultivo de A. platensis frecuentemente se contaminó con
otras especies de microalgas (principalmente de los géneros de cianobacterias
Anabaena, Microcystis y Oscillatoria), las cuales se encuentran comúnmente en el
agua potable, lo que visualmente originó matices de color verde diferentes al verde-
azul, característico de los medios observados cuando A. platensis es la única especie
existente. Sena y col. (2011) mencionan un tratamiento de alta alcalinidad (pH 12, con
empleo de KOH) de 72 h, como procedimiento inicial determinante para eliminar a
Microcystis spp. y otras cianobacterias, partiendo del hecho de que A. platensis
sobrevive a condiciones de dureza y alcalinidad extremas en lagos de soda (en inglés
“soda lake”). La toxicidad de un medio muy alcalino con contenido de sales potásicas
como KOH o K2CO3 para cianobacterias invasoras u oportunistas como Microcystis
aeruginosa, aparentemente viene del hecho de que esta especie y congéneres tienen
densidades bajas de canales de potasio, por lo que una entrada masiva de iones
potasio (K+) al compartimento intracelular no permite su salida rápida por los canales,
debido a su insuficiencia numérica, y ocurre un gran desequilibrio electroquímico con
inversión de la polaridad de la membrana plasmática. La consecuencia final es la
muerte de la célula por intoxicación con potasio.
Para la cuantificación del peso neto de la biomasa de A. platensis, resultó necesario
determinar una relación lineal entre la absorbancia y el peso seco de la biomasa, de tal
manera de conocer su peso neto en cualquier instante de tiempo de manera rápida,
precisa y no destructiva. Boussiba y Richmond (1979) fueron los únicos en cuantificar

119
biomasa seca de microalgas en cultivos controlados, pero no lo hicieron en sentido
estricto, ni mucho menos en una relación lineal para su determinación. Otros autores
como Chen y col. (1996), Chen y Zhang (1997) y Zhang y Chen (1999), no
desarrollaron una metodología cuantitativa para estimar la biomasa, ya que ésta fue
tomada húmeda, lo que puede conducir a errores importantes en la cuantificación de
su peso.
La tasa de crecimiento para un cultivo autotrófico de A. platensis en medio Zarrouk,
obtenida por Chen y Zhang (1997), fue de 0,0083 h-1, mientras que para un cultivo
mixotrófico (suplementado con glucosa) en medio Zarrouk, reportado por los mismos
autores, el valor fue de 0,026 h-1, ambos en condiciones de laboratorio. La tasa de
crecimiento per cápita o de recambio obtenida en este trabajo para el cultivo
autotrófico en medio 1, el más productivo de las cuatro combinaciones obtenidas para
condiciones controladas, fue de 0,50 días-1 = 0,021 h-1, similar a la obtenida por Chen
y Zhang (1997) para el cultivo mixotrófico, y más de dos veces superior a la
conseguida para su cultivo autotrófico. Entonces, para los fines de este trabajo resulta
significativo el rendimiento notablemente superior obtenido en cultivos con medios
minerales, y similar al mixotrófico sin suplir glucosa u otra fuente orgánica.
El ensayo factorial fraccionado 26-1 permitió evaluar el crecimiento diferencial de
poblaciones de A. platensis en un gran número de medios de cultivos simultáneos con
combinaciones aleatorias de macronutrientes. El crecimiento diferenciado para cada
tratamiento puede deberse a que estas cianobacterias crecen en condiciones
altamente selectivas, principalmente en concentraciones altas de sales (Borowitzka
1999). Es evidente que cultivos con tratamientos centrales y algunos con
concentraciones importantes de ciertas sales inorgánicas propiciaron crecimientos
vigorosos y relativamente saludables de las poblaciones de A. platensis, con poca o
nula contaminación de otras especies de microalgas. En contraste, otros tratamientos
fueron poco viables por carencias considerables de una buena parte de los
requerimientos nutricionales de la microalga, disminuyendo notablemente la capacidad
de carga del medio. No es descartable que debido al empleo de sales amónicas en los
tratamientos, sus concentraciones hayan sido suficientemente altas en algunos
recipientes, como para intoxicar o limitar el crecimiento de las poblaciones residentes
de A. platensis, conocido el efecto deletéreo que el amonio en concentraciones altas
tiene sobre esta especie (Boussiba 1989).
Como consecuencia de la selectividad de medios de cultivo de A. platensis, la especie
contó con dinámicas de crecimiento poblacional variables. Vonshak y Richmond
(1985) reportaron que tasas de crecimiento per cápita elevadas se observaron a

120
densidades poblacionales bajas, mientras que tasas de crecimiento bajas estuvieron
relacionadas con limitación por luz en cultivos densos bajo condiciones óptimas a cielo
abierto, lo que coincide con la mayoría de las curvas obtenidas para el ensayo
multifactorial a partir del punto de inflexión, cuyas dinámicas se maximizaron en
medios centrales, como el tratamiento 36, que contaron con las concentraciones
óptimas de los principales nutrientes, en especial las fuentes de carbono.
El tiempo de duplicación (tg) es un parámetro poblacional importante en la
caracterización de la dinámica de crecimiento poblacional de microorganismos, y tiene
gran aplicabilidad en biotecnología como medida de eficiencia de cultivos masivos de
microalgas para maximizar la velocidad y rendimiento de los mismos. Melack (1979)
obtuvo para A. platensis un tiempo de recambio (“turnover time”), comparable con el tg
obtenido en varios de medios del ensayo factorial fraccionado, de 8,9 a 18,9 h en
condiciones naturales en un lago de Kenia. Para especies congéneres de A. platensis
o próximas a ella, se tiene que Tomaselli y col. (1995) determinaron
experimentalmente valores de tg de 27 horas para Spirulina subsalsa en condiciones
de laboratorio; Garnier y col. (1994) obtuvieron un tg de 70 h para A. maxima en
condiciones de laboratorio.
Los cultivos a mayor escala (tanques circulares de 500 L y cultivadores tipo carrusel
de 2000 L) brindaron la oportunidad de probar el medio optimizado en botellones, para
iniciar el cultivo masivo de A. platensis. Es importante resaltar la disposición espacial
de los tanques para el experimento. Debido a que para el momento del ensayo en
tanques se aproximaba el solsticio de invierno, la proyección de la eclíptica solar sobre
el Ficotrón ocurrió a más de 30° al sur, más allá de los 10° respecto al Ecuador, lo cual
constituye una fuente de variación adicional “no deseada”. Para contrarrestar este
efecto, se escogió un “Diseño en Bloques Aleatorios” que permitió extraer la variación
“posible”, producto del fenómeno descrito. El análisis de varianza no produjo
diferencias significativas entre los bloques, lo que hace presumir que la inclinación u
oblicuidad de los rayos solares no tuvo una implicación directa en la dinámica de los
cultivos. En otros términos, la diferencia de minutos transcurridos entre los tanques
que se iluminaron primero en el eje cardinal Este, con los rayos iniciales del alba, y los
últimos tanques en el punto cardinal opuesto iluminados de último, no repercutió de
manera decisiva en el crecimiento de las poblaciones, a pesar de que los
microorganismos responden de manera rápida a cambios igualmente rápidos. La
profundidad de la lámina sí tuvo un efecto significativo, esto debido a que a mayor
profundidad los rayos solares penetran cada vez con menor intensidad y se forman

121
puntos oscuros donde las microalgas no pueden fotosintetizar y mueren, cayendo en
forma notable la productividad primaria.
También es destacable la disminución del rendimiento de la cosecha de biomasa en
cultivos a escalas mayores a las de botellones. En estos recipientes la producción de
biomasa fue alta en buena medida por el volumen de los mismos, pero también a su
disposición inclinada que aumentó en forma notable la superficie de exposición a los
rayos solares. La temperatura es otro factor decisivo en la producción primaria, la
misma en botellones fue 2 o 3ºC superior a los tanques, los cuales tuvieron en contra
su diseño, pues son recipientes muy altos que generan sombra abundante aún en
horas del mediodía y el material (polietileno) no es conductor de calor. En estanques
tipo carrusel el volumen empleado es de 2000 L, la capacidad calórica aumenta aún
más, no obstante sus temperaturas fueron ligeramente más altas que las de los
tanques, ya que este tipo de cultivadores tiene un diseño de superficie amplia con
poca profundidad (lámina delgada), lo que genera mayor exposición a los rayos
solares y generación de pocos puntos muertos o zonas no iluminadas. No obstante, la
producción de biomasa no fue elevada, quizá debido a algún factor físico como la
temperatura, para la cual hubo un seguimiento y se registraron valores moderados a
bajos (18 - 20°C en horas diurnas; 17 - 19°C durante las noches). La causa alternativa
puede obedecer a la preparación y suministro de medio nutritivo en cantidades
suficientes, así como sus solubilidades, tomando en cuenta los altos grados de
impurezas de las sales grado técnico o alimenticio aplicadas, en ocasiones
conformadas por conglomerados muy compactos.
La estructura de tallas en la población de filamentos de A. platensis permite explicar
mecanismos de autorregulación poblacional típicos de poblaciones con
densodependencia. A densidades poblacionales muy bajas, los recursos (luz y
nutrientes) son disponibles y predominan tricomas muy cortos y hormogonios
(filamentos reproductivos de las cianobacterias del Orden Oscillatoriales constituidos
por sólo dos células), lo que permite una adaptación rápida al medio y crecer en forma
exponencial, una vez se inicia la fase de crecimiento rápido. En el proceso de
maduración del cultivo, empiezan a dominar filamentos o tricomas de gran tamaño
(con muchas células) y en la fase de saturación la tasa de crecimiento instantánea se
reduce notablemente hasta llegar a cero en la capacidad de carga. En esta etapa no
hay producción de hormogonios y los filamentos maduros no reproductivos son el
componente de talla ampliamente predominante en el cultivo en fase de equilibrio.
Entonces, cuando la capacidad de carga es alcanzada, no hay incremento poblacional

122
y la población se atenúa u ocurren retrasos (oscilaciones amortiguadas y ciclos)
asociados a efectos de la densidad alrededor del punto de equilibrio estable (K).
La tasa de crecimiento per cápita explica la recuperación de una población luego de
una perturbación (May 1976). Una perturbación por extracción de una parte de la
población en estado de equilibrio puede ser respondida en función del tamaño de las
poblaciones remanentes en el medio y de su tasa intrínseca de crecimiento per cápita.
La tasa de crecimiento elevada de A. platensis, principalmente en condiciones
naturales, es debida a su forma de reproducción vegetativa por escisión de filamentos
y generación de hormogonios que se multiplican radialmente por fisión binaria en
pocos días. La perturbación no puede ser total o llegar a niveles basales, porque las
poblaciones remanentes en el cultivo son pequeñas y biológicamente inviables, lo que
puede conducir a la extinción. Una remoción de 75% y reemplazo con medio nutritivo
nuevo generó un efecto de dilución que no permitió que la población se adecuara en
forma rápida. Esto no es conveniente en términos biotecnológicos, puesto que se
busca maximizar el rendimiento de cosecha con una tasa de recuperación o resiliencia
elevada.
Como nota complementaria para el tema de poblaciones, llama la atención la
morfología rectilínea de los filamentos de A. platensis en condiciones de laboratorio
(Cámara de Crecimiento), cuyos filamentos originalmente tenían una estructura
helicoidal, típica del género. Una sucesión de varias generaciones desde el primer
inóculo en la Cámara de Crecimiento con exposición a luz artificial (400 lux) originó el
cambio a la forma rectilínea. Al respecto, se debe comentar que cuando la cepa fue
llevada al Ficotrón, y los cultivos se aclimataron a condiciones a cielo abierto, la misma
nuevamente empezó a adquirir la morfología helicoidal, luego de su exposición a la
radiación solar por tres generaciones sucesivas. La explicación a esta recuperación
está contenida en la calidad o composición de longitudes de onda de la luz solar,
específicamente a su componente de rayos ultravioletas, los que inducen a un
arrollamiento o arreglo de los tricomas en forma espiral, que generan en los filamentos
regiones alternas de mayor y menor exposición a las radiaciones más fuertes (Ciferri
1983).
En condiciones de laboratorio, la cianobacteria se adecuó a una fuente luz artificial
menos intensa (luz fría) y de menor calidad (a pesar del empleo de las denominadas
lámparas fluorescentes “day-light” que contienen la mayor parte de las longitudes de
onda del visible), y su disposición rectilínea permitió aumentar la superficie de
exposición de los filamentos para captar la luz con mayor eficiencia. En condiciones
naturales, las zonas oscuras o menos expuestas de los filamentos espirales o

123
helicoidales, como adaptación pertinente para hallar protección o menor exposición de
los filamentos a las radiaciones ultravioletas, suelen incrementar en forma notable las
concentraciones de ficocianinas, pigmentos antena exclusivos de las cianobacterias
que contribuyen a la captación de longitudes de onda adicionales a las captadas por la
clorofila a, y que contribuye al incremento de su eficiencia fotosintética. Resulta de
particular interés biotecnológico la producción de A. platensis (y A. máxima) en
condiciones naturales o semi-controladas a cielo abierto (estanques cerrados) para
obtener poblaciones con morfología helicoidal, por el gran atractivo que brindan las
ficocianinas a la industria de los colorantes y su empleo en salud y farmacéutica como
marcadores moleculares (Henrikson 1994).
8.3 Comunidades fitoplanctónicas
En aguas continentales, los humedales de Paria, Barlovento y Mantecal tienen como
característica común las presencias de grandes formaciones vegetales que ocupan la
mayor parte de su superficie, con predominio de plantas emergentes de hábito
herbáceo, de allí la denominación de humedales herbáceos (Mitsch y Gosselink 2007),
si bien las composiciones de especies, fisionomías y dominancias varían.
Asociaciones del fitoplancton y zooplancton a determinadas clases de vegetación
dentro de un mismo humedal se evidenciaron en análisis de componentes principales.
En el caso del humedal Palmares III, península de Paria, se encontró un patrón en
herradura de los parches de vegetación, lo que conlleva la presunción del
establecimiento de un gradiente espacial, con la composición de especies planctónicas
variando a lo largo de las zonas de vegetación monoespecífica y ecotonos. En este
mismo análisis multivariado se observaron relaciones bióticas entre especies
fitoplanctónicas, y de éstas con algunos organismos zooplanctónicos, como los
cladóceros, los cuales se alimentan del fitoplancton. La disposición parcelada de la
vegetación acuática genera asociaciones diferenciales de las comunidades del
fitoplancton y zooplancton en sus predios. Las intensidades variables de la radiación
solar bajo los techos de las asociaciones de plantas tienen causalidad inmediata en las
disposiciones espaciales o formas de crecimiento de las diferentes especies de
plantas emergentes. La consecuencia de esto es que la calidad de luz que se filtra
hasta la lámina de agua es escasa, por lo que la sombra generada va en detrimento
de la actividad fotosintética del fitoplancton, mientras que para el zooplancton
significan refugios eficaces de depredadores visuales.
La homogeneidad de buena parte de las variables abióticas en el agua robustece el
pensamiento de que la estructura aérea de la vegetación y ciertas condiciones

124
microclimáticas locales son las causas reales del establecimiento de patrones de
distribución espacial de las especies fitoplanctónicas y zooplanctónicas. En los
humedales ubicados en Paria, Barlovento y Mantecal contrastan en modo notable las
composiciones y abundancias de las especies entre los distintos parches de
vegetación. Por otra parte, las distintas zonas de vegetación generaron efectos locales
sobre algunas variables fisicoquímicas, como la concentración de oxígeno
(concomitante a cambios en la concentración de dióxido de carbono) y conductividad,
por tasas de descomposición diferenciales asociadas al tipo de material vegetal y
variaciones en las concentraciones de iones disueltos, respectivamente. En el caso del
humedal de Barlovento, tales cambios en la concentración de oxígeno y conductividad
se relacionaron con la disminución progresiva de la profundidad de la lámina de agua
por advenimiento de sequía, lo que produjo un efecto de concentración de iones. Las
asociaciones de plantas emergentes también establecen microclimas. Las
concentraciones de nutrientes, principalmente el fósforo y el nitrógeno, resultaron
bajas en ese humedal, lo que determina que el flujo de energía y materia para el
funcionamiento del ecosistema está determinado, en modo preponderante, por los
aportes de las plantas acuáticas en forma de hojarasca y materiales en distintos
estados de descomposición que constituye una fuente importante de nutrientes para la
comunidad del plancton (Mitsch y Gosselink 2007). Las cianobacterias pueden
contribuir al incremento de las fuentes de nitrógeno en el humedal mediante la fijación
de nitrógeno atmosférico.
El carácter salobre del humedal Palmares III, según sistema de clasificación de Tabilo-
Valdivieso (1999), se pone de manifiesto en la composición de especies
fitoplanctónicas descritas para este ambiente, en el cual se encontraron microalgas
típicas de agua dulce como clorofitas, euglenofitas y cianobacterias del género
Arthrospira, diatomeas con un espectro amplio de tolerancia a la salinidad (eurihalinas)
de los géneros Navicula y Nitzschia, así como pirrofitas (o dinoflagelados) del género
Protoperidinium y la cianobacteria Spirulina subsalsa, especies principalmente
relacionadas a ambientes estuarinos.
Para lagunas de inundación del bajo Orinoco, las grandes abundancias de
cianobacterias fijadoras de nitrógeno (en especial especies de los géneros Lyngbya y
Anabaena, éstas caracterizadas por sus grandes heterocistos), indican
concentraciones altas de este elemento en Los Pocitos, Macapaima y Ramonero. El
fitoplancton menos abundante en ambientes como Palital, Bañador y La Redonda, con
un número de especies muy alto en el Orden Desmidiales, sugiere que se trata de
ambientes limpios, oligotróficos y con aguas ligeramente ácidas (Comas 2008). Las

125
diatomeas resultaron muy abundantes en Las Palometas y La Redonda, este grupo es
muy importante para la alimentación del zooplancton encontrado en la zona,
principalmente rotíferos y cladóceros. La abundancia de diatomeas en esas lagunas
indica valores de pH ligeramente alcalinos, con presencia de silicatos (Wetzel 1975).
Por su parte, la laguna de Guarampo fue un ambiente diferente en cuanto a la
estructura de la comunidad fitoplanctónica, en este caso hubo abundancias menores
de la mayoría de sus componentes, con predominio notable de euglenofitas del género
Trachelomonas; este tipo de composición fitoplanctónica a menudo caracteriza a
ambientes ricos en materia orgánica y ligeramente eutrofizados (Comas 2008). La
relación estrecha de Guarampo con Palital, Las Palometas y La Redonda en el análisis
de componentes principales se debe a las densidades poblacionales menores
registradas en estos ambientes, mas no a las composiciones de especies, las cuales
son muy diferentes.
En general, para la mayoría de los ambientes de agua dulce estudiados la diversidad
fue alta donde las equidades, y en segundo término las riquezas, contribuyeron en
modo determinante a los valores del índice Shannon-Wiener. La reducción de los
valores de diversidad por equidades bajas incrementaron los índices de dominancia en
algunos sitios, donde se encontraron dominios amplios de algunas especies
fitoplanctónicas. Los estudios con índices de diversidad permiten caracterizar las
comunidades a un nivel simple, como sistemas independientes o aislados. Los índices
de distancia y similitud de Jaccard brindan comparaciones entre comunidades
distintas, donde se establece un segundo nivel de análisis de complejidad comunitaria
(diversidad beta) respecto al estudio de comunidades simples (diversidad alfa) (Del
Pino y col. 2006). En el humedal de Paria, las similitudes entre ambientes contiguos y
disimilitudes entre parches lejanos encontraron asidero en la propuesta de este índice
de complejidad al constatar, que efectivamente al haber pocas especies comunes a
ciertos ambientes, los clasifica como comunidades diferentes dentro de una gran
comunidad heterogénea dividida en ambientes parcelados.
Spirulina subsalsa, especie de gran valor biotecnológico, fue encontrada en Paria y
lagunas de inundación del Orinoco. Rodríguez (2001) la reporta en aguas salobres y
estuarinas, como el sistema del lago de Maracaibo (norte del lago). Esto evidencia que
S. subsalsa puede ser hallada en una gran variedad de ambientes, con una
distribución geográfica amplia en el país. En Barlovento y Mantecal se hallaron
algunas especies de cianobacterias de los géneros Lyngbya y Arthrospira, así como
clorofitas del género Scenedesmus, especies también referidas con potencialidades
importantes para la biotecnología. Las presencias de microalgas en ambientes

126
extremófilos, como salinas y lagunas hipersalinas, y altamente contaminados o
intervenidos, como las fosas petroleras y el muelle de Puerto Cabello, indican la
adaptabilidad de estos microorganismos a ambientes modificados por actividades
antrópicas. Sus potencialidades biotecnológicas se reflejan en sus capacidades para
vivir en tales ambientes, con capacidades para la absorción y resíntesis de
hidrocarburos a compuestos más simples, como Chlorella spp. y Scenedesmus spp.
(Albarracín 2007). Se conoce también la gran producción de pigmentos fotosintéticos
(carotenoides) de Dunaliella salina y D. viridis, especies de gran valor en la industria
de colorantes y en la producción de vitamina “A”, con poblaciones ubicadas en salinas
de Paraguaná y Laguna Boca Chica, isla de Margarita, respectivamente. Ambientes
marinos costeros no perturbados, como la bahía de Mochima, también ofrecen en su
diversidad algunas especies de gran valor biotecnológico para producción de
biocombustibles, Isochrysis galbana, Nitzschia valens y Chaetoceros spp.
Existe una gran variedad de ecosistemas terrestres y acuáticos en el país, producto de
su ubicación tropical y variedad de relieves, desde sistemas montañosos con alturas
importantes que modifican el clima, hasta llanuras continentales y costeras, vastos
territorios selváticos, depresiones, sistemas deltaicos y un gran frente marino. La
cuenca del río Orinoco sintetiza las tres grandes formas de relieve que existen en la
naturaleza: por un lado macizos antiguos y escudos, cordilleras de levantamiento
reciente (es decir, del Terciario) por el otro, y depresiones tectónicas y cuencas o
llanuras de acumulación, en tercer lugar. Para un país con estas características
geomorfológicas, y más si se encuentra en la zona intertropical, tal variedad ambiental
representa una gran ventaja ecológica y económica. Sólo Canadá y los Estados
Unidos, además de Venezuela y Colombia, que en su territorio tiene una parte
reducida del escudo guayanés, presentan una disposición geológica similar
(http://es.wikipedia.org/wiki/Río_Orinoco). Tal multiplicidad de ambientes produce una
gran complejidad de hábitats dulceacuícolas, salobres, marítimos y extremos, que
unida a la condición de país megadiverso, permite la exploración de comunidades
biológicas con un banco genético muy importante de especies promisorias para el
desarrollo de la biotecnología en campos como la alimentación, producción de
biocombustibles, biorremediación, farmacología, cosmética, colorantes, etc.

127
9. Conclusiones
1. Se obtuvieron dos medios de cultivo óptimos para el crecimiento de Arthrospira
platensis, uno en condiciones controladas (medio 1 = medio Parra (2005)) y otro en
condición natural (tratamiento 36, medio central), lo que inicia el aprovechamiento
biotecnológico de la especie a diferentes escalas.
2. La dinámica de crecimiento poblacional de Arthrospira platensis muestra
densodependencia logística, con evidencia de una estructura de tallas que varía
conforme la población crece y alcanza un equilibrio.
3. El crecimiento diferencial de poblaciones de Arthrospira platensis mostrado en
tratamientos con combinaciones aleatorias y centrales de nutrientes en un
experimento factorial fraccionado, señala que esta especie crece en condiciones
altamente selectivas.
4. La mayoría de los tratamientos con tiempos de duplicación, tg, más cortos (y por
ende tasas de crecimiento per cápita, r, más elevadas) se constituyeron en los
más productivos en biomasa.
5. La profundidad de la lámina de agua en sistemas de tanques circulares a cielo
abierto influye en la dinámica de crecimiento poblacional y producción de biomasa
de Arthrospira sp.
6. Se encontraron especies de cianobacterias (Arthrospira spp., Spirulina subsalsa,
Lyngbya spp., Oscillatoria spp. y Anabaena spp.) y microalgas eucarióticas
(Scenedesmus spp., Isochrysis galbana, Chlorella sp., Chaetoceros sp., Navicula
platalea, Dunaliella salina y D. viridis) de gran interés biotecnológico en diferentes
lugares de la geografía variada del país, lo que conduce a la idea de que
Venezuela cuenta con un potencial genético importante en su biodiversidad para la
biotecnología.

128
10. Bibliografía
Andrienko, N. y G. Andrienko 2006. Exploratory analysis of spatial and temporal Data. Springer-Verlag, Berlin, Heidelberg. 712 p.
Aiba, S. y T. Ogawa. 1977. Assessment of growth yield of a blue-green alga Spirulina platensis in axenic and continuous culture. J. Gen. Microbiol. 102: 179 – 182.
Albarracín, J. 2007. Microalgas: Potenciales productoras de biodiesel. XV Simposio Electrónico Internacional: La producción de biocombustibles con eficiencia, estabilidad y equidad. 16 p. En: http://www.ceid.edu.ar/biblioteca/biocombustibles /isabel_albarracin_microalgas_potenciales_productoras.pdf.
Anderberg, M.R. 1973. Cluster analysis for applications. Academia press, New York, NY. 359 p.
Ayala, F. y T. Vargas. 1987. Experiments on Spirulina culture on waste-effluent media at the pilot plant. Hydrobiologia 151/152: 91 – 93.
Azorín, F. 1970. Curso de muestreo y otras aplicaciones. Faces, Universidad Central de Venezuela. 346 p.
Banerjee, A., R. Sharma, Y. Chisti y U.C. Banerjee. 2003. Botryococcus braunii: a renewable source of hydrocarbons and other chemicals. Crit. Rev. Biotechnol.
22(3): 245 – 279.
Begon, M., J. Harper y C.R. Townsend. 1996. Ecology: individuals, populations and communities. Third Edition. Blackwell Science. 1068 p.
Berger, W.H. y F.L. Parker. 1970. Diversity of planktonic Foraminifera in deep-sea sediments. Science 168(3937): 1345 – 1347.
Blanco-Belmonte, L. 1990. Estudio de las comunidades de invertebrados asociados a las macrofitas acuáticas de tres lagunas de inundación de la sección baja del río Orinoco, Venezuela. Memoria Sociedad Venezolana de Ciencias Naturales La Salle 50(133-134): 71 – 107.
Bold, H. C. 1949. The morphology of Chlamydomonas chlamydogama, Sp. Nov. Bull. Torr. Bot. Club. 76: 101 – 108.
Borowitzka, M. 1999. Commercial production of microalgae: ponds, tanks, tubes and fermenters. Journal of Biotechnology 70: 313 – 321.
Boussiba, S. 1989. Ammonia Uptake in the Alkalophilic Cyanobacterium Spirulina platensis. Plant and Cell Physiology 30(2): 303 – 308.
Boussiba, S. y A.E. Richmond. 1979. Isolation and characterization of phycocyanins from the blue-green alga Spirulina platensis. Archives of Microbiology 120: 155 –
159.
Cañizares, R.O., A.R. Domínguez, L. Rivas, L. Travieso y F. Benítez. 1993. Free and immobilized cultures of Spirulina maxima for swine waste treatment. Biotech. Letters 15: 320 – 326.
Carmichael, W.W. 1994. The toxins of Cyanobacteria. Sci Am. 270(1): 78 – 86.
Casadevall, E., E. Dif, C. Largeau, C. Gudin, D. Chaumont y O. Desanti. 1985. Studies on batch and continuous cultures of Botryococcus braunii: hydrocarbon
production in relation to physiological state, cell ultrastructure, and phosphate nutrition. Biotechnol. Bioeng. 22: 286 – 295.

129
Caswell, H. 2001. Matrix Population Models: construction, analysis, and interpretation. Second Edition. Sinauer Associates, Inc. Publishers. Sunderland, Massachusetts. 722 p.
Chen, F., Y. Zhang y S. Guo. 1996. Growth and phycocyanin formation of Spirulina platensis in photoheterotrophic culture. Biotechnology Letters 18(5): 603 – 608.
Chen, F. y Y. Zhang. 1997. High cell density mixotrophic culture of Spirulina platensis on glucose for phycocyanin production using a fed-batch system. Enzyme and Microbial Technology 20: 221 – 224.
Cheng-Ng, R., H. Pons y E. Morales. 2005. Efectos inmunomoduladores de la cianobacteria Spirulina subsalsa. Memorias de la LV Convención Anual AsoVAC, Facultad de Ciencias, UCV, Caracas.
Chu, S.P. 1942. The influence of the mineral composition of the medium on the growth of planktonic algae. I. Methods and culture media. J.Ecol. 30: 284 – 325.
Ciferri, O. 1983. Spirulina, the edible microorganism. Microbiol. Rev. 47: 551 – 578.
Comas, A. 2008. Algunas características de la flora de algas y cianoprocariotas de agua dulce de Cuba. Algas 39: 21 – 29.
Conde, J.L., L.E. Moro, L. Travieso, E.P. Sánchez, A. Leiva y R. Dupeirón. 1993. Biogas purification process using intensive microalgae cultures. Biotech. Letters
15(3): 317 – 320.
Cressa, C., E. Vásquez, E. Zoppi, J.E. Rincón y C. López. 1993. Aspectos generales de la limnología en Venezuela. Interciencia 18(5): 237-248.
Dayananda, C., R. Sarada y V. Kumar. 2007. Isolation and characterization of hydrocarbon producing green alga Botryococcus braunii from Indian freshwater bodies. Electronic J. Biotech. 10(1): 78 – 91.
Del Pino, J.O., R. Zamora y J.A. Oliet. 2006. Empleo de diferentes índices de biodiversidad en los modelos basados en técnicas de decisión multicriterio. Universidad de Córdoba.
Duerr, E.O., M.R. Edralin y N.M. Price. 1997. Facilities requirements and procedures for the laboratory and outdoor raceway culture of Arthrospira spp. J Mar Biotechnol. 5: 1 – 11.
Dussart, B.H. 1984. Some Crustacea-Copepoda from Venezuela. Hydrobiologia 113:
25 – 174.
Edmonson, W.T. 1959. Freshwater biology. Second Edition. John Wiley & Sons, Inc, New York, NY. 1248 p.
Elster, H.J. y W. Ohle. 1978. Rotatoria: Die Rädertiere Mitteleuropas. Gebrüder Borntraeger, Berlin Stuttgart. 234 p.
Fábregas, J., C. Herrera, B. Cabezas y J. Abalde. 1985. Mass culture and biochemical variability of the marine microalgae Tetraselmis suecica Kylin (Butch) with high nutrient concentration. Aquaculture 49: 231 – 244.
Fluir, B. 1988. Common Principal Components and related multivariate methods. Wiley. 345 p.
García-Blairsy, G. 2008. Aplicaciones de los vegetales marinos. Centro de Biotecnología Marina. Universidad de Las Palmas. 81 p.
Garnier, F., J.P. Dubacq y J.C. Thomas. 1994. Evidence for a Transient Association of New Proteins with the Spirulina maxima Phycobilisome in Relation to Light Intensity. Plant Physiol. 106: 747 – 754.

130
Gente de Ciencias, A.C. 2007. Desarrollo y optimización del cultivo de microalgas promisorias en la nutrición animal (aves, porcinos, peces, camarones) y estructuración de la “Red Venezolana de Investigación en Microalgas”. Proyecto BID-Fonacit 2006000537. 1000 p.
González de Infante, A. y W. Riehl. 1992. Estudio taxonómico del fitoplancton del embalse de Guri (Venezuela). Acta Científica Venezolana 43: 190 – 199.
Gordon, E. y Y. Feo. 2007. Dinámica de crecimiento de Hymenachne amplexicaulis en un humedal herbáceo en el estado Miranda (Venezuela). Acta Bot. Venez. 30(1):
1 – 18.
Guevara, M., C. Lodeiros, O. Gómez, N. Lemus, P. Núñez, L. Romero, A. Vásquez y N. Rosales. 2005. Carotenogénesis de cinco cepas del alga Dunaliella sp. (Chlorophyceae) aisladas de lagunas hipersalinas de Venezuela. Rev. Biol. Trop.
53(3-4): 331 – 337.
Guillard, R.R.L. 1975. Culture of phytoplankton for feeding marine invertebrates. En: Smith, W.L. y M.H. Chanley (Eds.). Culture of Marine Invertebrates Animals. Plenum, New York. 29 – 60.
Hassell, M.P., J.H. Lawton y R. May. 1976. Patterns of Dynamical Behavior in single-species populations. J. Anim. Ecol. 45: 471 – 486.
Henrikson, R. 1994. Microalga Spirulina, superalimento del futuro. Segunda Edición.
Ediciones Urano. 60 p.
Hutchinson, G.E. 1961. The Paradox of the Plankton. The American Naturalist 95(882):
137 – 145.
Ichimura, T. 1996. Genome rearrangement and speciation in freshwater algae. Hydrobiologia 336: 1 – 17. En: Kristiansen, J. (Ed.). Biogeography of Freshwater
Algae. Developments in Hydrobiology. Kluwer Academic Publishers. Belgium. 165 p.
Infante, A. 1980. Los cladóceros del Lago de Valencia. Act. Cient. Venez. 31: 593 – 603.
Jaccard, P. 1901. Étude comparative de la distribution florale dans une portion des Alpes et des Jura. Bulletin de la Société Vaudoise des Sciences Naturelles 37: 547 – 579.
Jaime-Ceballos, B., H. Villarreal-Colmenares, T. García-Galano, R. Civera-Cerecedo y G. Gaxiola-Cortés. 2004. Empleo de polvo de Spirulina platensis en la alimentación de zoeas y mysis de Litopenaeus schmitti (Pérez-Farfante y Kensley, 1997). Memorias del VII Simposio Internacional de Nutrición Acuícola.
Jones, B.E. y W.D. Grant. 1999. Microbial diversity and ecology of the Soda Lakes of East Africa. Proceedings of the 8th International Symposium on Microbial Ecology. En: Bell C.R., Brylinsky M., Johnson-Green P. (Eds.). Atlantic Canada
Society for Microbial Ecology, Halifax, Canada.
Kjeldahl J. 1883. A new method for determining nitrogen in organic materials. Zeitschrift fur Analytische Chemie 22: 366 – 369.
L’évêque, C. 1987. Chad Basin. En: African Wetlands and Shallow Water Bodies (Zones humides et lacs peu profonds d'Afrique). Directory (Répertoire) M.J. Burgis y J.J. Symoens, (Eds.). ORSTOM, Coll. Travaux et Documents No. 211, Paris, 650 pp.
Mani, U., A. Sadliwala, U. Iyer y P. Parikh. 2000. The effect of Spirulina supplementation on blood hemoglobin levels of anemic adult girls. Journal of Food Science and Technology-Mysore 37(6): 642 – 644.

131
MARN. 1992. Estudio de la vegetación de la Península de Paria, Fase II. Informe técnico de la Dirección de Vegetación. Caracas. 87 p.
MARN. 1996. Información hidrológica y climatológica de Panaquire. Dirección de Hidrología y Meteorología. Caracas. 6 p.
MARN. 2000. Datos mensuales y anuales de precipitación de la Estación de Irapa, Edo. Sucre, 1953-2000. Dirección de Hidrología y Meteorología. Sistema Nacional de Información Hidrológica y Meteorológica. Caracas. 4 p.
Martínez-Palacios, C.A., M.C. Chávez-Sánchez, M.A. Olvera-Novoa y M.I. Abdo de la Parra. 1999. Fuentes alternativas de proteínas vegetales como substituto de la harina de pescado para la alimentación en acuicultura. Memorias del Tercer Simposio Internacional de Nutrición Acuícola, Monterrey, Nuevo León, México.
Mata García, L. 2003. Notas historiales sobre geología del petróleo en Margarita y Cubagua. En: http://www.angelfire.com/pro/petromar/.
Matto, C. y O. Parra. 1995. Introducción a la biología y sistemática de las algas de aguas continentales. Ediciones Universidad de Concepción, Santiago de Chile. 240 p.
May, R.M. 1976. Models for Single Populations. En: May, R.M. (Ed.). Theoretical Ecology. Blackwell Scientific Publications, London. 317 p.
Mcginnis, K.M., T.A. Dempster y M.R. Sommerfeld. 1997. Characterization of the growth and lipid content of the diatom Chaetoceros muelleri. Journal of Applied Phycology 9(1): 19 – 24.
Melack, J.M. 1979. Photosynthesis and Growth of Spirulina platensis (Cyanophyta) in an Equatorial Lake (Lake Simbi, Kenya). Limnology and Oceanography 24(4): 753 – 760.
Melack, J.M. 1988. Primary producer dynamics associated with evaporative concentration in a shallow, equatorial soda lake (Lake Elmenteita, Kenya). Hydrobiologia 158: 1 – 14.
Mercado, J.M. 1999. Fotosíntesis y cambios en la composición de la atmósfera. Ciencia al Día Internacional 2(2): 1 – 9.
Mitsch, W.J., y J.G. Gosselink. 2007. Wetlands. 4th Edition, John Wiley & Sons, Hoboken, NJ. 582 p.
Montgomery, D. 2001. Design and Analysis of Experiments. Fifth Edition. John Wiley & Sons, Inc. 684 p.
Mora, R., R. Moronta, J. Ortega y E. Morales. 2004. Crecimiento y producción de pigmentos de la microalga Chlorella sp. aislada de la Represa de Tulé, Municipio Mara, Estado Zulia, Venezuela. Ciencia 12(2): 117 – 124.
Morales, E., M. Rodríguez, D. García, C. Loreto y E. Marco. 2002. Crecimiento, producción de pigmentos y exopolisacáridos de la cianobacteria Anabaena sp. PCC 7120 en función del pH y CO2. Interciencia 27(7): 373 – 378.
Murphy, J. y Riley, J.P. 1962. A modified single solution method for the determination of phosphate in natural waters. Anal Chim. Acta 27: 31 – 36.
Myers, A.A. y P.S. Giller. 1991. Analytical Biogeography: An integrated approach to the study of animal and plants distributions. Chapman & Hall, London, Great Britain. 578 p.
Nandeesh, M.C., B. Gangadhara, J.K. Manissery y L.V. Venkataraman. 2001. Growth performance of two Indian major carps, catla (Catla catla) and rohu (Labeo

132
rohita) fed diets containing different levels of Spirulina platensis. Bioresource Technology 80: 117 – 120.
Naranjo-Briceño, L., D. Rojas-Tortolero, H. González, R. Torres, J. Zegarra, L. Sena-D’Anna y D. Sosa del Castillo. 2010. Arthrospira platensis como biofactoría de metabolitos secundarios de interés farmacológico: el ácido pipecólico. Rev Latinoam Biottecnol Amb Algal 1(1): 64 – 90.
Njuguna, G.S. 1988. Nutrient-phytoplankton relationships in a tropical meromictic soda lake. Hydrobiologia 158: 15 – 28.
Ortega, M.M. 1984. Catálogo de algas continentales recientes de México. Universidad Nacional Autónoma de México. México. 565 p.
Parra, J.A. 2005. Aislamiento y purificación de dos proteínas de la cianobacteria Spirulina platensis. Trabajo Especial de Grado, Universidad Central de
Venezuela, Caracas, Venezuela. 40 p.
Paulson, J. 2005. Development and assessment of models for predicting the phytoplankton assemblage patterns in Lake Kemp. Thesis in Fisheries Science, Faculty of Texas Tech University in Partial Fulfillment of the Requirements for the Degree of Master of Science. 68 p.
Peña, D. y J. Rodríguez. 2003. Descriptive measures of multivariate scatter and linear dependence. Journal of Multivariate Analysis 97: 609 – 619.
Pervushkin, S.V., A.V. Voronin, V.A. Kurkin, A.A. Sokhina e I.F. Shatalaev. 2001. Proteins from Spirulina platensis biomass. Chemistry of Natural Compounds
37(5): 476 – 481.
Pielou, E.C. 1975. Ecological diversity. John Wiley & Sons, Inc, New York. 165 p.
Prosperi, C.H. 2000. Cyanobacteria in human affaires. Interciencia 25 (6): 303 – 306.
Raoof, B., B. Kaushik y R. Prasanna. 2006. Formulation of a low-cost medium for mass production of Spirulina. Biomass and Bioenergy 30: 537 – 542.
Reid, J.W. 1984. Chave de identificação e lista de referências bibliográficas para as espécies continentais sulamericanas de vida livre da Ordem Cyclopoida (Crustacea, Copepoda). Bolm. Zool. Univ. S. Paulo. 9: 17 – 143.
Rodríguez, G. 2001. The Maracaibo System, Venezuela. En: Seeliger, U. y B. Kjerfve (Eds.). Coastal Marine Ecosystems of Latin American. Ecological Studies 144. Berlin, Springer Verlag. 355 p.
Rodríguez, J.C. y J.A. Betancourt. 1999. Caracterización físico-química de una laguna de inundación del tramo Orinoco medio y su relación con la biomasa de la cobertura de bora (Eichhornia crassipes (Mart.) Solms). Interciencia 24(4): 243 –
250.
Romero, Y., C. Lodeiros, M. Esclapé, N. Marín, M. Guevara y E. Morales. 2002. Efecto tóxico del cadmio sobre microalgas aisladas del nororiente de Venezuela. Interciencia 27(3): 104 – 109.
Sachdeva, R., R. Kaur y J. Kaur Sangha. 2004. Effect of Supplementation of Spirulina on the Haematological Profile and Intellectual Status of School Girls (7-9 years). J. Hum. Ecol. 15(2): 105 – 108.
Sánchez, M., J. Bernal-Castillo, C. Rozo e I. Rodríguez. 2003. Spirulina (Arthrospira): an edible microorganism. A review. En: www.javeriana.edu.co/universitas_ scientiarum/vol8n1/J_bernal.htm.
Scheaffer, R.L., W. Mendenhall y L. Ott. 1987. Elementos de muestreo. Grupo Editorial Iberoamérica S.A. de C.V., México. 322 p.

133
Schlösser, U.G. 1994. SAG-Sammlung von Algenkulturen at the University of Göttingen. Catalogue of Strains 1994. Bot. Acta 107: 113 – 186.
Seale, D.B., M.E. Boraas y G.J. Warren. 1987. Effects of sodium and phosphate on growth of cyanobacteria. Water research 21(6): 625 – 631.
Sena, L., D. Rojas, E. Montiel, H. González, J. Moret y L. Naranjo. 2011. A strategy to obtain axenic cultures of Arthrospira spp. Cyanobacteria. World Journal of Microbiology Biotechnology 27(5): 1045 – 1053.
Serpa, R.F. y A. Calderón. 2006. Efecto de diferentes fuentes de nitrógeno en el contenido de carotenoides y clorofila de cuatro cepas de Dunaliella salina TEOD. Ecología Aplicada 5(2): 93 – 99.
Shannon, C.E. y W. Weaver. 1949. The mathematical theory of communication. University Illinois Press, Urbana. 117 p.
Sim, T.S. y A. Goh. 1988. Ecology of microalgae in a high rate pond for piggery effluent purification in Singapore. World Journal of Microbiology and Biotechnology 4(3):
285 – 297.
Simpson, E.H. 1949. Measurement of diversity. Nature 163: 688.
Soler, A., M. Valdivia y O. Dieppa. 2000. Uso de la Spirulina como pigmentante de la piel y la grasa de los pollos de ceba. Rev. Fac. Cs. Vets. UCV. 41(1): 19 – 24.
Spolaore, P., C. Joannis-Cassan, E. Durán y A. Isambert. 2006. Commercial applications of microalgae. J Biosci Bioeng. 101(2): 87 – 96.
Tabilo-Valdivieso, E. 1999. Los beneficios de los Humedales en América Central: el potencial de los humedales para el desarrollo humano. 2da Edición. Programa Regional en Manejo de Vida Silvestre. Univ. Nac. de Heredia, Costa Rica. 58 p.
Tomaselli, L., M.C. Margheri y A. Sacchi. 1995. Effects of light on pigments and photosynthetic activity in a phycoerythrin-rich strain of Spirulina subsalsa. Aquatic Microbial Ecology 9: 27 – 31.
Torres, R. y Zoppi de Roa, E. 2010. Latencia en cladóceros y copépodos (Crustacea) de un humedal de la península de Paria, Venezuela. Métodos en Ecología y Sistemática 5(3): 22 – 35.
Utermöhl, H. 1958. Zur Vervollkommnung der quantitativen Phytoplankton Methodik. Mitt.Int.Ver Limnol. 9: 1 – 8.
Varela, M., R. Varela, E. Costas y A. Campos. 1986. Estudio al microscopio de transmisión de algunas diatomeas del río Orinoco y Delta Amacuro, Venezuela. Memoria Sociedad Venezolana de Ciencias Naturales La Salle 157: 49 – 67.
Velho, L.F.M. y F.A. Lansac-Tôha. 1996. Testate Amoebae (Rhizopoda - Sarcodina) from zooplankton of the High Paraná river floodplain, State of Mato Grosso do Sul, Brazil: II. Family Difflugidae. Stud Neotrop Fauna & Environm 31: 179 – 192.
Velho, L.F.M., F.A. Lansac-Tôha y M. Serafim-Junior. 1996. Testate Amoebae (Rhizopoda-Sarcodina) from zooplankton of the High Paraná river floodplain, State of Mato Grosso do Sul, Brazil: I. Families Arcellidae and Centropyxidae. Stud Neotrop Fauna & Environm 31: 35 – 50.
Vonshak, A. y A. Richmond. 1985. Problems in developing the biotechnology of algal biomass production. Plant and Soil 89(1-3): 129 – 135.
Wetzel, R.G. 1975. Limnology. Saunders College Publishing, Philadelphia. 743 p.
Woese, C.R. 1987. Bacterial evolution. Microbiol Rev. 51: 221 – 271.

134
Wood, R.B. y J.F. Talling. 1988. Chemical and algal relationships in a salinity series of Ethiopian inland waters. Hydrobiologia 158: 29 – 67.
Yacubson, S. 1974. El fitoplancton de la laguna de San Javier del Valle (Estado Mérida), Venezuela. Rexue Algologique 11(1-2): 91 – 131.
Zhang, Y-M. y F. Chen. 1999. A simple method for efficient separation and purification of c-phycocyanin and allophycocyanin from Spirulina platensis. Biotechnology Techniques 13: 601 – 603.
Zoppi de Roa, E. y W. Vásquez. 1991. Additional cladoceran records for Mantecal and new for Venezuela. Hydrobiologia 225: 45 – 62.
Zoppi de Roa, E. 1993. Nuevas adiciones a la fauna de rotíferos de Venezuela. Rev. Hydrobiol. Trop. 26 (3): 165 – 173.

135
11. Enlaces
http://es.wikipedia.org/wiki/Eustasia
http://es.wikipedia.org/wiki/Densidad_óptica
www.guiageo-americas.com/mapas/venezuela.htm
http://es.wikipedia.org/wiki/Barlovento_(Venezuela)
http://earth.google.com/
http://es.wikipedia.org/wiki/Cuenca_del_Orinoco
http://es.wikipedia.org/wiki/Río_Orinoco
http://es.wikipedia.org/wiki/Mantecal
http://es.wikipedia.org/wiki/Apure
http://www.disfrutevenezuela.com/Municipio-Peninsula-de-Macanao-Mapa.html
http://www.mochima.org/
http://www.tuplaya.com/paginasfalcon/salinas/lascumaraguas.htm
http://www.ipapc.gov.ve/
http://es.wikipedia.org/wiki/Puerto_Cabello
http://es.wikipedia.org/wiki/Nudo_(unidad)

136

137
Apéndice 1
Medios de cultivo para microalgas f/2 Medium (Guillard 1975) Fuente: http://www-cyanosite.bio.purdue.edu/media/table/f2.html
NaNO3 (75.0 g/L dH2O) 1.0 ml
NaH2PO4·H2O (5.0 g/L dH2O) 1.0 ml
Na2SiO3·9H2O (30.0 g/L dH2O) 1.0 ml
f/2 Trace Metal Solution 1.0 ml
f/2 Vitamin Solution 0.5 ml
Filtered seawater to 1.0 L
Autoclave after all additions.
Note: f/2 Medium contains extensive silica precipitate and should be used only when
growing diatoms. For other algal groups use f/2-Si Medium.
f/2 Trace Metal Solution:
FeCl3·6H2O 3.15 g
Na2EDTA·2H2O 4.36 g
CuSO4·5H2O (9.8 g/L dH2O) 1.0 ml
Na2MoO4·2H2O (6.3 g/L dH2O) 1.0 ml
ZnSO4·7H2O (22.0 g/L dH2O) 1.0 ml
CoCl2·6H2O (10.0 g/L dH2O) 1.0 ml
MnCl2·4H2O (180.0 g/L dH2O) 1.0 ml
Distilled water to 1.0 L
Autoclave.
f/2 Vitamin Solution:
Vitamin B12 (1.0 g/L dH2O) 1.0 ml
Biotin (0.1 g/L dH2O) 10.0 ml
Thiamine HCl 200.0 mg
Distilled water to 1.0 L
Filter sterilize into plastic vials and store in refrigerator.
Note: Vitamin B12 and Biotin are obtained in a crystalline form. When preparing the Vitamin B12 Stock Solution allow for approximately 11% water of crystallization (For each 1.0 mg of Vitamin B12 add 0.89
ml dH2O). When preparing the Biotin Stock Solution allow for approximately 4% water of crystallization
(For each 1.0 mg of Biotin add 9.6 ml dH2O).

138
Fuente:
Vera, A., M. Martínez, K. Morillo y S. Montes. 2011. Cultivo discontinuo de Chlorella sp. en medios enriquecidos con el exudado gomoso de Acacia macracantha. En:
http://revistas.luz.edu.ve/index.php/bcib/article/viewFile/90/3326.

139
Medio Chu-13 modificado por Dayananda (2007)
Modified CHU13 Medium, one liter [3]
Compound mg/L
KNO3 400
K2HPO4 80
CaCl2 dihydrate 107
MgSO4 heptahydrate 200
Ferric Citrate 20
Citric acid 100
CoCl2 0.02
H3BO3 5.72
MnCl2 tetrahydrate 3.62
ZnSO4 heptahydrate 0.44
CuSO4 pentahydrate 0.16
Na2MoO4 0.084
0.072 N H2SO4 1 drop
The remaining volume is pure, de-ionized water.
Fuente: http://en.wikipedia.org/wiki/Chu_13

140
Medio Bristol UTEX
Bristol Medium
Directions
H.C. Bold's modification of Bristol's recipe (Bold 1949). General purpose freshwater
medium and as bristol's solution, an essential component of other media--see Bold 1NV,
Bold 3N, Bristol-NaCl, LDM, Proteose, Soil extract, and Trebouxia.
For 1 L Total
1. To approximately 900 mL of dH2O add each of the components in the order specified
while stirring continuously.
2. Bring total volume to 1 L with dH2O.
*For 1.5% agar medium add 15 g of agar into the flask; do not mix.
3. Cover and autoclave medium.
4. Store at refrigerator temperature.
# Component Amount Stock Solution
Concentration
Final
Concentration
1 NaNO3 (Fisher
BP360-500) 10 mL/L 10 g/400mL dH2O 2.94 mM
2 CaCl2·2H2O (Sigma
C-3881) 10 mL/L 1 g/400mL dH2O 0.17 mM
3 MgSO4·7H2O
(Sigma 230391) 10 mL/L 3 g/400mL dH2O 0.3 mM
4 K2HPO4 (Sigma P
3786) 10 mL/L 3 g/400mL dH2O 0.43 mM
5 KH2PO4 (Sigma P
0662) 10 mL/L 7 g/400mL dH2O 1.29 mM
6 NaCl (Fisher S271-
500) 10 mL/L 1 g/400mL dH2O 0.43 mM
Fuente: http://web.biosci.utexas.edu/utex/mediaDetail.aspx?mediaID=29

141
Medio Serpa y Calderón (2005)
Fuente:
Serpa, R.F. y A. Calderón. 2006. Efecto de diferentes fuentes de nitrógeno en el contenido de carotenoides y clorofila de cuatro cepas de Dunaliella salina TEOD. Ecología Aplicada 5(2): 93 – 99.

142
Apéndice 2
ENSAYO Nro. 2: Optimización en botellones
Factorial 26-1
con 5 ptos. centrales Desing Expert
St. Id Run Block Type
NaHCO3 Na2CO3 K2HPO4 NaCl K2SO4 NH4NO3
mg/L mg/L mg/L mg/L mg/L mg/L
33 0 1 Block 1 Center 10207,5 3022,5 375 750 750 1764
28 28 2 Block 1 Fact 13610 4030 250 1000 1000 1176
2 2 3 Block 1 Fact 13610 2015 250 500 500 2352
11 11 4 Block 1 Fact 6805 4030 250 1000 500 1176
14 14 5 Block 1 Fact 13610 2015 500 1000 500 2352
25 25 6 Block 1 Fact 6805 2015 250 1000 1000 1176
30 30 7 Block 1 Fact 13610 2015 500 1000 1000 1176
22 22 8 Block 1 Fact 13610 2015 500 500 1000 2352
6 6 9 Block 1 Fact 13610 2015 500 500 500 1176
7 7 10 Block 1 Fact 6805 4030 500 500 500 1176
27 27 11 Block 1 Fact 6805 4030 250 1000 1000 2352
3 3 12 Block 1 Fact 6805 4030 250 500 500 2352
34 0 13 Block 1 Center 10207,5 3022,5 375 750 750 1764
4 4 14 Block 1 Fact 13610 4030 250 500 500 1176
31 31 15 Block 1 Fact 6805 4030 500 1000 1000 1176
10 10 16 Block 1 Fact 13610 2015 250 1000 500 1176
12 12 17 Block 1 Fact 13610 4030 250 1000 500 2352
24 24 18 Block 1 Fact 13610 4030 500 500 1000 1176
5 5 19 Block 1 Fact 6805 2015 500 500 500 2352
18 18 20 Block 1 Fact 13610 2015 250 500 1000 1176
23 23 21 Block 1 Fact 6805 4030 500 500 1000 2352
29 29 22 Block 1 Fact 6805 2015 500 1000 1000 2352
35 0 23 Block 1 Center 10207,5 3022,5 375 750 750 1764
8 8 24 Block 1 Fact 13610 4030 500 500 500 2352
26 26 25 Block 1 Fact 13610 2015 250 1000 1000 2352
13 13 26 Block 1 Fact 6805 2015 500 1000 500 1176
20 20 27 Block 1 Fact 13610 4030 250 500 1000 2352
21 21 28 Block 1 Fact 6805 2015 500 500 1000 1176
1 1 29 Block 1 Fact 6805 2015 250 500 500 1176
9 9 30 Block 1 Fact 6805 2015 250 1000 500 2352
17 17 31 Block 1 Fact 6805 2015 250 500 1000 2352
15 15 32 Block 1 Fact 6805 4030 500 1000 500 2352
32 32 33 Block 1 Fact 13610 4030 500 1000 1000 2352
16 16 34 Block 1 Fact 13610 4030 500 1000 500 1176
19 19 35 Block 1 Fact 6805 4030 250 500 1000 1176
36 0 36 Block 1 Center 10207,5 3022,5 375 750 750 1764
37 0 37 Block 1 Center 10207,5 3022,5 375 750 750 1764

143
Design-Expert® Software
Biomasa
Design Points
E- 500.000
E+ 1000.000
X1 = A: NaHCO3
X2 = E: K2SO4
Actual Factors
B: Na2CO3 = 3022.50
C: K2HPO4 = 375.00
D: NaCl = 750.00
F: NH4NO3 = 1764.00
E: K2SO4
6805.00 8506.25 10207.50 11908.75 13610.00
Interaction
A: NaHCO3
Bio
ma
sa
0.8
1.2
1.6
2
2.4
Design-Expert® Software
Biomasa
Design Points
F- 1176.000
F+ 2352.000
X1 = D: NaCl
X2 = F: NH4NO3
Actual Factors
A: NaHCO3 = 10207.50
B: Na2CO3 = 3022.50
C: K2HPO4 = 375.00
E: K2SO4 = 750.00
F: NH4NO3
500.00 625.00 750.00 875.00 1000.00
Interaction
D: NaCl
Bio
ma
sa
0.8
1.2
1.6
2
2.4

144
Design-Expert® Software
Biomasa
Design Points
B- 2015.000
B+ 4030.000
X1 = A: NaHCO3
X2 = B: Na2CO3
Actual Factors
C: K2HPO4 = 375.00
D: NaCl = 750.00
E: K2SO4 = 750.00
F: NH4NO3 = 1764.00
B: Na2CO3
6805.00 8506.25 10207.50 11908.75 13610.00
Interaction
A: NaHCO3
Bio
ma
sa
0.8
1.2
1.6
2
2.4
Design-Expert® Software
Biomasa
Design Points
E- 500.000
E+ 1000.000
X1 = A: NaHCO3
X2 = E: K2SO4
Actual Factors
B: Na2CO3 = 3022.50
C: K2HPO4 = 375.00
D: NaCl = 750.00
F: NH4NO3 = 1764.00
E: K2SO4
6805.00 8506.25 10207.50 11908.75 13610.00
Interaction
A: NaHCO3
Bio
ma
sa
0.8
1.2
1.6
2
2.4

145
Design-Expert® Software
Biomasa
Design points above predicted value
Design points below predicted value
2.3555
0.838
X1 = A: NaHCO3
X2 = E: K2SO4
Actual Factors
B: Na2CO3 = 3022.50
C: K2HPO4 = 375.00
D: NaCl = 750.00
F: NH4NO3 = 1764.00
6805.00
8506.25
10207.50
11908.75
13610.00 500.00
625.00
750.00
875.00
1000.00
1.2
1.525
1.85
2.175
2.5
B
iom
asa
A: NaHCO3 E: K2SO4
Design-Expert® Software
Biomasa
Design points above predicted value
Design points below predicted value
2.3555
0.838
X1 = D: NaCl
X2 = F: NH4NO3
Actual Factors
A: NaHCO3 = 10207.50
B: Na2CO3 = 3022.50
C: K2HPO4 = 375.00
E: K2SO4 = 750.00
500.00
625.00
750.00
875.00
1000.00 1176.00
1470.00
1764.00
2058.00
2352.00
1.4
1.64
1.88
2.12
2.36
B
iom
asa
D: NaCl F: NH4NO3

146
Design-Expert® Software
Biomasa
Design points above predicted value
Design points below predicted value
2.3555
0.838
X1 = A: NaHCO3
X2 = B: Na2CO3
Actual Factors
C: K2HPO4 = 375.00
D: NaCl = 750.00
E: K2SO4 = 750.00
F: NH4NO3 = 1764.00
6805.00
8506.25
10207.50
11908.75
13610.00 2015.00
2518.75
3022.50
3526.25
4030.00
1.2
1.525
1.85
2.175
2.5
B
iom
asa
A: NaHCO3 B: Na2CO3

147
Apéndice 3 Fuente: http://bellman.ciencias.uniovi.es/d_experimentos/d_experimentos_archivos/sr.pdf
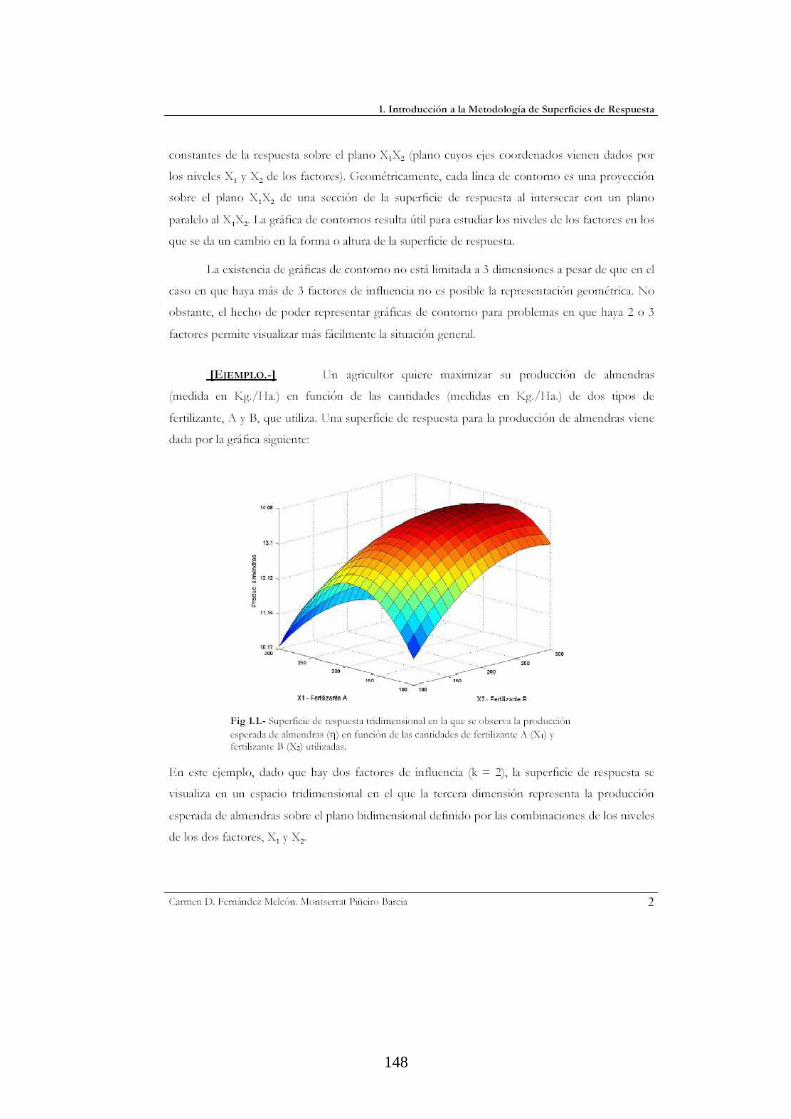
148

149

150


















