
Studies to Evaluate Disinfectant Efficacy and Facility Disinfection ...

Gene Therapy and Molecular Biology Vol 12, page 371
371
Gene Ther Mol Biol Vol 12, 371-382, 2008
A short term pilot open label study to evaluate efficacy and safety of LG839, a customized DNA directed nutraceutical in obesity: Exploring Nutrigenomics Research Article Kenneth Blum1,3,8,9,*,#, Thomas JH Chen2, Lonna Williams3, Amanda LC Chen2,*, William B. Downs3, Roger L. Waite3, T. Huntington3, Simon Sims4, Thomas Prihoda5, Patrick Rhoads6, Jeffrey Reinking7, Dasha Braverman8, Mallory Kerner8, Seth H. Blum9, Brien Quirk10, Eric R Braverman11 1Department of Physiology & Pharmacology, Wake Forest University School of Medicine, Winston-Salem, North Carolina, USA 2Chang Jung Christian University, Tainan, Taiwan Republic of China 3Department of Nutrigenomics, LifeGen®, Inc. La Jolla, California, USA 4 DNA Services of America, Lafayette, Louisiana, USA 5 Department of Pathology, University of Texas Health Science Center, San Antonio, Texas, USA 6 Central Valley Comprehensive Obesity Clinic, Modesto, California, USA 7Department of Medicine, University of California School of Medicine at Davis, Davis California, USA 8Department of Clinical Neurology, Path Medical and Research Foundation, New York, New York, USA 9 Department of Psychoneurogenetics, Synaptamine, Inc., San Antonio, Texas, USA 10 Draco Natural Products, Inc. Eugene, Oregon, USA 11Department of Neurosurgery, Weill Cornell College of Medicine, New York, New York, USA __________________________________________________________________________________ *Correspondence: Kenneth Blum, Ph.D., Department of Physiology & Pharmacology, Wake Forest University School of Medicine, Medical Center Boulevard, Winston-Salem, North Carolina, 27157, USA; Tel: 210-823-1504; e-mail [email protected] Amanda LC Chen, Chang Jung Christian University, Tainan, Taiwan Republic of China: e-mail [email protected] #All reprint request to Kenneth Blum e-mail: [email protected] Key words: LG839; Synaptamine™, Reward Deficiency Syndrome (RDS), Obesity, Diabetes, nutrigenomics, polymorphisms, genes Abbreviations: Allelic Discrimination, (AD); body mass index, (BMI); cardiovascular disease, (CVD); CCAAT/enhancer-binding protein alpha, (C/EBPα); cocaine- and amphetamine-regulated transcript, (CART); deoxynucleotide 5’-triphosphates, (dNTPs); dopamine D2 receptor, (DRD2); adrenergic receptors, (ADBRs); interleukin-6, (IL-6); leptin receptor, (LEPR); leptin, (LEP); lipid metabolism including hepatic lipase, (LIPC); lipoprotein lipase, (LPL); logarithm of odds, (LOD); methylenetetrahydrofolate reductase, (MTHFR); perilipin, (PLIN); peroxisome proliferator activator receptor delta, (PPARδ); Peroxisome Proliferator-Activated Receptor gamma, (PPAR-γ); polymerase chain reaction, (PCR); quantitative trait loci, (QTL); restriction fragment length polymorphism, (RFLP); serotonin receptor, (5-HT2a); sterol regulatory element-binding transcription factor 1, (SREBP-1); uncoupling proteins, (UCPs) Conflict of Interest: Kenneth Blum, Roger L. Waite, Lonna Williams, B William Downs, own stock in LifeGen® Inc. Synaptamine ™ and LG839 and all patent variants have been exclusively licensed to LifeGen® Inc.
Received: 7 October 2008; Revised: 3 December 2008
Accepted: 5 December 2008; electronically published: January 2009
Summary We hypothesized that genotyping certain known candidate genes would provide DNA-individualized customized nutraceuticals that may have significant influence on body re-composition by countering various genetic traits. It is well known that obesity and related symptoms significantly aggravates type 2 diabetes. Both obesity and diabetes are influenced by the interaction of both genes and environmental factors. Exploration of the current literature has identified a number of candidate genes to be associated with both of these two disorders and include amongst others the dopamine D2 receptor (DRD2), methylenetetrahydrofolate reductase (MTHFR), serotonin receptor (5-HT2a),

Blum et al: Exploring Nutrigenomics
372
Peroxisome Proliferator-Activated Receptor gamma (PPAR-γ), and Leptin (OB) genes. In the present study, we systematically evaluated the impact of polymorphisms of these five candidate genes as important targets for the development of a DNA-customized nutraceutical LG839 [dl-phenylalanine, chromium, l-tyrosine other select amino-acids and adaptogens]) to combat obesity with special emphasis on body recomposition as measured by Body Mass Index (BMI). A total of 21 individuals were evaluated in a preliminary investigational study of LG839. Based on the results of buccal swab genotyping of each subject, an individualized customized nutraceutical formula was provided as a function of measured gene polymorphisms of the five gene candidates assessed. At the inception of the study and every two weeks subsequently, each subject completed a modified Blum-Downs OPAQuE Scale™ [Overweight Patient Assessment Questionnaire]. The alleles included the DRD2 A1; MTHFR C 677T; 5HT2a 1438G/A; PPAR-γPro12A1a and Leptin Ob1875<208bp. Pre- and post ad hoc analysis revealed a significant difference between the starting BMI and the BMI following an average of 41 days (28-70d) of LG839 intake in the 21 individuals. The pre- BMI was 31.2 (weight/Ht2) compared to the post BMI of 30.4 (weight/Ht2) with a significance value of P < 0.034 (one tailed). Similarly the pre -weight in pounds (lb) was 183.52 compared to the post weight of 179 lb with a significance value of P<(0.047). We also found trends for reduction of late night snacking, carbohydrate craving reduction, reduction of stress, reduction of waist circumference. Moreover, in the 41 day period we found a trend in weight loss whereby 71.4% of subjects lost weight. Thus 15 out of 21 subjects lost weight with a z score of 2.4 and significance value of P <(0.02). In this group 53% lost on average over 2.5% of their starting weight. Further confirmation of these preliminary results (ongoing) warrants investigation and should ultimately provide novel DNA directed “omic” therapeutic targets of novel anti-obesity agents especially in diabetes and other related diseases.
I. Introduction Our generating hypothesis was that genotyping
certain known candidate genes would provide DNA-individualized customized genetic targets by manipulation of specific ingredient components of a nutraceutical that may have significant influence on body re-composition by countering various genetic traits involved in obesity-related deficient pathways. This paper will present preliminary evidence to support both safety and efficacy potential of LG839 and provide a brief review of the literature of gene-environmental influence on obesity risk.
A. Genes and obesity One example of how genes may affect behavior
related to obesity involves the genetic aspects of the dorsal striatum of the brain. The dorsal striatum plays a role in consummatory food reward, and striatal dopamine receptors are reduced in obese individuals, compared to non-obese subjects. This suggests that the striatum and dopaminergic transmission in this brain region contribute to the development of obesity. Stice and colleagues in 2008 using functional MRI found that subjects may overeat to compensate for a blunted or hypofunctioning dorsal striatum, especially in subjects carrying the dopamine D2 A1 variant (DRD2A1 allele). Certainly we are cognizant that multiple gene polymorphisms play a role in obesity and related behaviors.
Rapid progress in human genome decoding has accelerated the search for the role of gene polymorphisms in the pathogenesis of complex multifactorial diseases. It is evident that several major chronic diseases are closely related (often through obesity) to deregulation of energy homeostasis. Multiple polymorphic genes encoding central and peripheral determinants of energy intake and expenditure have been revealed over the past decade (Martinez et al, 2008). Food intake control may be affected by polymorphisms in the genes encoding taste receptors and a number of peripheral signaling peptides
such as insulin, leptin, ghrelin, cholecystokinin, and corresponding receptors. Polymorphic central regulators of energy intake include hypothalamic neuropeptide Y, agouti-related protein, melanocortin pathway factors, CART (cocaine- and amphetamine-regulated transcript), some other neuropeptides, and receptors for these molecules. Potentially important polymorphisms in the genes encoding energy expenditure modulators (alpha- and beta- adrenoceptors, uncoupling proteins, and regulators of adipocyte growth and differentiation and the dopamine D2 receptor genes ) related gene polymorphisms comprising those involved in the pathogenesis of obesity, blood pressure regulation, hemostasis control, and homocysteine metabolism are known and these multiple polymorphisms affect lipid transport and metabolism and their interactions with diet. There is an apparent progress in the field with hundreds of new gene polymorphisms discovered and characterized, however firm evidence consistently linking them with pathogenesis of complex chronic diseases is still limited. It appears that several new directions emerge as targets of research on the role of genetic variation in relation to diet and complex chronic diseases. Regulation of energy homeostasis is a fundamental problem insufficiently investigated in this context so far (Martinez et al, 2008). Impacts of genetic variation on systems controlling angiogenesis, inflammatory reactions, and cell growth and differentiation (comprising regulation of the cell cycle, DNA repair, and DNA methylation) are also largely unknown and need thorough analysis. However, these goals can be achieved by complex simultaneous analysis of multiple polymorphic genes controlling carefully defined and selected elements of relevant metabolic and regulatory pathways in meticulously designed large-scale studies (Loktionov, 2003).
Indeed, a number of genes involved in the regulation of energy expenditure, appetite, lipid metabolism and adipogenesis have been reported to affect the risk of treatment failure in some obese subjects. Some candidate

Gene Therapy and Molecular Biology Vol 12, page 373
373
genes for the prognosis of weight loss response related to energy expenditure are those codifying for the adrenergic receptors (ADBRs) and uncoupling proteins (UCPs), while genes related to appetite potentially affected by energy restriction are leptin (LEP), leptin receptor (LEPR), melanocortin pathways genes (MC3R, POMC), dopamine D2 receptor (DRD2) and the serotonin receptor. Furthermore, adipogenesis related genes such as peroxisome proliferator-activated receptor gamma (PPARγ2) (Rosmond et al, 2003) and genes related to cytokines such as interleukin-6 (IL-6) and lipid metabolism including hepatic lipase (LIPC), perilipin (PLIN) and lipoprotein lipase (LPL) have also been associated with the weight lowering outcome induced by hypocaloric diets. Therefore, preliminary evidence from human studies supports the existence of a genetic component in the fat reduction process associated to a negative energy balance (Table 1).
B. Polygenetic approaches Obesity has become a global epidemic and
contributes to the increasing burden of type 2 diabetes, cardiovascular disease, stroke, some types of cancer, and premature death worldwide. Obesity is highly heritable and arises from the interactions of multiple genes, environmental factors, and behavior. Heritabilities for obesity-related phenotypes varies from 6% to 85% among various populations. As of October 2005, 253 quantitative trait loci for obesity-related phenotypes have been localized in 61 genome-wide linkage scans, and genetic variants in 127 biologic candidate genes have been reported to be associated with obesity-related phenotypes from 426 positive findings. Gene-gene interactions have
been also observed in several genes, and some genes were found to influence the effect of dietary intake and physical activity on obesity-related phenotypes. Integration of genetic epidemiology with functional genomics and proteomics studies will be required to fully understand the role of genetic variants in the etiology and prevention of obesity (Barnard et al, 2008).
Moreover, the concept of gene-environment interaction is receiving support from emerging evidence coming primarily from studies involving diet and cardiovascular disease (CVD), diabetes and various risk factors. The accumulating evidence shows that common variants at candidate genes for lipid metabolism, inflammation, and obesity are associated with altered plasma levels of classic and new biomarkers of metabolic syndrome. Major contributors to this knowledge have been a series of large population studies containing phenotype-rich databases and dietary information to which genetic data have been added (Ordovas and Shen, 2008).
Current population studies are starting to incorporate experimental and analytical approaches that could provide more solid and comprehensive results. However, other limitations, such as the size of the populations required to examine higher-level interactions, are still major obstacles to translating this knowledge into practical public health applications. Nevertheless, data from numerous molecular and genetic epidemiological studies provide tantalizing evidence suggesting that gene-environment interactions, i.e., the modulation by a genetic polymorphism of a dietary component effect on a specific phenotype (e.g., cholesterol levels and obesity), can interact in ways that increase the risk for developing chronic disease, including susceptibility to developing the metabolic syndrome.
Table 1. Number of Scientific Studies Validating Specific Genes Analyzed in Proposed Panel for Nutrigenomic Application. As of July 31, 2008
Gene General Reward & appetite regulation
Stress Metabolism & energy
Neuro-endocrine
Immune system
Leptin OB 3618 57 104 981 3 108 Serotonergic 2A receptor Gene 289 13 8 2 6 4 Phenylethanolamine-N-Methyltransferase 247 3 38 1 6 5
Dopamine D2 receptor 1999 334 31 8 73 23
Tumor necrosis factors (or the TNF-family 593 -0- 37 443 -0- 160
Steroid sulfatase (STS) gene 156 -0- -0- 2 -0- 15 peroxisome proliferator-activated receptor γ (PPARγ) 2878 -0- 100 87 1 303
Carbohydrate responsive element-binding protein (ChREBP) gene 27 -0- -0- 24 27 -0-
FTO Gene 107 -0- -0- 10* -0- -0- Monoamine Oxidase 882 47 61 7 18 15 ADAM 8-ADAM metallopeptidase domain 8, also known as ADAM8 13 -0- -0- 1 1 -0-
Corticotrophin-Releasing Factor (CRF) 612 -0- 243 16 202 221
Vitamin D gene 5443 -0- 64 82 28 223

Blum et al: Exploring Nutrigenomics
374
Once further experience is gained from patients and/or individuals at high risk, more personalized genetic-based approaches may be applied toward the primary prevention and treatment of obesity.
While there is enough data from human studies it is important to support these studies with animal experimentation, albeit possibility of non-relevance between murine genes and human obesity, utilizing a well established approach referred to as QTL. Cross-breeding experiments with different mouse strains have successfully been used by many groups to identify genetic loci that predispose for obesity. In order to provide a statistical assessment of these quantitative trait loci (QTL) as a basis for a systematic investigation of candidate genes, Wuschke and colleagues have performed in 2007 a meta-analysis of genome-wide linkage scans for body weight and body fat. From a total of 34 published mouse cross-breeding experiments, they compiled a list of 162 non-redundant QTL for body weight and 117 QTL for fat weight and body fat percentage. Collectively, these studies include data from 42 different parental mouse strains and >14,500 individual mice. The analysis revealed significant evidence (logarithm of odds (LOD) score >4.3) for linkage of body weight and adiposity to 49 different segments of the mouse genome. The most prominent regions with linkage for body weight and body fat (LOD scores 14.8-21.8) on chromosomes 1, 2, 7, 11, 15, and 17. At least 34 candidate genes and genetic loci, which have been implicated in regulation of body weight and body composition in rodents and/or humans, are found in these regions, including CCAAT/enhancer-binding protein alpha (C/EBPα), sterol regulatory element-binding transcription factor 1 (SREBP-1), peroxisome proliferator activator receptor delta (PPARδ), and hydroxysteroid 11-beta dehydrogenase 1 (HSD11B1). The risks for any individual to become obese may involve the impairment of many physiological processes’ and a number of major pathways. The next section will provide specific information related to these defined pathways.
C. Genes and specific pathways The following Table 1 provides the number of
scientific studies of various known obesity related genes that our laboratory has proposed for the development of a nutrigenomic panel to counter certain genetic traits. This table is presented to inform the reader as to the extent of the literature.
D. Genes and BMI It is well known that a number of genes are
associated with body mass index (BMI) and we will briefly discuss only the serotonin and dopamine receptor genes as two examples.
In terms of serotonin (5HT2a receptor polymorphisms) it was reported that in an obese population case control study, when only the obese group was analyzed, it was observed that AA subjects presented a lower body mass index (BMI) than G allele carriers (35.2+/-5.3 kg/m2 vs 37.5+/-7.8 kg/m2; P=0.039). Moreover, significant differences were also obtained in waist perimeter that was lower in AA subjects compared
to G allele carriers (105+/-11 cm vs 112+/-17 cm; P=0.011). In conclusion, although the -1438G>A polymorphism is not a relevant marker for obesity risk, this variant may play a role in determining BMI in obese subjects (Sorlí et al, 2008). Moreover a similar result was obtained for polymorphisms of the DRD2gene. In Pima Indians with diabetes individuals with a 'CG' genotype at the Ser311Cys SNP of the Dopamine D2 receptor gene had a higher BMI than those with a 'CC' genotype (36.7 vs 35.5 kg/m2, P= 0.04).
Furthermore, in neuroimaging studies (Volkow et al, 2008), the associations between striatal D2 receptors and prefrontal metabolism in obese subjects suggest that decreases in striatal D2 receptors could contribute to overeating via their modulation of striatal prefrontal pathways, which participate in inhibitory control and salience attribution. The association between striatal D2 receptors and metabolism in somatosensory cortices (regions that process palatability) could underlie one of the mechanisms through which dopamine regulates the reinforcing properties of food and may ultimately contribute to BMI.
No obesity disease-causing genes, however, have been conclusively identified, most likely because this is a multigenic disease for which effects of many causative genes may be small and combined with environmental effects. However, while there are more sophisticated approaches such as Prioritisation of candidate disease genes (Tiffin et al, 2008), less sophisticated clinical approaches are still informative.
E. Other supporting nutrigenomic studies In exploring the literature to determine results of
other laboratories that may also utilize a nutrigenomic approach to counter genetic traits related to obesity treatment PUBMED analysis only resulted in a citation of our original research in this emerging field.
In an earlier study in this journal we have discussed the role of Nutrigenomics in obesity (Chen et al, 2008). It is our contention that unraveling the multitude of nutrigenomic (nutritional genomics), transriptomic (gene expression), epigenomic (organization of the chromatin) proteomics (posttranslational modifications) and metabolomic (metabolic profiles) patterns that arise from nutrient-induced bioactive cellular mechanisms are likely to provide insights into a tailored approach to adhere to a diet and induce health and lead to anti-obesity novel personalized medicine and nutrition (Trujillo et al, 2006).
In this regard our laboratory has embarked on a nutrigenomic approach and recently published a number of studies suggesting the validity of this approach (Meshkin et al, 2008). Additionally, in a recent published study (Blum et al, 2008b), whereby five candidate genes and associated polymorphism were assessed: methylenetetrahydrofolate reductase-k, MTHFR (folate metabolism); Peroxisome Proliferator-Activated Receptor- PPRA-γ2 (lipid regulator); serotonin receptor -5-HT2a (sweet taste); Leptin-Leptin Ob (obesity); and dopamine D2 receptor -DRD2 gene (CART). Outcome results of this earlier pilot revealed significant benefits of DNA customization of LG839 (80 days of treatment) as follows:

Gene Therapy and Molecular Biology Vol 12, page 375
375
weight loss (p <0.008); sugar craving reduction (p<0.008); appetite suppression (p<0.004); snack reduction (p< 0.005); reduction of late night binging (p<0.007); increased perception of over-eating (p<0.02)]; increased energy (p<0.004); enhanced quality of sleep (p<0.02) and increased happiness (p<0.02).
Other work from our laboratory showed a significant association of the DRD2 gene Taq 1A allele and percent body fat (Chen et al, 2007a). Additionally our laboratory showed an association of the PPAR -γPro 12 allele with a self identified obese Dutch population compared to unscreened control Dutch population (Chen et al, 20007b). In follow-up studies carriers of the DRD2 gene Taq 1A allele significantly complied with days on treatment with LG839 compared to DRD2A2 carriers by a factor of 2 fold (Blum et al, 2008a). Over the last two decades earlier work from our laboratory clinically evaluated a patented complex Synaptamine™ as an anti- glucose craving agent in a number of studies with positive outcomes: a 90 day out-patient barometric clinic (Blum et al, 1990); a 2-year out-patient barometric clinic compared to vitamin placebo (Blum et al, 1997); and a one year open trial in a consumer based cross sectional study providing a PATH analysis (Blum et al, 2006).
In consideration of all of above we decided to further evaluate LG839 customized for each patient following genotyping of MTHFR, PPRA-γ2, -5-HT2a, Leptin Ob and the DRD2 genes in a short term (41 days) open label study whereby each subject served as their own control.
II. Materials and Methods A. Subjects A total of 21 individuals (1 males and 20 females; mean
age 37.5 ±1.7 yr, range 22-51yr) clients of DNA Services of America, a DNA service company located in Lafayette, Louisiana, were evaluated in a preliminary investigational study of LG839. The IRB of the Path Research & Medical Foundation approved the project (NIH Registration # IRB00002334 ) and all participants signed an approved consent form prior to the study. The subjects received educational material about the experimental concept, however were only told to eat appropriately. No attempt was made to control diet as well as to exercise (i.e. no attempt to develop a standardized exercise program).
B. Blum-Downs OPAQuE Scale™ This cross-sectional, observational study included a genetic
analysis involving a panel of genes, and an evaluation questionnaire providing two key self-reported retrospective data sets on the subjects’ health before taking LG839 and then after taking LG839. Self-reported evaluations were collected through an online questionnaire disseminated over email to study participants. These subjects with a starting BMI of 31.2 were evaluated by survey analysis over 41(28-70 d) days of administration of LG839. Based on the results of buccal swab genotyping of each subject, a customized nutraceutical formula was provided as a function of measured gene polymorphisms of the five gene candidates assessed. At the inception of the study and every two weeks subsequently, each subject completed a modified Blum-Downs OPAQuE Scale™ [Overweight Patient Assessment Questionnaire-see questions Table 2]. The BMI was subsequently calculated by our staff using this self-guiding survey.
C. Study outcomes The primary objective was to evaluate the safety and
efficacy of LG839 in obese and overweight subjects, and specifically to look at differences in weight, mood, sleep, digestion, wellness and other health issues before and after taking LG839. All participants provided written consent, and the study protocol was approved by the institutional review board of the non-profit research organization, PATH Medical & Research Foundation, located in New York, NY USA. Data were collected on line independently by LooseFoot Computing Ltd. of Regina, Saskatchewan, Canada and manually by two of us (LW and TH). Genotyping of the five candidate genes for polymorphic alleles resulted in a number of significant associations of this population. These alleles included the DRD2 A1; MTHFR C 677T; 5HT2a 1438G/A; PPAR -γPro12A1a and Leptin Ob1875<208bp.
D. Laboratory measurements and genotyping The laboratory testing was performed in a high-
complexity, CLIA-certified laboratory under contract with the company based in Los Angeles, California, USA. Genotyping was carried out as a research project. All subjects were genotyped based on a neutral identification number and read without knowledge of the individual being typed. Total genomic DNA was extracted from each coded blood sample, and aliquots were used for polymerase chain reaction (PCR) analysis.
E. DRD2 The DRA1 and DRA2 genotyping was performed using
PCR (The oligo- nucleotide primers 5’-CCGTCGACCCTTCCTGAGTGTCATCA-3’ (Blum et al 1990; Comings et al, 1996) and 5’CCGTCGACGGCTGGCCAAGTTGTCTA-3’were used to amplify a 310-base pair (bp) fragment spanning the polymorphic Taq1A1 site of the DRD2 gene. PCR was performed in 30mL reaction mixtures containing 1.5mM MgCl2, Table 2. Blum-Downs OPAQuE Scale™ [Overweight Patient Assessment Questionnaire
Assessment area and question* Anxiety Appetite Drug craving Tobacco craving Depression Ease of Sleep Energy Exercise Get along with others Happiness Interrupted sleep Late night eating Perception of Over-eating Quality of sleep Regularity of digestion Snacking Sugar craving Waist (cm) Weight (kg)
*Responses during LG839 administration minus baseline responses. BMI was calculated on each study participant by LW, TH and TP.

Blum et al: Exploring Nutrigenomics
376
2mM 2’- deoxynucleotide 5’-triphosphates (dNTPs), 0.5 uM primers, 1 μg of template DNA 1, 5U of Taq polymerase (Boehringer Mannheim Corp., Indianapolis, IN), and PCR buffer (20 mM Tris-HCL [pH 8.4] and 50mM KCl). After an initial denaturation at 94ºC for 4 minutes, the DNA was amplified with 35 cycles of 30 seconds at 94ºC, 30 seconds at 58ºC, and 30 seconds at 72ºC, followed by a final extension step of 5 minutes at 72ºC. The PCR product was digested with 5 U of Taq 1 for 22 hours at 65ºC for the Taq1A polymorphism. Digestion products were then resolved on a 3% agarose gel (5V/cm) containing 0.65 ug/ml ethidium bromide. There were three DRD2 Taq1A genotypes: 1) the predominant homozygote A2/A2, which exhibits two restriction fragments of 180 and 130 bp; 2) the heterozygote A1/A2, which exhibits three restriction fragments of 310, 180, and 130bp; and 3) the rare homozygote A1/A1, which produces only the uncleaved 310-bp fragment
F. MTHFR Genotyping for the MTHFR C677T polymorphism was
performed using PCR and restriction fragment length polymorphism (RFLP) analysis. We designed PCR primers 5’-CCCAGCCACTCACTGTTTTAG-3’ and 5’-TGGGAAGAACTCAGCGAACT-3’ with DNASIS Pro Ver.2.0 (Hitachi Software Engineering Co. Ltd., Tokyo, Japan). Since the C to T transition at nucleotide 677 produces a Hinf I digestion site, the amplified 469-bp product derived from the mutant gene was cleaved into 393-bp and 76-bp fragments by Hinf I (TaKaRa Bio Inc., Shiga, Japan), which leaves the wildtype gene unaffected. After electrophoresis through 6% polyacrylamide gel, the digestion products were visualized by staining with ethidium bromide.
G. 5-HT2a Genotyping of the -1438G/A polymorphism of the 5-HT2A
gene was carried out by PCR and restriction digestion as described previously (Nonogaki et al, 2006). Genomic leukocyte DNA (100ng in a final volume of 10μL) was amplified by PCR using the following primers: 5’-AAGCTGCAAGGTAGCAACAGC-3’ and 5’-AACCAACTTATTTCCTACCAC-3’. The primers amplified a product of 468 bp. The PCR conditions were as follows: an initial denaturation step at 94 °C for 3 minutes, followed by 35 cycles of denaturation at 94 °C for 15 seconds, annealing at 55 °C for 15 seconds, and extension at 72 °C for 30 seconds, with a final extension of 10 minutes at 72 °C. The PCR reaction product was digested at 37 °C overnight with 5 U of the restriction enzyme MspI (New England Biolabs, Beverly, MA). The -1438G allele was cut into 244-bp and 224-bp fragments, whereas the -1438A allele remained undigested. The fragments were separated on a 2% agarose gel and visualized by staining with ethidium bromide.
H. PPAR-γ2 Two hundred base pairs of sequence surrounding PPAR-γ2
Pro12Ala was provided to Applied Biosystems (Foster City, CA) to develop Taqman Allelic Discrimination (AD) assays using their assay by design platform. Genotyping of the Pro12Ala AD was performed using the following primers (0.9 molL each); Forward 5'-TTATGGGTGAAACTCTGGGAGATT-3' and reverse 5'-TGCAGACAGTGTATCAGTGAAGGA-3' and the Taqman MGB probes Fam- TTCTGGGTCAATAGG and Vic-CTTTCTGCGTCAATAG (0.1moli/L each; Applied Biosystems). Four μl of a 10 ng/L stock of DNA was dispensed into 384-well PCR plates using a Biomek FX robot (Beckman Coulter), to which 6uL of a mix containing primers, TaqMan
MGB probes and TaqMan Universal PCR Master Mix (Applied Biosystems) were added. These were sealed with optical seals (Applied Biosystems) and incubated at 95°C for 10 min followed by 40 cycles of 95°C for 15 s and 60°C for 1 min before analysis on a 7900HT plate reader (Applied Biosystems). Individual genotypes were determined using SDSv2.1 software (Applied Biosystems) (Rosmond et al, 2003).
I. Leptin (OB) We used the following dinucleotide repeats (D7S1873,
D7S1875, D7S514 and D7S680) present on the YAC contig containing the human OB gene as described by Liu and colleagues in 2007.The primers used for PCR were the same as in their report. Of these, D7S1875 was closest to the OB gene. We refer to this as OB1875. The frequency of the different alleles of the OB1875 polymorphism has been reported previously (Comings et al 1996). The alleles ranged in size from 199 to 225 bp in length. Our a priori approach to the examination of dinucleotide repeats in behavioral disorders is based on other research (Rance et al, 2007). Comingsand colleagues in 1996 was to first divide the alleles into approximately equal groups, since there was a natural tendency for a bimodal distribution, which was the case for the OB1875 polymorphism and the cut was made at 208 bp. The resulting genotypes were <208 bp/<208 bp, <208 bp/≥208 bp, and ≥208 bp/≥208bp. In this experiment we used Coming’s cut off point in our analysis providing for only positive and negative allelic associations.
J. Nutraceutical customization Based upon the genetic profiles derived, certain ingredients
in LG839 were customized according to subjects’ genotype (Table 3).
K. Measurements of Efficacy and Safety Separate from the laboratory measurements, all
measurements of efficacy and safety were self-reported by the study subjects using a one-time retrospective online questionnaire. The primary endpoint was weight loss in kilograms and BMI (the subject of this report). Secondary endpoints included changes in appetite, sugar cravings, waist circumference, mood, sleep, and digestion (the subject of on-going catchment data for subsequent analysis with increasing numbers of study subjects). The changes in these factors were measured generally using a scale of 1 to 5 where 5 was the least healthy rating and 1 was the most healthy rating to provide consistency across the multiple measures. Tertiary endpoints included changes in incidences of depression and cravings for alcohol/tobacco/recreational drugs. To monitor persistency, we asked study subjects to self-report on their frequency of compliance to the product’s serving instructions.
L. Statistics The means and standard deviations were calculated before
and during LG839 -DNA based customized treatment for each item in the questionnaire. To test for a change in weight, the paired t-test (one - tailed) was used on the change score, with change calculated as response during treatment minus the response before treatment for each item. The BMI at start was compared with the last measurement ranging from 28-70 days with an average of 41 days. The Statistical Analysis System (SAS Institute, version 9.1 for the PC, Cary, North Carolina, 2006) was used for these calculations. Due to the small sample size we used a one-tail test instead of a 2 tailed test.

Gene Therapy and Molecular Biology Vol 12, page 377
377
Table 3. Customization of the nutraceutical LG839 according to genotype in 21 subjects
Genotype Ingredient Serving MTHFR C677T Homozygous or Normal/Wild Type (CC)1-12 Vitamin B9 (folic acid) 800 µg
MTHFR C677T Heterozygous Mutant (CT) or Homozygous Mutant (TT)8 Vitamin B9 (folic acid) 5 mg
5-HT2A -1438A Heterozygous or Normal 13 SuperCitrimax (-) Hydroxycitric Acid (extract of Garcinia Cambogia) 2,500 mg
5-HT2a – 1438A Heterozygous or Homozygous Mutant or Leptin OB 1875 14
SuperCitrimax (-) Hydroxycitric Acid (extract of Garcinia Cambogia) 4,667 mg
Dopamine D2 Receptor Wild type A2/A2 14-16
Synaptamine Complex (DL-phenylalanine, Chromium, L-glutamine, L-tyrosine, L-5-HTP, Vitamin B6 [pyridoxal-5-phosphate]),Rhodiola rosea, passion flower
1,725 mg
Dopamine D2 Receptor Heterzygous A1/A2 or Homozygous A1/A1
Synaptamine Complex (DL-phenylalanine, Chromium, L-glutamine, L-tyrosine, L-5-HTP, Vitamin B6 [pyridoxal-5-phosphate]), Rhodiola rosea, passion flower
2,750 mg
Reproduced from Blum et al, 2008 with kind permission from Advance Therapeutics. Note: In terms of weight loss formulations the PPAR gene variation is linked to a novel substance Irvingia gobonensis seed extract shown to up-regulate the adiponectin gene (Oben JE, Ngondi JL, Blum K (2008) Inhibition of Irvingia gabonensis seed extract (OB131) on adipogenesis as mediated via down regulation of the PPARgamma and Leptin genes and up-regulation of the adiponectin gene. Lipids Health Dis 7, 44. References 1. Federation of American Societies for Experimental Biology, Life Sciences Research Office (1995) Prepared for the Interagency Board for Nutrition Monitoring and Related Research. Third Report on Nutrition Monitoring in the United States: Volumes 1 and 2. U.S. Government Printing Office, Washington, DC. 2. Grosse SD, Waitzman NJ, Romano PS Mulinare J (2008) Reevaluating the benefits of folic Acid fortification in the United States: economic analysis, regulation, and public health. Am J Public Health 95, 1917-1922. 3. Huhta JC, Hernandez-Robles JA (2005) Homocysteine, folate, and congenital heart defects. Fetal Pediatr Pathol 24(2):71-9. 4. Homocysteine Lowering Trialists' Collaboration. (2005) Dose-dependent effects of folic acid on blood concentrations of homocysteine: a meta-analysis of the randomized trials. Am J Clin Nutr 82, 806-12. 5. Grundy SM, Cleeman JI, Daniels SR, Donato KA, Eckel RH, Franklin BA, Gordon DJ, Krauss RM, Savage PJ, Smith Jr SC, Spertus JA, Costa F (2005) Diagnosis and management of the metabolic syndrome. An American Heart Association/National Heart, Lung, and Blood Institute Scientific Statement. Executive summary. Cardiol Rev 13, 322-7. 6. Russo GT, Di Benedetto A, Alessi E, Giandalia A, Gaudio A, Ientile R, Horvath KV, Asztalos B, Raimondo G, Cucinotta D (2006) Mild hyperhomocysteinemia and the common C677T polymorphism of methylene tetrahydrofolate reductase gene are not associated with the metabolic syndrome in Type 2 diabetes. J Endocrinol Invest 29, 201-7. 7. Thawnashom K, Tungtrongchitr R, Petmitr S, Pongpaew P, Phonrat B, Tungtrongchitr A, Schelp FP (2005) Methylenetetrahydrofolate reductase (MTHFR) polymorphism (C677T) in relation to homocysteine concentration in overweight and obese Thais. Southeast Asian J Trop Med Public Health 36, 459-66. 8. Sheu WH, Chin HM, Lee WJ, Wan CJ, Su HY, Lang HF (2005) Prospective evaluation of folic acid supplementation on plasma homocysteine concentrations during weight reduction: a randomized, double-blinded, placebo-controlled study in obese women. Life Sci 76, 2137-2145. 9. Henning BF, Tepel M, Riezler R, Gillessen A, Doberauer C (1998) Vitamin supplementation during weight reduction--favourable effect on homocysteine metabolism. Res Exp Med (Berl) 198, 37-42. 10. Volek JS, Gomez AL, Love DM, Weyers AM, Hesslink R Jr, Wise JA, Kraemer WJ (2002) Effects of an 8-week weight-loss program on cardiovascular disease risk factors and regional body composition. Eur J Clin Nutr 56, 585-92. 11. Dixon JB, Dixon ME, O'Brien PE (2001) Elevated homocysteine levels with weight loss after Lap-Band surgery: higher folate and vitamin B12 levels required to maintain homocysteine level. Int J Obes Relat Metab Disord 25, 219-27. 12. Gallistl S, Sudi K, Mangge H, Erwa W, Borkenstein M (2000) Insulin is an independent correlate of plasma homocysteine levels in obese children and adolescents. Diabetes Care 23, 1348-52. 13. Thom E (1996) Hydroxycitrate (HCA) in the Treatment of Obesity. Int J Obes Relat Metab Disord 20(suppl 4), 75. 14. Preuss HG, Bagchi D, Bagchi M, Ohia SE, Bramble JD, Garis R, Rao CV (2004) Efficacy of a Novel, Natural Extract of (-)-Hydroxycitric Acid (HCA-SX) and a Combination of HCA-SX, Niacin Bound Chromium and Gymnema sylvestre Extract in Weight Management in Human Volunteers: A Pilot Study. Nutrition Research 24, 45-58 15. Chen TJ, Blum K, Payte JT, Schoolfield J, Hopper D, Stanford M, Braverman ER (2004) Narcotic antagonists in drug dependence: pilot study showing enhancement of compliance with SYN-10, amino-acid precursors and enkephalinase inhibition therapy. Med Hypotheses 63, 538-48. 16. Milner JD, Irie K, Wurtman RJ (1986) Effect of phenylalanine on the release of endogenous dopamine from rat striatal slices. J Neurochem 47, 1444-1448.
Blum et al: Exploring Nutrigenomics
378
III. Results In the present study, the genotype data is the subject
of another report elsewhere and in the present study it was used solely to define the customization requirements of the
administered formula. We found that the following percentage genotypes (Drd2A1; MTHFRC 677T; 5Ht2a 1438g?A; PPAR-Gamma Pro12A1a and Leptin OB1875<280bp):
Gene combinations** Percentage Presence of Polymorphism*
MTHFR 11.81 SEROTONIN 7.84 SEROTONIN ÷ MTHFR 13.42 SEROTONIN ÷ MTHFR ÷LEP 5.77 SEROTONIN ÷ PPAR ÷ MTHFR 5.58 DOPAMINE ÷ MTHFR 5.29 DOPAMINE ÷ SEROTONIN 4.54 DOPAMINE ÷ SEROTONIN ÷ MTHFR 10.40 ** These are the combinations found for the five candidate genes
A total of 21 individuals were evaluated in a preliminary investigational study of LG839. These subjects with a starting BMI of 31.2 were evaluated by survey analysis over 41(28-70 d) days of administration of LG839. Based on the results of buccal swab genotyping of each subject, a customized nutraceutical formula was provided as a function of measured gene polymorphisms of the five gene candidates assessed.
Genotyping of the five candidate genes for polymorphic alleles resulted in a number of significant associations of this population. These alleles included the DRD2 A1; MTHFR C 677T; 5HT2a 1438G/A; PPAR -γPro12A1a and Leptin Ob1875<208bp. Pre- and post ad hoc analysis revealed a significant difference between the starting BMI and the BMI following 41 days of LG839 intake in the 21 individuals. The pre- BMI was 31.2 SEM
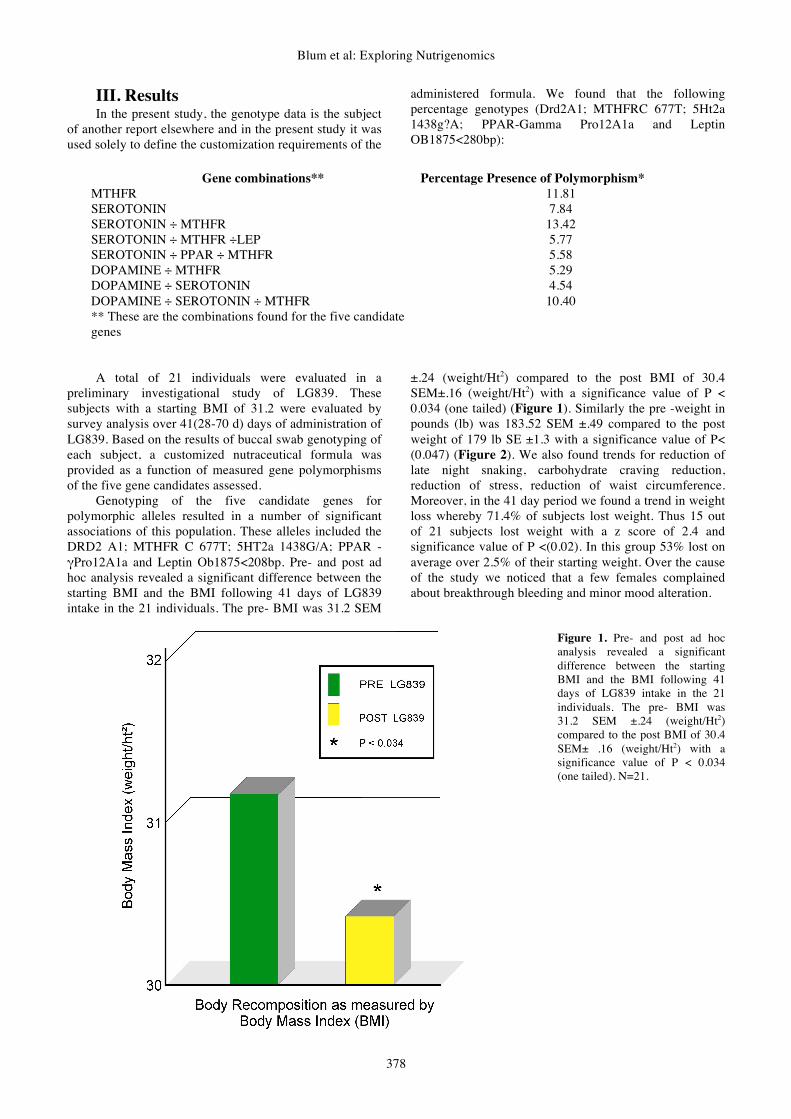
±.24 (weight/Ht2) compared to the post BMI of 30.4 SEM±.16 (weight/Ht2) with a significance value of P < 0.034 (one tailed) (Figure 1). Similarly the pre -weight in pounds (lb) was 183.52 SEM ±.49 compared to the post weight of 179 lb SE ±1.3 with a significance value of P< (0.047) (Figure 2). We also found trends for reduction of late night snaking, carbohydrate craving reduction, reduction of stress, reduction of waist circumference. Moreover, in the 41 day period we found a trend in weight loss whereby 71.4% of subjects lost weight. Thus 15 out of 21 subjects lost weight with a z score of 2.4 and significance value of P <(0.02). In this group 53% lost on average over 2.5% of their starting weight. Over the cause of the study we noticed that a few females complained about breakthrough bleeding and minor mood alteration.
Figure 1. Pre- and post ad hoc analysis revealed a significant difference between the starting BMI and the BMI following 41 days of LG839 intake in the 21 individuals. The pre- BMI was 31.2 SEM ±.24 (weight/Ht2) compared to the post BMI of 30.4 SEM± .16 (weight/Ht2) with a significance value of P < 0.034 (one tailed). N=21.

Gene Therapy and Molecular Biology Vol 12, page 379
379
Figure 2. The one tailed pre -weight in pounds (lb) was 183.52 SEM ±.49 compared to the post weight of 179 lb SEM ± 1.3 with a significance value of P< (0.047) N=21.
IV. Discussion It is well known that obesity and related symptoms
significantly aggravates type 2 diabetes. Both obesity and diabetes are influenced by the interaction of both genes and environmental factors. These findings take on an even greater importance relative to the well known neuro- interaction of dopamine and glucose (Blum et al, 2006). Confirmation of reduced glucose craving behavior by LG839 may reduce the risk of a number of genes which affect aberrant behavior for type 2 diabetics. Barnard and colleagues found in 2008 that DRD2 A1 allele was highly prevalent, occurring in 47% of white participants (n = 49), which was significantly higher than the 29% prevalence previously reported in non-diabetic whites (P = 0.01). The A1 allele was found in 55% of black participants (n = 44). Black participants with A1(+) genotypes had significantly greater mean body weight (11.2 kg heavier, P = 0.05) and greater intake of fat (P = 0.002), saturated fat (P = 0.01), and cholesterol (P = 0.02) compared with A2A2 (A1(-)) individuals while dietary changes during the study did not favor one genotype group.
It is noteworthy that one mechanism may involve the role of chromium salts (an ingredient in LG839), a known anti-diabetic compound affecting glucose utilization/body composition as a function of the DRD2 gene polymorphisms (Chen et al, 2007a), whereby carriers of the DRD2 A1 allele did not respond to chromium picolinate compared to DRD2 A2 carriers. Thus previous work suggests the need to prior genotype individuals to assist in obtaining information concerning proper dosing. The findings of this report coupled with our most recent
publications (Blum et al 2008a,b) further support a nutrigenomic approach to weight management.
Other more recent work investigated reward sensitivity with regard to binge eating behavior. The sensitivity of dopamine reward pathways has been implicated in the risk for various psychiatric disorders including compulsive overeating. The evidence is divided, however, about the direction of causal association. One argument is that a Reward Deficiency Syndrome (RDS) is the risk factor, while others contend that hyper-sensitivity to reward enhances the motivation for pleasurable activities like eating. Davis and colleagues found in 2008 genotype x group ANOVAs revealed significant main effects and an interaction on the personality measures for Taq1A. Binge Eating Disorder (BED) and obese subjects reported greater reward sensitivity than normal-weight controls, but only among those carrying the A1 allele. They also found that normal-weight controls with at least one copy of the T allele of the C957T marker had significantly lower reward sensitivity scores than any of the other groups who did not differ from each other. Given evidence linking the A1 allele with reduced receptor density, an inverse relationship was expected between psychological measures of reward sensitivity and presence of the A1 allele. One explanation for their findings could be that the BED and obese participants possess another genetic variant that interacts with the A1 allele to produce higher dopamine activity. However, in RDS it is possible that low dopaminergic activity may translate to aberrant reward hypersensitivity because of the anxious need to obtain a dopamine “fix” (Blum et al, 2006; Davis et al, 2008).

Blum et al: Exploring Nutrigenomics
380
While the present study represents a small sample of individuals who purchased the DNA customized LG839 at DNA-Services of America clinics across the nation, the significant reduction of BMI as well as s significant decrease in weight suggests the need for additional studies involving a tailored personalized DNA based approach. We are somewhat surprised in that LG839 -DNA based was able to cause significant changes in both BMI and weight in subjects self-Identified as desirous of utilizing this anti-obesity regimen in only 41 days (an average days on treatment).We further caution any interpretation of these results because we utilized a less conservative one-tailed statistic to analyze the data.
These results are consistent with earlier reports involving LG839 variants (Blum et al, 1990, 1997, 2006). Furthermore, a loss of 2.5 kg in only 41 days is quite robust especially when considering guidelines from the United Sates FDA regarding meaningful weight loss of approved medical use of any anti-obesity pharmaceutical. The NI H 1998 Practical Guide: Identification, Evaluation, and Treatment of Overweight and Obesity in Adults defines normal weight as a body mass index (BMI) of 18.5 - 24.9 kg/m2, overweight as a BMI of 25 - 29.9 kg/m2, and obese as a BMI ≥30 kg/m2. The Practical Guide recommends weight loss through a combination of diet modification, increased physical activity, and behavior therapy for obese patients, and for patients who are overweight or have a high-risk waist circumference, when accompanied by two or more risk factors. In the event that lifestyle changes do not promote weight loss after 6 months, the Practical Guide recommends that drugs should be considered as adjunctive therapy for select patients who have gender based BMI ≥30 kg/m2, or a BMI ≥27 kg/m2 if concomitant obesity-related risk factors or disease exist.
The FDA’s approach to the approval of prescription weight-loss drugs mirrors the recommendations provided in the NIH’s Practical Guide. For example, the prescription of orlistat was studied and approved for long-term weight loss in patients moderately-to-severely overweight (BMI 27-9.9 kg/m2) with comorbid conditions such as hypertension, type 2 diabetes, or dyslipidemia and in obese subjects (BMI ≥30 kg/m2) regardless of the presence of comorbidities. Limiting approval of prescription weight-loss drugs to moderately-to-severely overweight individuals with comorbidities and to obese patients maximizes the therapeutic risk-benefit profile by targeting drug therapy to individuals whose risk for weight-related disease is high and likely to outweigh the risks associated with any given pharmacological agent. In clinical studies, orlistat demonstrated weight loss of 5% or more at 6 and 12 months (Food And Drug Administration Center For Drug Evaluation and Research (CDER), 2006). In light of the current results which suggest a potential weight loss of 2.5% in 41 days (in this group 53% lost on average over 2.5% of their starting weight), or less than 3 months, the LG839 product warrants further investigation to evaluate whether these pilot study results are replicable in a randomized, placebo-controlled study of a longer duration.
Limitations of this present study involving sample size and lack of placebo controls caution any definitive interpretation of these results albeit significant findings, and must await further confirmation. In this present pilot study which was an extension of previously reported outcomes with LG839 (Blum et al, 2008b), each patient served as it’s own control. In future studies albeit difficult and costly, we intend to develop studies which will provide a placebo group matched to each individualized DNA solution. We believe that the breakthrough bleeding one of the ingredients in the formulation (which will be altered in future studies) and the mood alteration may be due to enhanced serotonin levels. In an open label study because a control is not employed interpretation of the result is suspect.
Since our formula includes both dl-phenylalanine and chromium we find it of considerable interest that a novel chromium -d-phenylalanine complex in other studies possesses better cardio-protective and insulin-sensitizing properties against obesity in ob/ob mice than chromium picolinate alone (Dong et al, 2007). A major benefit of this novel compound is the potential of eliminating the proposed cytoclastic toxicity of chromium picolinate alone. This novel complex which is imbedded in Synaptamine™ will be the subject of future investigation. Of great interest is the finding of slowed intracellular Ca2+
decay observed in myocytes from ob/ob mice following a 3 month treatment with Cr(D-Phe)3.This takes on real importance when we consider the important role of intracellular calcium in neurotransmitter release of neuronal dopamine. A substance linked to glucose craving (Blum et al, 2007).
V. Conclusion Based on this preliminary investigation and others
(Blum et al 2008 a,b) we suggest that DNA-directed targeting of certain regulator genes along with customized nutraceutical intervention may provide a unique framework and strategic modality to combat obesity, weight problems, and their associated co-morbidities. In light of many dietary supplement ingredients and other tactics used empirically for weight loss without preliminary analysis to deem their merit, this study maintains previous foundational research involving the LG839 technology that a scientific and rationale approach of laboratory testing and laboratory-directed nutritional supplementation may present beneficial health outcomes in the treatment of obesity, along with healthy eating and exercise. This pioneering research will pave the way for additional gene-environmental studies utilizing nutigenomics as a principle modality in the combat of obesity related diseases including larger samples and longer term therapy. One major outcome if this pilot study suggests that LG839 is well tolerated in subjects in the short term and other work from our lab supports this notion as well (Chen et al, 2008b).
Thus confirmation of these preliminary results (ongoing) will support our hypothesis and warrants investigation. These and other large independent controlled reports should ultimately provide novel DNA directed “omic” therapeutic targets of novel anti-obesity

Gene Therapy and Molecular Biology Vol 12, page 381
381
agents especially in diabetes and other obesity related diseases.
Acknowledgements The authors appreciate the funding support of the
PATH Medical & Research Foundation, New York, New York, LifeGen, Inc. La Jolla, California and Electronic Waveform Lab, Huntington , Beach California. They especially appreciate the donations of Don Reuben, Eduardo Cruz, Rein Norma, and Elizabeth Daiber.
References
Barnard ND, Noble EP, Ritchie T, Cohen J, Jenkins DJ, Turner-McGrievy G, Gloede L, Green AA, Ferdowsian H (2008) D2 dopamine receptor Taq1A polymorphism, body weight, and dietary intake in type 2 diabetes. Nutrition, in press.
Blum K, Chen ALC, Chen TJH, Rhoades P, Prihoda TJ, Downs BW, Bagchi, Bagchi M, Blum SH, Williams L, Braverman ER, Kerner M, Waite RL, Quirk B, White L (2008a) Dopamine D2 Receptor Taq A1 allele predicts treatment compliance of LG839 in a subset analysis of a pilot study in the Netherlands. Gene Ther Mol Biol 12, 129-140.
Blum K, Chen ALC, Chen TJH, Rhoades, Prihoda TJ, Downs BW, Waite RL, Williams L, Braverman Er, Braverman D, Arcuri V, Kerner M, Blum SH, Reinking J, Palomo T (2008b) LG839 (an experimental DNA-Customized nutraceutical): Anti-obesity effects and polymorphic gene correlates of Reward Deficiency Syndrome (RDS). Adv Ther (in press)
Blum K, Chen TJ, Meshkin B ,Downs BW, Gordon CA, Blum SH, Mengucci JF, Braverman ER, Martinez-Pons M (2006) Reward deficiency syndrome in obesity: a preliminary cross-sectional trial with a Genotrim variant. Adv Ther 23, 1040-1051.
Blum K, Chen TJ, Meshkin B, Downs BW, Gordon CA, Blum S, Mangucci JF, Braverman ER, Arcuri V, Deutsch R, Pons MM (2007) Genotrim, a DNA-customized nutrigenomic product, targets genetic factors of obesity: hypothesizing a dopamine-glucose correlation demonstrating reward deficiency syndrome (RDS). Med Hypotheses 68, 844-852.
Blum K, Chen TJH, Meshkin B, Blum SH, Mengucci JF, Notaro A, Arcuri V, Waite RL, Braverman ER (2007) The PPAR- Gamam Pro 12Ala allele polymorphism of the Peroxisome Proliferator -Activated Receptor gene (PPARG2) is a risk factor with a self-identified obese Dutch population. Gene Ther Mol Biol 11, 37-42.
Blum K, Cull JG, Chen TJH, Swan SG, Holder JM, Wood R, Braverman ER, Bucci LR, Trachtenberg MG (1997) Clinical evidence for effectiveness of Phencal™ in maintaining weight loss in an open-label, controlled, 2-year study. Curr Ther Res 58, 745-763.
Blum K, Noble EP, Sheridan PJ, Montgomery A, Ritchie T, Jagadeeswaran P, Nogami H, Briggs AH, Cohn JB (1990) Allelic association of human dopamine D2 receptor gene in alcoholism. JAMA 263, 2055-2060.
Blum K, Trachtenberg MC, Cook DW (1990) Neuronutrient effects on weight loss in carbohydrate bingers: an open clinical trial. Curr Ther Res 48, 217-233.
Chen ALC, Blum K, Chen TJH, Rhinking J, Waite RL, Downs BW, Braverman ER, Arcuri V, Kerner M, Notaro A, Cassel K, Blum SH, Bagchi D, Bagchi M, Robarge A, Kaats G, Comings E. (2008) The impact of biomics technology and
DNA directed anti-obesity targeting of the brain reward circuitry. Gene Therapy and Mol Biol 12, 45-68.
Chen TJH, Blum K, Kaats G, Braverman ER, Eisneberg A, Sherman M, Davis K, Comings DE, Wood R, Pullin D, Arcuri V, Varshavskiy M, Mengucci JF, Blum SH, Downs BW, Meshkin B, Williams L, Schoolfield J, White L (2007a) Chromium Picolinate (CrP) a putative anti-obesity nutrient induces changes in body composition as a function of the Taq 1 dopmine D2 receptor polymorphisms in a randomized double-blind placebo controlled study. Gene Ther Mol Biol 11, 161-170.
Chen TJH, Blum K, Kaats G, Braverman ER, PullinM D, Downd BW, Martinez-Pons M. Blum SH, Mengucci J, Bagchi D, Bagchi M, Robarge A, Meshkin B, Arcuri V, Varshavisky M, Notaro A, Comings DE, White L (2007b) Reviewing the role of putative candidate genes in “Neurobesigenics” aclinical subtype of Reward Deficiency Syndrome (RDS). Gene Ther Mol Biol 11, 61-74.
Comings DE, Gade R, MacMurray JP, Muhleman D, Peters WR (1996) Genetic variants of the human obesity (OB) gene: Association with psychiatric symptoms and body mass index in young women, and interaction with the dopamine D2 receptor (DRD2) gene. Mol Psychiatry 1, 325-335.
Davis C, Levitan RD, Kaplan AS, Carter J, Reid C, Curtis C, Patte K, Hwang R, Kennedy JL (2008) Reward sensitivity and the D2 dopamine receptor gene: A case-control study of binge eating disorder. Prog Neuropsychopharmacol Biol Psychiatry 32, 620-8.
Dong F, Yang X, Sreejayan N, Ren J (2007) Chromium (D-Phenylalanine)3 Improves Obesity-Induced Cardiac Contractile Defect in ob/ob Mice. Obesity (Silver Spring) 15, 2699-2711.
Food And Drug Administration Center For Drug Evaluation and Research (CDER) Joint Meeting of the Nonprescription Drugs and the Endocrinologic & Metabolic Drugs Advisory Committee, January 23, 2006.
Liu D, Zhang Y, Du Y, Yahg G, Zhang X (2007) Identification and characterization of single nucleotide polymorphisms in 6 growth-correlated genes in porcine by denaturing high performance liquid chromatography. DNA Seq 18:220-7.
Loktionov A (2003) Common gene polymorphisms and nutrition: emerging links with pathogenesis of multifactorial chronic diseases (review). J Nutr Biochem 14, 426-451.
Martinez JA, Parra MD, Santos JL, Moreno-Aliaga MJ, Marti A, Martinez-Gonzalez MA (2008) Dependent response to energy-restricted diets in obese subjects: towards personalized nutrition Genotype. Asia Pac J Clin Nutr 17(Suppl 1), 119-22.
Meshkin B, Chen TJH, Chen ALC, Prihoda TJH, Morrisette H, Braverman ER, Blum SH, Cassel K, Williams L, Waite RL, Downs BW, Tung H, Rhoades P, Blum K (2008) Health economics of nutrigenomics in weight management. Gene Ther Mol Biol 12, 25-30.
Nonogaki K, Nozue K, Oka Y (2006) Increased hypothalamic 5-HT2A receptor gene expression and effects of pharmacologic 5-HT2A receptor inactivation in obese Ay mice. Biochem Biophys Res Commun 2351, 1078-1082.
Ordovas JM, Shen J (2008) Gene-environment interactions and susceptibility to metabolic syndrome and other chronic diseases. J Periodontol 79 Suppl 8, 1508-13.
Rance KA, Johnstone AM, Murison S, Duncan JS, Wood SG, Speakman JR (2007) Plasma leptin levels are related to body composition, sex, insulin levels and the A55V polymorphism of the UCP2 gene. Int J Obes (Lond) 31, 1311-1318.
Rosmond R, Chagnon M, Bouchard C (2003) The Pro12Ala PPARγ2 gene missense mutation is associated with obesity and insulin resistance in Swedish middle-aged men. Diabetes Metab Res Rev 19, 159-163.

Blum et al: Exploring Nutrigenomics
382
Sorlí JV, Francés F, González JI, Guillén M, Portolés O, Sabater A, Coltell O, Corella D (2008) Impact of the -1438G>a polymorphism in the serotonin 2A receptor gene on anthropometric profile and obesity risk: a case-control study in a Spanish Mediterranean population. Appetite 50, 260-65.
Stice E, Spoor S, Bohan C, Small DM (2008) Relation between Obesity and blunted striatal response to food is moderated by Taq A1 allele. Science 322, 449-452.
Tiffin N, Okpechi I, Perez-Iratxeta C, Andrade-Navarro MA, Ramesar R (2008) Prioritisation of candidate disease genes for metabolic syndrome by computational analysis of its defining phenotypes. Physiol Genomics, in press.
Trujillo E, Davis C, Milner J (2006) Nutrigenomics, proteomics, metabolomics, and the practice of dietetics. J Am Diet Assoc 6, 403-413.
Volkow ND, Wang GJ, Telang F, Fowler JS, Thanos PK, Logan J, Alexoff D, Ding YS, Wong C, Ma Y, Pradhan K (2008) Low dopamine striatal D2 receptors are associated with prefrontal metabolism in obese subjects: Possible contributing factors. Neuroimage, in press.
Wuschke S, Dahm S, Schmidt C, Joost HG, Al-Hasani H (2007) A meta-analysis of quantitative trait loci associated with body weight and adiposity in mice. Int J Obes (Lond) 31, 829-41.


















