
A role for Glucagon-like peptide-1 in the Pathophysiology ... · Pathophysiology of ... Schematic...
37
1 A role for Glucagon-like peptide-1 in the Pathophysiology of Irritable Bowel Syndrome
Transcript of A role for Glucagon-like peptide-1 in the Pathophysiology ... · Pathophysiology of ... Schematic...

1
A role for
Glucagon-like
peptide-1 in the
Pathophysiology of
Irritable Bowel
Syndrome

2
Contents Non-Standard Abbreviations ............................................................................................. 3
Introduction ......................................................................................................................... 5
The Gastrointestinal Tract ................................................................................................. 5
The Enteric Nervous System (ENS) ................................................................................... 7
What is Irritable Bowel Syndrome? ................................................................................... 8
Irritable Bowel Syndrome Models ..................................................................................... 9
Wistar Kyoto Rat Irritable Bowel Syndrome Model ..................................................... 9
Maternal Separation Rat Irritable Bowel Syndrome Model........................................... 9
Glucagon-like Peptide-1 .................................................................................................. 10
Treating Irritable Bowel Syndrome ................................................................................. 11
The Brain Gut Axis .......................................................................................................... 11
The Hypothalamic-Pituitary-Adrenal Axis .................................................................. 12
Irritable Bowels Syndrome and the Immune System........................................................ 13
Interleukin-6 ................................................................................................................. 14
Aims of the Study .............................................................................................................. 15
Materials and Methods ..................................................................................................... 16
Animals ............................................................................................................................ 16
Tissue Preparation ........................................................................................................... 16
Whole mount preparation of Submucosal Plexus ........................................................ 16
Cross Section Preparation of the Distal Colon ............................................................. 17
Immunofluorescence ........................................................................................................ 17
Enzyme Linked Immunosorbent Assay (ELISA) ............................................................... 19
Statistical Analyses .......................................................................................................... 19
Results ................................................................................................................................. 20
Immunofluorescence labelling of GLP-1Rs in SMP preparations .................................. 20
Colonic Expression of L-cells and GLP-1R and IL-6 expressing cells ........................... 24
Plasma Concentrations of Glucagon-like peptide-1 ........................................................ 25
Discussion ........................................................................................................................... 27
Further Studies ................................................................................................................ 29
References........................................................................................................................... 31

3
Non-Standard Abbreviations
5-HT - 5-hydroxytryptamine
ACTH - adrenocorticotropic hormone
CaCl2 - calcium chloride
cm - centimetres
CRF - corticotropin releasing factor
DPP IV - dipeptidyl peptidase-4
ELISA - enzyme linked immunosorbant assay
ENS - enteric nervous system
FITC - fluorescein isothiocyanate
g - grams
GIT - gastrointestinal tract
GLP-1R - glucagon like peptide-1 receptor
h - hour
HPA - Hypothalamic Pituitary Axis
IBS - Irritable Bowel Syndrome IL-6,8,10,1β - interleukin-6,8,10,1β
KCl - potassium chloride
KH2PO4 - monopotassium phosphate
MgCl2 - magnesium chloride
ml - millilitre
mmol/L - millimoles per litre
MS - maternal separation
NaCl - sodium chloride
NaHCO3 - sodium bicaronate
NaH2PO4 - monosodium phosphate
Na2HPO4 - disodium hydrogen phosphate
PBS - phosphate buffered saline
pM - picomolar
TNF - tumour necrosis factor
µm - micrometres
WKY - Wistar Kyoto

4
Abstract
Introduction: Irritable bowel syndrome (IBS) symptoms include visceral hypersensivity
and altered gut motility. Additionally absorpto-secretory function is altered resulting in
constipation and/or diarrhoea. There is evidence which suggests altered immune cytokine
profiles may contribute to symptom flares. Interleukin-6 (IL-6) has been proposed as a
potential biomarker for IBS as it is elevated in IBS plasma. It also activates submucosal
neurons and stimulates colonic secretion. However other molecules may be important in a
specific characterisation of IBS, post-prandial exacerbated of symptoms. A possible
candidate for this is Glucagon-like peptide-1 (GLP-1), an incretin hormone which is
secreted following a meal and has antispasmodic effects on the gut. GLP-1 receptors are
also expressed in submucosal neurons and modulate neurally evoked chloride secretions.
Therefore the role of GLP-1 and the potential cross-talk between GLP-1 and IL-6 was
investigated further.
Methods: Whole mount preparations of the submucosal plexus from male Sprague
Dawley (SD) rats were fixed and stained with GLP-1R and dual labelled with IL-6,
calbindin, calretinin, nNOS, S100, Synapsin I or PSD-95. Cross sections were also
prepared from adult male SD and Wistar Kyoto rats and dual labelled with GLP1 and
GLP-1R or IL-6. GLP-1 levels were determined using an ELISA and results were
correlated with previously determined levels of circulating IL-6, TNF-α, IL-1β and IL-8.
Results: In immunohistochemistry studies GLP-1R were expressed in a punctate manner
in the neuronal fibres, cell bodies and glial cells. 26% (n=9/35) of calbindin neurons, 21%
(n=18/84) of calretinin neurons, 37.5% (n=48/128) of S100 cells and the majority, 63%
(n=27/43) of nNOS were GLP-1R positive. Dual labelling with GLP-1R and Synapsin I or
PSD-95 showed that GLP-1R is present in the synapses. GLP-1R co-localised with 30%
(n=26/87) of IL-6 positive neurons revealing potential crosstalk between the molecules. In
Elisa studies (n=9 samples per group) GLP-1 levels were significantly elevated in IBS-A
(p<0.05) plasma and there was a trend towards significance in IBS-D plasma samples.
Moreover plasma GLP-1 levels correlated with IL-6 levels.
Conclusion: These data demonstrate that GLP-1Rs are expressed on colonic submucosal
neurons, where GLP-1 induced cellular signalling is likely to be mediated by nitrergic
mechanisms. Moreover many of these cells co-express IL-6 revealing the potential cross-
talk between IL-6 and GLP-1. Correlation between IL-6 and GLP-1 levels in human IBS
plasma implicates this incretin hormone in the pathogenesis of IBS.

5
Introduction
The Gastrointestinal Tract
Fig 1:Schematic diagram of the gastrointestinal tract (Tharakan et al., 2010)
The gastrointestinal tract (GIT) provides the body with a continual supply of water,
nutrients and electrolytes. Food is propelled through the tract, where it is digested by
secreted digestive juices facilitating nutrient and water absorption. Regulation of this
fundamental process is both by extrinsic and intrinsic neuronal control and through
endocrine and paracrine processes. Figure 1 shows the various parts to the GIT and
digestive system. Each part of the GIT is adapted for specific functions: the oesophagus is
needed for the passage of food, the stomach functions as a temporary storage of food and
also digests food, the small intestine is needed to digest and absorb food and the large
intestine is needed for the absorption of water and electrolytes (Guyton and Hall, 2001).

6
The large intestine consists of the cecum, colon, rectum and anal canal. The colon can be
divided into four parts; the ascending colon, the transverse colon, the descending colon and
the sigmoid colon. The ascending colon and proximal transverse colon are known as the
proximal colon and the distal transverse, descending and sigmoid colon are known as the
distal colon. The distal colon is the anatomical part under investigation in this study. The
wall of the colon as well as most of the GIT, typically includes, from outer to inner layer;
the serosa, a longitudinal muscle layer, a circular muscle layer, the submucosa and the
mucosal layer. As figure 2 shows, the mucosal lining of the large intestine consists of
simple columnar epithelium that has numerous crypts, known as crypts of Lieberkuhn.
These crypts contain three cell types; absorptive, goblet and granular, with goblet cells as
predominating cell type (Seeley et al., 2003).
Fig 2: Histology of the large intestine (Martini et al., 2012)
Goblet cells of the colon are responsible for the mucosal excretions. The function of the
mucous is to lubricate the wall of the lumen and to help the faecal matter stick together as
well as protecting the intestinal wall from bacterial activity that occurs within the faeces.
The mucous contains bicarbonate ions which is responsible for protecting the intestinal
wall from acids formed (Guyton and Hall, 2001). The colonic absorption of water occurs
by a molecular pump which exchanges HCO3- for Cl- in epithelial cells in response to acid

7
produced by colic bacteria. Also another pump exchanges Na+ for H+, water then crosses
the colon by osmosis due to the NaCl gradient.
Irritation to the wall of the colon causes an increase in mucous secretion by triggering local
enteric reflexes. The submucosal plexus and myenteric plexus are both part of the enteric
nervous system (ENS). As well as the enteric nervous system, the GIT is supplied with
parasympathetic and sympathetic innervations (Seeley et al., 2003). The submucosal
plexus which was under investigation in this study in relation to Irritable Bowel Syndrome
(IBS) only has parasympathetic input and controls mainly the secretion and absorption
functions. The myenteric plexus is responsible for contractility of the GIT and has both
sympathetic and parasympathetic input (Guyton and Hall, 2001).
The Enteric Nervous System (ENS)
The ENS is one of the main divisions of the autonomic nervous system and governs the
function of the gastrointestinal tract. The ENS is collected into two types of ganglia the
myenteric (Auerbach’s) plexus and the submucosal (Meissner’s) plexus. The myenteric
plexus is contained between the longitudinal and circular muscle layer. This plexus
contains many interconnecting neurons that extend the entire GIT. When this plexus is
stimulated there is increased tonic contraction of the gut wall, increased intensity of
rhythmical contractions, an increase in the rhythm of the contraction and increased velocity
of conduction of excitatory waves. The myenteric plexus has excitatory as well as
inhibitory neurons. The inhibitory neurotransmitter of this plexus is possibly vasoactive
intestinal polypeptide.
The submucosal plexus is contained within the submucosa. It controls the function within
each minute segment of the intestine. The plexus controls local intestinal secretion, local
absorption and local contraction of the submucosal muscle as the sensory signals that
originate in the epithelium are integrated into the submucosal plexus. The local contraction
causes various degrees of infoldings of the mucosa.
The different neurons within the submucosal and myenteric plexus release different types
of neurotransmitters. The neurotransmitters include acetylcholine, norepinephrine,

8
adenosine triphosphate, serotonin, dopamine, cholecystokinin, substance P, vasoactive
intestinal polypeptide, somatostatin, leu-enkephalin, met-enkephelin and bombesin
(Guyton and Hall, 2001). A colonic mucosal biopsy would contain the above mentioned
neurotransmitters as well as proteases and histamine. An increased activation of
submucosal neurons is seen in the presence of colonic mucosal biopsy supernatants from
Irritable Bowel Syndrome (IBS) patients (Buhner et al., 2012). Therefore it is important to
study the submucosal plexus further in relation to IBS.
What is Irritable Bowel Syndrome?
IBS is a functional gastrointestinal disorder. Symptoms include abdominal pain, discomfort
and altered bowel habits (Yale et al., 2008), affecting approximately 10% of the population
(Corsetti et al., 2004). IBS is a heterogeneous, multisystem, multidomain disorder as
opposed to a single disease entity (Crentsil, 2005) with several mechanisms being
proposed to explain the altered sensory responses observed (Yale et al., 2008). It is also a
polygenic disorder and the IBS phenotype can be determined by many common genetic
variants (Saito et al., 2010). To evaluate the role of genes in IBS, candidate genes are
studied that have an association with the intermediate phenotypes of IBS such as altered
colonic transit, colonic motility and compliance and decreased sensation thresholds and
ratings. Some of the candidate genes include genes involved in the alteration of
serotonergic mechanisms (Camilleri and Katzka, 2012).Studying genes involved in
serotonergic mechanisms show polymorphisms in the promoter region of serotonin
transporters in IBS (Murphy et al., 2004) .
Genetic abnormalities along with abnormalities in the enteric and the central nervous
system which lead to altered pain and motor functions are all thought to account for the
symptoms of IBS (Drossman, 2005). Also psychological stressors and ongoing enteric
infections have been implicated in the onset and maintenance of IBS (FitzGerald et al.,
2009). Indeed 7-31% of patients develop IBS after gastroenteritis (McKendrick, 1996) and
this is termed post infectious IBS, thus revealing the importance of immune activation in
this disorder.

9
As there is no biochemical marker or structural abnormality for IBS, diagnosis is based on
the presence of clinical symptoms according to Rome III criteria (Dorn et al., 2009).
However groups can be either distinguished as being IBS-D (diarrhoea predominant), IBS-
C (constipation predominant) or IBS-A (alternating between diarrhoea and constipation).
There is also a variability of symptoms between male and females and within each group
(Yale et al., 2008).
Irritable Bowel Syndrome Models
Wistar Kyoto Rat Irritable Bowel Syndrome Model
The Wistar Kyoto rat is a pre-clinical model of IBS. IBS is associated with an enhanced
visceral sensitivity in response to altered brain gut axis signalling such as that occurs
during stress. The WKY rats are a viscerally hypersensitive rat model of IBS as they are
genetically predisposed to pathological stress responses with alterations in colonic
accommodation and morphology (Martínez et al., 2007) which leads to enhanced colonic
motility and faecal output following a psychological stressor (O'Malley et al., 2010). Also
following stressful stimuli WKY rats display exaggerated corticosterone levels (O'Mahony
et al., 2010). A study has also shown that while the plasma Interleukin-6 (IL-6) levels are
similar, mucosal scrapings from WKY rats contain higher levels of IL-6 and excised
colons secrete more IL-6 than Sprague Dawley (SD) colons. The WKY secretions have
also been shown to excite submucosal neurons more so than SD secretions (O'Malley et
al., 2011a). O’Malley et al. also observed that under resting conditions the WKY rat in
comparison to the SD rat displays a pro-absorptive phenotype as was elicited by the
decrease in secretory currents. However this reversed in the presence of IL-6 where WKY
colonic tissues evoked a larger secretory current (O'Malley et al., 2012).
Maternal Separation Rat Irritable Bowel Syndrome Model
The maternal separation (MS) rat model is another model of IBS. This model involves a
postnatal stress that induces alterations in the hypothalamic-pituitary-adrenal (HPA) axis of
the adult offspring (O'Malley et al., 2010). Like the WKY rat model the MS model exhibit
symptoms of anxiety and depression (O'Mahony et al., 2008). The MS rat also exhibits

10
exaggerated GI response to inflammatory stimuli and colorectal distension, showing
increased stress induced faecal output. This model shows an increased level of circulating
IL-6 following an immunological challenge as well as altered 5-HT levels, and visceral
hypersensitivity (O'Mahony et al., 2009).
The two rat models exhibit alterations in colonic motility (O' Mahony et al., 2011) which is
a prominent phenotype of IBS (Camilleri and Katzka, 2012). As gut motility is a major
function of glucagon-like peptide-1 studying GLP-1 in relation to IBS is of clinical interest
(Hellström, 2009).
Glucagon-like Peptide-1
GLP-1 is formed in the L cells following cleavage from a pro-glucagon precursor molecule
by post-translational processing and is a C-terminally amidated 7-37 amino acid peptide
(Hellström, 2011). GLP-1 regulates numerous gastrointestinal functions, it decreases
gastric acid secretions (Schjoldager et al., 1989), gastrointestinal transit, motility
(Hellström et al., 2008) and gastric wall tone (Schirra et al., 2002).
The MMC are waves of activity that sweep through the intestine during the fasting state
and they help trigger peristaltic waves which facilitate transport of the bolus through the
GIT (Tolessa et al., 1998b). Helstrom et al. showed that GLP-1 inhibits the migrating
myoelectric complex (MMC) during fasting in healthy and IBS patients (Hellström et al.,
2008).
Along with regulating gastrointestinal motility functions GLP-1 has anti-hyperglycaemic
actions. GLP-1 is released from L-cells in the ileum and colon following intake of
carbohydrates and fat and is considered an important regulator of postprandial insulin
secretion from the pancreas in response to a meal (Hellström et al., 2008). GLP-1 acts on
β-cells of the pancreas to cause an increase in insulin secretion even before blood sugar
levels become elevated (Hellström, 2009). GLP-1 has also been shown to suppress
glucagon release from the pancreas and to enhance the sensitivity of pancreatic B-cells to
glucose (Gutniak et al., 1994).

11
GLP-1 has a half-life of about 1-2 minute and is degraded by depiptidyl peptidase IV
(DPP- IV) which is an enzyme present in the plasma (Hellström, 2011). The fact that it has
such a short half-life together with the fact that GLP-1 has no effects on intestinal muscle
strips in vitro concludes that direct endocrine or autocrine actions are unlikely. Therefore
neurocrine actions of GLP-1 are most likely (Tolessa et al., 1998a).
GLP-1 has also been has also been shown to be implicated in stress induced alteration in
colonic transit via central CRF pathways and peripheral cholinergic pathways in rats
(Nakade et al., 2007). IBS symptoms are exacerbated following a meal as lipids in the
intraduodenum are shown to increase visceral sensitivity (Caldarella et al., 2005) and as
GLP-1 is released following intake of carbohydrates and fat (Hellström et al., 2008) it is
important to investigate GLP-1 in relation to IBS.
Treating Irritable Bowel Syndrome Despite the prevalance and economic burden of IBS treatment options remain limited.
Current treatment options include dietary adjustment, for example increasing the amount of
fiber in the diet, the use of antidiarroheals and the use of laxatives (Dalrymple and Bullock,
2008). IBS pain is due to visceral hypersensitivity and patients who have this pain as their
most common complaint are most impacted by IBS and and there are currently no
treatments targeting this pain (Olden, 2003).
As previously described GLP-1 plays a role in the regulation of gut motility and therefore
could be important clinically in treating pain that is associated with abnormal motor
activity in the gut. A clinical trial was formulated where the GLP-1 hormone analogue
ROSE-010 was administered to suitable IBS patients. The results found that the GLP-1
analogue ROSE-010 caused meaningful pain relieve in a significant proportion of the
patients and delayed gastric emptying of food without retarding small bowel or colonic
transit (Hellström et al., 2009).
The Brain Gut Axis The Brain gut axis includes the parasympathetic innervation and the sympathetic
innervation, which supplies nerve fibres from the brain and spinal cord to the GIT. The
brain gut axis also includes the previously described enteric nervous system (Saulnier et
12
al., 2013). A system which has a major influence on the brain gut axis is the HPA axis
(Dinan, 1994).
The Hypothalamic-Pituitary-Adrenal Axis
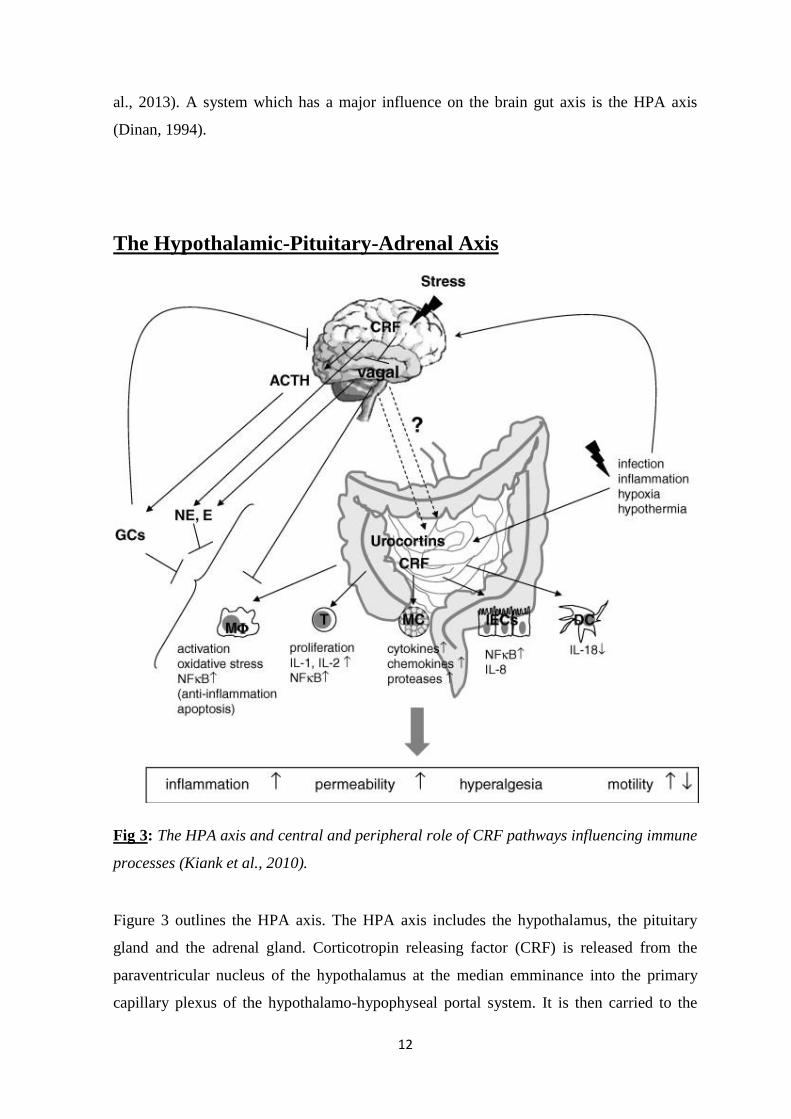
Fig 3: The HPA axis and central and peripheral role of CRF pathways influencing immune
processes (Kiank et al., 2010).
Figure 3 outlines the HPA axis. The HPA axis includes the hypothalamus, the pituitary
gland and the adrenal gland. Corticotropin releasing factor (CRF) is released from the
paraventricular nucleus of the hypothalamus at the median emminance into the primary
capillary plexus of the hypothalamo-hypophyseal portal system. It is then carried to the

13
anterior lobe of the pituitary where it stimulates corticotropes to release
adrenocorticotropic hormone (ACTH). ACTH stimulates the synthesis of cortisol,
glucocorticoids and mineralocorticoids (Gillespie and Nemeroff, 2005). CRF containing
neurons are widely distributed throughout extrahypothalmic brain areas and acts as a
neuromodulatory agent in coordinating behaviour, endocrine, autocrine and immune
responses to stress (Corsetti et al., 2004). CRF release is also controlled by
neurotransmitters such as 5-HT and norepinephrine in response to stress (Dinan, 1996)
Patients with IBS show an exaggerated stress response when administering CRF and this
could be seen by the enhanced levels of ACTH and cortisol (Dinan et al., 2006, FitzGerald
et al., 2009). The increase in ACTH could be due to the exogenous CRF acting
synergically with the pro-inflammatory cytokines and other secretogogues to bring about
ACTH release. The increase could also be due to the CRF1 receptor on the anterior
pituitary being up-regulated in IBS (Gillespie and Nemeroff, 2005). The enhanced level of
ACTH and pro-inflammatory cytokines were shown not to be due to a decreased
sensitivity of the glucocorticoid receptor (Dinan et al., 2006). There is also evidence of a
similar upregulation of CRF1 receptor in the gut as a result of stress. This upregulation of
CRF1 receptors warrants further investigation of CRF1 receptor antagonists as a treatment
option for IBS (Fukudo et al., 1998, Gillespie and Nemeroff, 2005).
Irritable Bowels Syndrome and the Immune System
It is well established that acute gastrointestinal infections initiate the onset of IBS
symptoms in at least a sub-group of patients (McKendrick and Read, 1994). While some
IBS patients show persistent levels of low grade inflammation with increased levels of pro-
inflammatory cytokines and activated T lymphocytes and mast cells (van der Veek et al.,
2005). It is also shown that IBS patients have altered GI permeability (Camilleri et al.,
2012) and most pro-inflammatory cytokines have the capacity to alter intestinal epithelial
permeability (Al-Sadi et al., 2009).
Figure 3 outlines CRF pathways and the influence they have on immune processes.
Immune cells such as macrophages, T lymphocytes, mucosal mast cells, and dendritic cells

14
respond to CRF receptor signalling mainly by promoting inflammation. However central
CRF evoke an anti-inflammatory response, which may counter-regulate inflammatory
responses in the gut, by increasing glucocorticoids and catecholamines and decreasing
parasympathetic activity (Kiank et al., 2010). In response to infection CRF containing
neurons also respond to pro-inflammatory cytokines (Chrousos, 1995).
Cytokines are small cell signalling protein molecules secreted by leukocytes and various
other cells. Fundamentally cytokines influence innate and adaptive immunity (Ortiz-Lucas
et al., 2010). Specifically pro-inflammatory cytokines such as IL-6 and IL-8 are shown to
be increased in patients with IBS and are shown to activate the HPA axis (Dinan et al.,
2006, FitzGerald et al., 2009). Analysis has also shown that HPA hyperresponsivity is
related to enhanced IL-6 levels (Dinan et al., 2006). In turn activating the HPA axis causes
an increased release of glucocorticoids which depresses inflammatory processes of the gut
(Turnbull and Rivier, 1999). It is unclear whether the increase in IL-6 and IL-8 are
biological markers for a trait of the syndrome or IBS itself (Dinan et al., 2006).
Interleukin-6
Interleukin-6 (IL-6) is a pro-inflammatory and anti-inflammatory cytokine (Scheller et al.,
2011) and stimulates secretion into the colonic lumen by modulating submucosal neurons.
IL-6 is elevated in the mucosa and plasma of the pre-clinical model of IBS, the WKY rat
(O'Malley et al., 2012, O'Malley et al., 2011a). IL-6 levels are also increased in the plasma
of IBS patients which may reflect an important mechanism which leads to symptom flares
(Dinan et al., 2006).
Following administration of cholinesterase inhibitor there is an increase in IL-6 synthesis
which correlates to an increase in abdominal bloating (Dinan et al., 2008). IL-6 along with
IL-1 β was shown to act as excitatory neuromodulators of myenteric neurons via
presynaptic inhibition by inhibiting acetylcholine release from pre-synaptic neurons
(Keller et al., 2001). It has also been shown that IL-6 inhibits nicotinic and noradrenergic
neurotransmitter release in guinea pig submucosal neurons (Xia et al., 1999).IL-6 receptors
are expressed on rat submucosal neurons and following exposure to recombinant IL-6 there
is an increase in extracellular calcium and colonic excretion that overrides the pro-

15
absorptive phenotype, which is regulated by epithelial cholinergic activity at rest
(O'Malley et al., 2011b).
Adipose tissue is a major source of IL-6 concentration and therefore contributes to the
induction of insulin resistance. Contracting skeletal muscles during exercise also increases
circulating IL-6 concentrations. It is also proposed to promote nutrient availability and
improves whole body insulin sensitivity. Pancreatic alpha cells are also a primary target of
IL-6 action. IL-6 promotes alpha cell proliferation and inhibits apoptosis. In a high fat diet
alpha cells mass expands in an IL 6 dependent manner. IL-6 was also shown to promote
GLP-1 secretion and production from intestinal L cells and pancreatic alpha cells
(Ellingsgaard et al., 2011)
Other cytokines such as interferon γ, tumour necrosis factor (TNF), transforming growth
factor and interleukin-10 (IL-10) regulate inflammatory and immune responses
(Gonsalkorale et al., 2003a). A UK study showed that frequency of the high IL-10
producer genotype for IL-10 (an anti-inflammatory cytokine) were significantly reduced in
patients with IBS as compared with healthy controls (Gonsalkorale et al., 2003b).
Aims of the Study
The aims of the study were to indentify the location of GLP-1R and IL-6 within the
submucosal ganglia of the distal colon. After getting location of GLP-1R and IL-6 within
the submucosal ganglia we wanted to observe the quantity and location of GLP-1, IL-6 and
GLP-1R across the entire cross section of the distal colon. To relate this to the human
condition we investigated levels of GLP-1 in IBS patients and compared them to healthy
controls and aimed to correlate these values with levels of other pro-inflammatory
cytokines.

16
Materials and Methods
Animals Male Sprague Dawley rats were purchased from Harlan UK weighing approximately 250-
350 g. The rats were housed in groups of four to six per cage. They were maintained on a
12/12 h dark–light cycle and were given food and water ad libitum and kept at a room
temperature of 21-23oC. Principles of laboratory animal care were followed, and all
procedures were carried out in accordance with EU directive 89/609/EEC and approved by
the Animal Experimentation and Ethics Committee.
Tissue Preparation
Whole mount preparation of the Submucosal Plexus
A section of the colon was excised from each rat, cut 8cm proximal from the anus. The
excised distal colon was stored in ice cold Krebs (pH7.4) solution at 4oC; containing in
mmol/L: 117 NaCl, 4.8 KCl, 2.5 CaCl2, 1.2 MgCl2, 25 NaHCO3, 1.2 NaH2PO4 and 11 D-
glucose. The pellets were then expunged using a 10 ml syringe filled with Krebs. Using a
forceps the section of colon was pulled over a glass rod allowing for the mesenteric blood
vessels to be removed. The colon was lightly scored along where the mesentery was
removed, using a blade.
Fig4: Layers of the colon. JD Wood et al
Figure 4 shows the various layers of the colon. The longitudinal and circular muscle
containing the myenteric plexus were removed using a cotton swab. A scissors was used to

17
cut all the way through the mesentery and the remaining submucosal layer was pinned out
mucosal side up on a sylgard lined plate filled with Krebs saline. Figure 4 also shows that
the submucosal ganglion is contained within a separate layer from the mucosa. Using
forceps the mucosal layer was removed and the remaining submucosal plexus was
prepared for Immunofluorescent Imaging as detailed below.
Cross Section Preparation of the Distal Colon
Sections of colon from four separate Sprague Dawley rats (the control model) and four
separate Wistar Kyoto rats (the pre-clinical IBS model) had been fixed and frozen from a
previous study. Short sections of the colons were cut and brought up to -250C in the
cryostat (Leica, Nussloch, Germany; CM1900 UV). Using the cryostat, sections were
prepared. Firstly Tissue-Tek® O.C.T. (optimum cutting temperature) compound
embedding medium was applied to the stage and left to freeze for ten minutes. The colon
was then mounted vertically onto the stage and more mounting medium was applied over
the colon, again the medium was left to freeze for another 10 minutes. 10 μm sections were
taken with 30 μm between sections. Eight sections were taken from each colon and applied
to two separate Fisherbrand glass slides, four sections were applied to a slide to be stained
for anti-GLP-1R and anti-GLP-1 and four sections were applied to a slide to be stained
with anti-IL-6 and anti-GLP-1.
Immunofluorescence
Immunofluorescence of whole mount SMP preparations
Using 96-well plates seven separate sections of the submucosal plexus were stained from
each of the four Sprague Dawley rats. The whole mount preparations were first fixed
overnight in Zamboni’s fixative [paraformaldehyde (from 16% stock solution), picric acid
(saturated aqueous), 10M NaOH, 0.1M phostphate buffer] at 4oC. The sections were
washed three times for 15 minutes in phosphate buffered saline (PBS) (containing in
mmol/L: 137 NaCl, 2.7 KCl, 10 Na2HPO4.2H2O, 2.0 KH2PO4) and agitated. The
preparations were then permeabilised for 1 hour in 0.1% Triton X-100 in PBS at room
temperature (R.T.). Again the sections were washed three times (X3) for 15 minutes in
PBS. For 1 hour the sections were incubated in blocking solution (1% donkey serum in
PBS) at R.T. All sections were incubated overnight at 4oC with a 1:250 dilution of anti-

18
GLP-1 receptor (GLP-1R) antibody (affinity purified rabbit polyclonal antibody a synthetic
peptide conjugated to KLH derived from within 250-350 of Human GLP1R; Abcam,
Cambridge, UK). The sections were washed again in PBS for 15 minutes X3. The GLP-1R
antibody labelled tissues, were incubated with a 1:250 dilution of Texas Red conjugated
donkey anti-rabbit secondary antibody (Jackson ImmunoResearch, West Grove, PA,
USA). The sections were washed for 10 minutes in PBS X3. For dual labelling the sections
were incubated overnight with a separate primary antibody including neuronal markers for
calbindin (1:300; mouse, Swant, Bellinzona, Switzerland) calretinin (1:300, goat; Swant)
and neuronal nitric oxide synthase (nNOS, 1:300; goat, Abcam), the glial cell marker S100
(1:300, mouse; Sigma-Aldrich, St. Louis, MO) and the anti-rat IL-6 antibody (affinity
purified goat antibody immunised E.coli-derived rIL-6, 1:300; R&D Systems, Abingdon,
UK). Sections were also stained with the pre-synaptic marker Synapsin I (1:250, goat;
Santa Cruz Biotechnology, Santa Cruz, CA) and the post-synaptic marker PSD-95 (1:200;
mouse, Santa Cruz Biotechnology). Slides were again washed for 10 minutes in PBS X3.
The sections were then incubated for 2 hours with Fluorescein isothiocyanate (FITC)
conjugated antibody (1:250 goat anti mouse Jackson ImmunoResearch) and (1:250 donkey
anti-goat; Milipore). Dako fluorescent mounting medium was used to mount the sections
onto glass slides and a cover slip was placed over them.
Immunofluorescence of Cross Sections of Colon
The cross sections were prepared for immunofluorescent imaging. Two slides containing
four sections were prepared for each rat. One of the two slides from each rat was incubated
with a 1:200 dilution of GLP-1 antibody (mouse antibody; Antibody Shop). A 1:300
dilution goat anti-mouse rhodamine conjugated secondary antibody was used as a
fluorophore (Jackson ImmunoResearch). This slide was then dual labelled with a 1:250
dilution of GLP-1R antibody (Abcam) with FITC conjugated donkey anti-rabbit secondary
antibody as the fluorophore. The second slide from each rat was incubated with a 1:200
dilution of GLP-1 antibody and with a 1:300 dilution of the anti-mouse rhodamine
conjugated secondary antibody. The second slide was also incubated with the anti-rat IL-6
antibody (1:300; R&D Systems) and with a FITC conjugated anti-goat secondary acting as
the fluorophore.

19
Image analyses
Neurons in submucosal ganglia were imaged using Cell-F software (Soft Imaging
Solutions) and an Olympus BX51 fluorescent microscope [Olympus America, Inc,
Melville, NY, USA] with an Olympus DP71 digital camera [Optronics, Goleta, CA,USA]
with filter sets for TRITC [excitation (557nm), emission (576nm)] and FITC [excitation
(470nm), emission (525nm)]. Images were also obtained from the FLUOVIEW FVlOi-
Olympus-confocal. Three ganglia per tissue were imaged from four separate rats. With the
Cell-F software co-localisation between the cell markers (calbindin, calretinin, nNOS and
S100) and GLP-1R was determined by counting the number of cells positive for the cell
markers and calculating the percentage of these cells that co-localised with GLP-1R. Co-
localisation between the synaptic markers (PSD-95 and Synapsin I) and GLP-1R was
determined using the confocal. Co-localisation between and location of GLP-1R and GLP-
1 was also determined using confocal microscopy as was co-localisation between and
location of GLP-1 and IL6 on the colon cross sections.
Enzyme Linked Immunosorbent Assay (ELISA)
A sandwich ELISA was carried out to determine the GLP-1 concentration in plasma
samples from 9 healthy, 9 IBS-C, 9 IBS-D and 9 IBS-A patients following manufacturer’s
guidelines (Milipore; Cat. EGLP-35K). The assay was run in duplicate. DPP-IV
(dipeptidyl peptidase-IV) was added to the samples to inhibit GLP-1 degradation. A multi-
pipette was used to ensure accurate measurements of solutions were made. The plates were
read on a Synergy HT fluorescent microplate reader with an excitation/emission
wavelength of 355 nm and 460 nm respectively and the lowest level of GLP-1 that could
be detected was 2 pM. Having solutions of known GLP-1 concentrations, from 0 to 100
pM, a standard curve was created. From this curve the GLP-1 concentrations of the plasma
samples were determined.
Statistical Analyses
Experiments were conducted in at least four different animals and in the case of the ELISA
at least 9 patients per group. ELISA data was analysed using GraphPad prism for windows
(version 7). Paired or unpaired two-tailed Student’s t-test were used where appropriate,
P<0.05 was considered significant.

20
Results
Immunofluorescence labelling of GLP-1Rs in SMP
preparations
Immunohistochemistry techniques were carried out on whole mount preparations taken
from the distal colon of male, adult SD rats. To determine the neuronal cell type that
expressed GLP-1R co-localisation, dual labelling studies were conducted with calbindin
antibodies to label cholinergic sensory neurons, anti-calretinin for cholinergic motor
neurons and anti-nNOS for nitrergic neurons. Anti-S100 labelled glial cells in the
submucosal ganglia.
In Figure 5 the images show GLP-1R (red stain) dual labelled with calbindin (Fig 5(a)
n=9/35), calretinin (Fig 5(b) n=18/84), nNOS (Fig 5(c) n=27/43) and S100 (Fig 5(c)
n=48/128) (green stain), (Fig 5; n= 3 ganglia per tissue from 4 different rats). GLP-1R
expression is found in the cell bodies but is also prominent in the neuron fibres and glial
cells. The prominent staining in the neuronal cell fibres is also quite punctate indicating
that GLP-1Rs are clustered together.
In Figure 7 it shows the percentage of total neurons that are positive for the different cell
markers which co-localises with GLP1R. Dual labelling with GLP-1R and calbindin
antibodies show a 26% (n=9/35) co-localisation in the cell bodies of neurons. Dual
labelling with GLP-1R and calretinin shows a 21% (n=18/84) co-localisation in the cell
body of neurons. Dual labelling with GLP-1R and nNOS shows a 63% (n=27/43) co-
localisation in the cell body of neurons. There was also GLP-1R staining of glial cells as
dual labelling with GLP-1R and S100 shows a 37.5% (n=48/128) co-localisation with the
glial cells of neurons.

21
Fig 5(a): GLP1R (Texas Red) Fig 5(b): GLP1R (Texas Red) and nNOS
and Calbindin (FITC), (FITC)
Fig 5(c): GLP1R (Texas Red) Fig 5(d): GLP1R (Texas Red) and
and calretinin (FITC) and S100 (FITC)
Fig 5: Scale bars= 20 μm
The punctate nature of the GLP1-R staining suggested that the receptor was potentially
expressed at synapses. To investigate this further, tissues were dual labelled with GLP1-R
antibody and the presynaptic marker Synapsin I and also dual labelled with GLP-1R
antibody and the postsynaptic marker PSD-95. GLP-1R expression (red stain) and
Synapsin I or PSD-95 (green stain) dual labelling can be seen in the confocal images in
Figure 6. Some co-localisation was seen between GLP-1R and the presynaptic marker
Synapsin I (Fig 6(a)) and the postsynaptic marker PSD-95 (fig 6(b)).
Cell Body

22
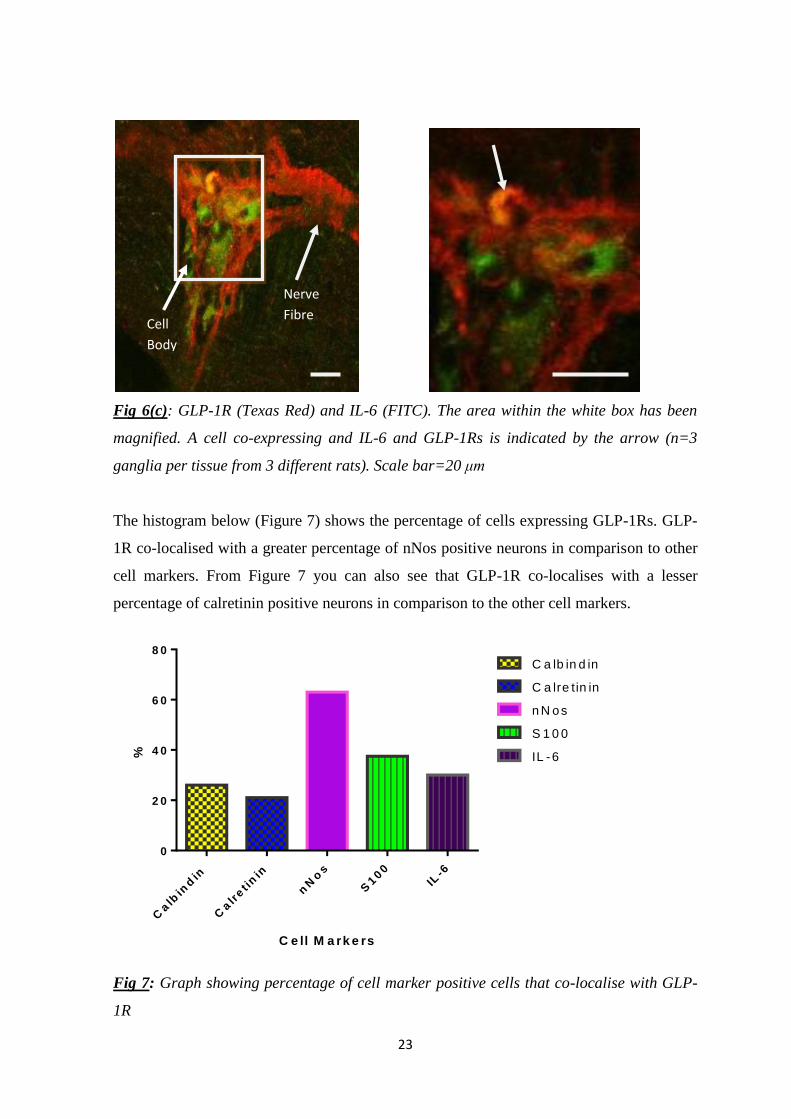
To determine cross-talk, co-localisation studies were conducted between IL-6 and GLP-
1R. Dual labelling with GLP-1R and IL-6, shows that co-localisation is present in 30%
(n=26/87) of neuronal cell bodies. In Figure 6(c) one neuronal body (indicated by arrow)
shows co-localisation between GLP-1R and IL-6. GLP-1R is the red stain and IL-6 which
is conjugated to the FITC secondary antibody is the green stain.
Fig 6(a): GLP1R (Texas Red) and Synapsin I (FITC). The area within the white box has
been magnified. Regions of co-localisation (yellow staining) are indicated by the arrows.
Scale bar=20 μm
Fig 6(b): GLP-1R (Texas Red) and PSD-95 (FITC). The area within the white box has
been magnified. Regions of co-localisation (yellow staining) are indicated by the arrows.
Scale bar =20 μm
23
Fig 6(c): GLP-1R (Texas Red) and IL-6 (FITC). The area within the white box has been
magnified. A cell co-expressing and IL-6 and GLP-1Rs is indicated by the arrow (n=3
ganglia per tissue from 3 different rats). Scale bar=20 μm
The histogram below (Figure 7) shows the percentage of cells expressing GLP-1Rs. GLP-
1R co-localised with a greater percentage of nNos positive neurons in comparison to other
cell markers. From Figure 7 you can also see that GLP-1R co-localises with a lesser
percentage of calretinin positive neurons in comparison to the other cell markers.
C e ll M a rk e rs
%
Calb
ind
in
Calr
et i
nin
nN
os
S100
IL-6
0
2 0
4 0
6 0
8 0
C a lb in d in
C a lre tin in
n N os
S 1 0 0
IL -6
Fig 7: Graph showing percentage of cell marker positive cells that co-localise with GLP-
1R
Nerve
Fibre Cell
Body

24
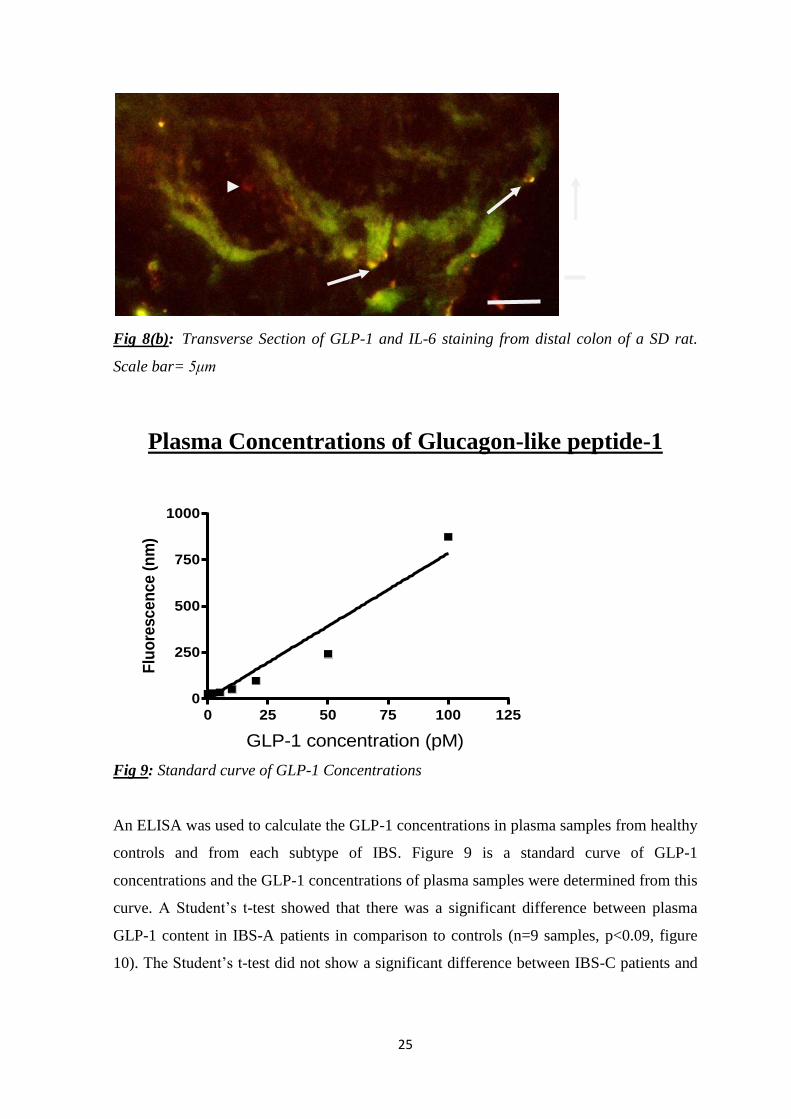
Colonic Expression of L-cells and GLP-1R and IL-6
expressing cells To determine whether GLP-1 secreting L-cells were in close proximity to GLP-1R
expressing submucosal neurons transverse sections were immunolabelled with GLP-1 and
GLP-1R and sections were also labelled with GLP-1 and IL-6 antibody. The
immunohistochemistry techniques were also used to compare the GLP-1, IL-6 and GLP-
1R staining between the IBS pre-clinical model the WKY rat, which has altered GI
morphology and the control model the SD rat. Co-localisation can be seen in Figure 8(a).
The co-localisation is between GLP-1 (red staining) and GLP-1 receptor (green staining)
and shows a couple of L-cells (indicated by arrows) in the mucosal layer of the WKY rat.
The co-localisation between IL-6 (green staining) and GLP-1 (red staining) can be seen in
Figure 8(b). A GLP-1 expressing L-cell that does not co-localise with IL-6 staining is
indicated by the arrowhead and two L-cells co-expressing GLP-1 and IL-6 are indicated by
the arrows.
Fig 8(a): Transverse section of GLP-1 and GLP-1R staining from distal colon of a WKY
rat. Scale bar= 5μm
25
Fig 8(b): Transverse Section of GLP-1 and IL-6 staining from distal colon of a SD rat.
Scale bar= 5μm
Plasma Concentrations of Glucagon-like peptide-1
Fig 9: Standard curve of GLP-1 Concentrations
An ELISA was used to calculate the GLP-1 concentrations in plasma samples from healthy
controls and from each subtype of IBS. Figure 9 is a standard curve of GLP-1
concentrations and the GLP-1 concentrations of plasma samples were determined from this
curve. A Student’s t-test showed that there was a significant difference between plasma
GLP-1 content in IBS-A patients in comparison to controls (n=9 samples, p<0.09, figure
10). The Student’s t-test did not show a significant difference between IBS-C patients and
0 25 50 75 100 1250
250
500
750
1000
Flu
ore
scen
ce (
nm
)
GLP-1 concentration (pM)

26
healthy controls (n=9 samples, p>0.05), however there was a trend toward a significant
difference between IBS-D patients and healthy patients (n=9 samples, p=0.09, figure 10).
Fig 10: GLP-1 concentrations (pM) in healthy controls, and IBS patients. A significant
elevation in GLP-1 levels in IBS-A (n=9) in comparison to healthy controls (p<0.05, t-
test).
Interestingly when compared to previously published data on the levels of cytokines in IBS
plasma (McKernan et al., 2011) a significant correlation was found between the levels of
IL-6 and GLP-1 in healthy controls, IBS-C, IBS-D and IBS-A (r2=0.12, p<0.05, Figure
11). No correlation was detected between GLP-1 and IL-1β, TNF-α or IL-8.
Linear regresson GLP-1 IL-6
6.5 7.0 7.5 8.0 8.50
1
2
3
4
5
GLP-1 (pg/ml)
IL-6
(p
g/m
l)
Figure 11: Linear regression between levels of IL-6 and GLP-1 in healthy controls, IBS-C,
IBS-D and IBS-A.
Healthy IBS-C IBS-D IBS-A0
2
4
6G
LP
-1 (
pM
)
N=9 N=9 N=9 N=9
*
p=0.09

27
Discussion
The findings from this study show that GLP-1 concentrations are elevated in IBS-A plasma
and this is correlated with raised IL-6 levels previously reported (Dinan et al., 2006).
However it is likely that GLP-1 is more than just a biomarker of IBS as GLP-1 receptor
staining is expansive and punctate among submucosal neuronal fibres and neurons where it
may contribute to changes in colonic absorption and/or secretion. The data also show that
IL-6 staining was in neuronal bodies among submucosal neurons and co-localised with
GLP-1R in a number of cases, thereby revealing potential for cross-talk between the
molecules.
GLP-1 has been shown to inhibit the MMC of the antrum of the stomach which leads to
inhibition of gastric emptying and also inhibits MMC in the duodenum and jejunum of the
small intestine in both healthy subjects and in IBS subjects. GLP-1 is also shown to inhibit
gastrointestinal motility following a meal (Hellström et al., 2008). GLP-1 analogue ROSE-
010 has also been shown in clinical trials to alleviate abdominal pain and inhibit gastric
emptying without slowing colonic transit in IBS-C patients (Hellström et al., 2009). What
is not clear however is what underlying mechanism GLP-1 is mediating these alleviating
effects and to what extent does it differ among IBS subtypes. GLP-1 and GLP-1R have
been shown to be present in the stomach and in the small intestine (Eissele et al., 1992),
(Hellström et al., 2008) and distal colon (Eissele et al., 1992, Amato et al., 2010). While
submucosal expression of GLP-1Rs has been reported in colonic submucosal neurons
(Baldassano et al., 2012), our findings have shown the type of ganglionic cells expressing
the receptor and co-localisation with the pro-inflammatory cytokine, IL-6.
The GLP-1 receptor co-localises with 63% of nNOS positive neurons. Neuronal nitric
oxide synthase synthesises nitric oxide which is a free radical signalling molecule and acts
as a smooth muscle cell relaxant in the GIT (Shah et al., 2004). GLP-1 causes gastric
accommodation (Delgado-Aros et al., 2002) and this accommodation depends on a nitregic
link (Barragán et al., 1994, Tolessa et al., 1998a). Indeed applying a NO synthase inhibitor
leads to a blockage in postprandial augmentation of gastric volume by GLP-1(Andrews et
al., 2007). nNOS has been found to co-localise with GLP-1R in the myenteric neurons of
the duodenum and proximal colon indicating that nitrergic pathways play a role in the

28
inhibitory effects of GLP-1 on muscle activity (Amato et al., 2010). nNOS has also been
implicated in the pathogenesis of IBS (Reinders et al., 2005, Tjong et al., 2011), with
elevated nNOS expression and NO production from the distal colon of MS rodents (Tjong
et al., 2011) . Clinical studies have also shown elevated rectal mucosal and plasma NO
levels in IBS patients (Reinders et al., 2005, Yazar et al., 2005). Our data suggests the
effects of GLP-1 on submucosal neurons in the distal colon may also be mediated by
nitrergic pathways.
GLP-1R co-localises with 21% of calretinin neurons and 26% of calbindin neurons.
Calretinin is a calcium binding protein and is predominantly in motor neurons. Calbindin is
also a calcium binding protein and is predominantly in sensory neurons (Abalo et al.,
2009). Our data showed the co-localisation between calbindin and GLP-1R was lower than
the co-localisation between nNOS and GLP-1R. Like-wise co-localisation between
calretinin and GLP-1R was lower than the co-localisation between nNOS and GLP-1R.
Our data therefore suggests that the effects of GLP-1R on submucosal neurons of the distal
colon is mediated to a greater extent by nitrergic pathways than motor or sensory
pathways.
A previous study has shown that IL-6 and IL-6R staining was primarily cytosolic and co-
localised with the neuronal cell markers calretinin, calbindin and the glial cell maker S100
(O'Malley et al., 2011b). We have demonstrated that GLP-1R is primarily expressed in
nNOS positive neurons. Interestingly IL-6 receptor has been shown to co-localise with a
similar percentage of nNOS labelled cells as does GLP-1R (O'Malley et al., 2011b,
O'Malley et al., 2011a). In this study IL-6 is shown to be in the neuronal bodies showing
cytosolic expression. Dual labelling with IL-6 and GLP-1R showed that co-localisation is
present in 30% of neuronal bodies revealing a potential area of cross talk. Further studies
to investigate if IL-6 receptors and GLP-1 receptors are in close proximity would also
provide valuable information.
The GLP-1R staining was punctate along the neuronal bodies and fibres. To determine if
these receptor clusters were at synapses neurons were dual labelled with the pre-synaptic
marker, Synapsin I and the post-synaptic marker, PSD-95. GLP-1R clusters appeared to
co-localise with both Synapsin I and PSD-95 indicating that GLP-1 may be important in

29
the synaptic excitation of submucosal neurons and may directly influence the secretion of a
neurotransmitter or act as a neurotransmitter itself.
Cross Sections were obtained in his study to demonstrate the location and quantity of GLP-
1, GLP-1R and IL-6 across the entire cross section of the distal colon. GLP-1 and IL-6
were shown to co-localise within L-cells in the colon. L- cells within the gut secrete GLP-
1 and a study has shown that IL-6 receptors are also expressed on L-cells. IL-6 has also
been shown to cause an increase in GLP-1 secretion from L-cells (Ellingsgaard et al.,
2011), hence with elevations in IL-6 in IBS patients this may result in increased secretion
of GLP-1. Indeed the correlation seen in our study between IL-6 and GLP-1 levels in the
IBS samples would substantiate this theory.
In summary GLP-1R co-localised with calbindin, calretinin, nNOS and S100 and to the
greatest extent with nNOS, therefore concluding that GLP-1 could be exerting its effects
through a nitrergic pathway. Co-localisation studies also allowed us to conclude that a
certain level of cross talk may exist between GLP1-R and IL-6 and this may result in
functional changes in the gut such as altered secretion and absorption. In this way, GLP-1
may contribute to the pathogenesis of IBS.
Further Studies As the ELISA data of IBS-C and IBS-D plasma samples were trending towards
significance further experiments could be carried out with more plasma samples.
The functional role of GLP-1 in IBS may be further characterised by carrying out calcium
imaging studies. Exposure of submucosal neurons to GLP-1 would determine whether
GLP-1 excites or inhibits submucosal neurons. Using an animal model of IBS, the
sensitivity of submucosal neurons in a model of altered GI function could be assessed, as
has been carried out for IL-6 (O'Malley et al., 2011b). Moreover supernatants pooled from
WKY rats could be added to the submucosal preparation from both SD and WKY rats.
Following neuronal excitation by the added supernatant, an anti-GLP-1 neutralising
antibody could then be added to the supernatant. If there is a reduced calcium response it
would show whether GLP-1 in secretions are also involved in exciting submucosal
neurons.

30
To analyse effects of GLP-1 on absorption and secretion across the colonic epithelial
membrane Ussing Chamber electrophysiology experiments could be conducted using SD
and WKY rats. The effects of GLP-1 levels on secretion would be compared between both
rat models. The Ussing chambers design could be exploited to investigate the capacity that
IL-6 has on GLP-1 secretion and also to investigate the capacity that GLP-1 has on IL-6
secretion.
Also as CRF expression is seen to be prominent in the submucosal plexus of the proximal
and distal colon (Yuan et al., 2010) and GLP-1 has been shown to be mediated in stress
induced alteration of colonic transit via CRF mediated pathways (Nakade et al., 2007),
further studies could be carried out to determine potential cross-talk between GLP-1 and
CRF within the submucosal ganglia. Immunofluorescent techniques could be carried out to
determine co-localisation between CRF and GLP-1. Also Ussing chamber experiments
could be carried out to investigate the capacity that CRF has on GLP-1 secretion.
The Immunofluorescent results in this study could be further investigated by carrying out
Western Blot Analysis. Western Blot Analysis would be carried out on colonic human
biopsies to determine the concentration of IL-6 and GLP-1 in the submucosal plexus and
other layers of the colon. It would allow for a comparison of concentrations of IL-6 and
GLP-1 between the different layers of the colon.

31
References ABALO, R., VERA, G., RIVERA, A. J., MORO-RODRÍGUEZ, E. & MARTÍN-
FONTELLES, M. I. 2009. Postnatal maturation of the gastrointestinal tract: a
functional and immunohistochemical study in the guinea-pig ileum at weaning.
Neurosci Lett, 467, 105-10.
AL-SADI, R., BOIVIN, M. & MA, T. 2009. Mechanism of cytokine modulation of
epithelial tight junction barrier. Front Biosci, 14, 2765-78.
AMATO, A., CINCI, L., ROTONDO, A., SERIO, R., FAUSSONE-PELLEGRINI, M. S.,
VANNUCCHI, M. G. & MULÈ, F. 2010. Peripheral motor action of glucagon-like
peptide-1 through enteric neuronal receptors. Neurogastroenterol Motil, 22, 664-
e203.
ANDREWS, C. N., BHARUCHA, A. E., CAMILLERI, M., LOW, P. A., SEIDE, B.,
BURTON, D., BAXTER, K. & ZINSMEISTER, A. R. 2007. Nitrergic contribution
to gastric relaxation induced by glucagon-like peptide-1 (GLP-1) in healthy adults.
Am J Physiol Gastrointest Liver Physiol, 292, G1359-65.
BALDASSANO, S., WANG, G. D., MULÈ, F. & WOOD, J. D. 2012. Glucagon-like
peptide-1 modulates neurally evoked mucosal chloride secretion in guinea pig
small intestine in vitro. Am J Physiol Gastrointest Liver Physiol, 302, G352-8.
BARRAGÁN, J. M., RODRÍGUEZ, R. E. & BLÁZQUEZ, E. 1994. Changes in arterial
blood pressure and heart rate induced by glucagon-like peptide-1-(7-36) amide in
rats. Am J Physiol, 266, E459-66.
BUHNER, S., LI, Q., BERGER, T., VIGNALI, S., BARBARA, G., DE GIORGIO, R.,
STANGHELLINI, V. & SCHEMANN, M. 2012. Submucous rather than myenteric
neurons are activated by mucosal biopsy supernatants from irritable bowel
syndrome patients. Neurogastroenterol Motil, 24, 1134-e572.
CALDARELLA, M. P., MILANO, A., LATERZA, F., SACCO, F., BALATSINOU, C.,
LAPENNA, D., PIERDOMENICO, S. D., CUCCURULLO, F. & NERI, M. 2005.
Visceral sensitivity and symptoms in patients with constipation- or diarrhea-
predominant irritable bowel syndrome (IBS): effect of a low-fat intraduodenal
infusion. Am J Gastroenterol, 100, 383-9.
CAMILLERI, M. & KATZKA, D. A. 2012. Irritable bowel syndrome: methods,
mechanisms, and pathophysiology. Genetic epidemiology and pharmacogenetics in
irritable bowel syndrome. Am J Physiol Gastrointest Liver Physiol, 302, G1075-84.
CAMILLERI, M., MADSEN, K., SPILLER, R., GREENWOOD-VAN MEERVELD, B.,
VAN MEERVELD, B. G. & VERNE, G. N. 2012. Intestinal barrier function in
health and gastrointestinal disease. Neurogastroenterol Motil, 24, 503-12.
CHROUSOS, G. P. 1995. The hypothalamic-pituitary-adrenal axis and immune-mediated
inflammation. N Engl J Med, 332, 1351-62.

32
CORSETTI, M., CAENEPEEL, P., FISCHLER, B., JANSSENS, J. & TACK, J. 2004.
Impact of coexisting irritable bowel syndrome on symptoms and
pathophysiological mechanisms in functional dyspepsia. Am J Gastroenterol, 99,
1152-9.
CRENTSIL, V. 2005. Will corticosteroids and other anti-inflammatory agents be effective
for diarrhea-predominant irritable bowel syndrome? Med Hypotheses, 65, 97-102.
DALRYMPLE, J. & BULLOCK, I. 2008. Diagnosis and management of irritable bowel
syndrome in adults in primary care: summary of NICE guidance. BMJ, 336, 556-8.
DELGADO-AROS, S., KIM, D. Y., BURTON, D. D., THOMFORDE, G. M.,
STEPHENS, D., BRINKMANN, B. H., VELLA, A. & CAMILLERI, M. 2002.
Effect of GLP-1 on gastric volume, emptying, maximum volume ingested, and
postprandial symptoms in humans. Am J Physiol Gastrointest Liver Physiol, 282,
G424-31.
DINAN, T. G. 1994. Glucocorticoids and the genesis of depressive illness. A
psychobiological model. Br J Psychiatry, 164, 365-71.
DINAN, T. G. 1996. Serotonin and the regulation of hypothalamic-pituitary-adrenal axis
function. Life Sci, 58, 1683-94.
DINAN, T. G., CLARKE, G., QUIGLEY, E. M., SCOTT, L. V., SHANAHAN, F.,
CRYAN, J., COONEY, J. & KEELING, P. W. 2008. Enhanced cholinergic-
mediated increase in the pro-inflammatory cytokine IL-6 in irritable bowel
syndrome: role of muscarinic receptors. Am J Gastroenterol, 103, 2570-6.
DINAN, T. G., QUIGLEY, E. M., AHMED, S. M., SCULLY, P., O'BRIEN, S.,
O'MAHONY, L., O'MAHONY, S., SHANAHAN, F. & KEELING, P. W. 2006.
Hypothalamic-pituitary-gut axis dysregulation in irritable bowel syndrome: plasma
cytokines as a potential biomarker? Gastroenterology, 130, 304-11.
DORN, S. D., MORRIS, C. B., HU, Y., TONER, B. B., DIAMANT, N., WHITEHEAD,
W. E., BANGDIWALA, S. I. & DROSSMAN, D. A. 2009. Irritable bowel
syndrome subtypes defined by Rome II and Rome III criteria are similar. J Clin
Gastroenterol, 43, 214-20.
DROSSMAN, D. A. 2005. What does the future hold for irritable bowel syndrome and the
functional gastrointestinal disorders? J Clin Gastroenterol, 39, S251-6.
EISSELE, R., GÖKE, R., WILLEMER, S., HARTHUS, H. P., VERMEER, H., ARNOLD,
R. & GÖKE, B. 1992. Glucagon-like peptide-1 cells in the gastrointestinal tract and
pancreas of rat, pig and man. Eur J Clin Invest, 22, 283-91.
ELLINGSGAARD, H., HAUSELMANN, I., SCHULER, B., HABIB, A. M., BAGGIO, L.
L., MEIER, D. T., EPPLER, E., BOUZAKRI, K., WUEEST, S., MULLER, Y. D.,
HANSEN, A. M., REINECKE, M., KONRAD, D., GASSMANN, M., REIMANN,
F., HALBAN, P. A., GROMADA, J., DRUCKER, D. J., GRIBBLE, F. M.,
EHSES, J. A. & DONATH, M. Y. 2011. Interleukin-6 enhances insulin secretion

33
by increasing glucagon-like peptide-1 secretion from L cells and alpha cells. Nat
Med, 17, 1481-9.
FITZGERALD, L. Z., KEHOE, P. & SINHA, K. 2009. Hypothalamic--pituitary-- adrenal
axis dysregulation in women with irritable bowel syndrome in response to acute
physical stress. West J Nurs Res, 31, 818-36.
FUKUDO, S., NOMURA, T. & HONGO, M. 1998. Impact of corticotropin-releasing
hormone on gastrointestinal motility and adrenocorticotropic hormone in normal
controls and patients with irritable bowel syndrome. Gut, 42, 845-9.
GILLESPIE, C. F. & NEMEROFF, C. B. 2005. Hypercortisolemia and depression.
Psychosom Med, 67 Suppl 1, S26-8.
GONSALKORALE, W. M., MILLER, V., AFZAL, A. & WHORWELL, P. J. 2003a.
Long term benefits of hypnotherapy for irritable bowel syndrome. Gut, 52, 1623-9.
GONSALKORALE, W. M., PERREY, C., PRAVICA, V., WHORWELL, P. J. &
HUTCHINSON, I. V. 2003b. Interleukin 10 genotypes in irritable bowel syndrome:
evidence for an inflammatory component? Gut, 52, 91-3.
GUTNIAK, M. K., LINDE, B., HOLST, J. J. & EFENDIĆ, S. 1994. Subcutaneous
injection of the incretin hormone glucagon-like peptide 1 abolishes postprandial
glycemia in NIDDM. Diabetes Care, 17, 1039-44.
GUYTON, A. C. & HALL, J. E. 2001. Textbook of medical physiology, Philadelphia,
Saunders.
HELLSTRÖM, P. M. 2009. GLP-1: broadening the incretin concept to involve gut
motility. Regul Pept, 156, 9-12.
HELLSTRÖM, P. M. 2011. GLP-1 playing the role of a gut regulatory compound. Acta
Physiol (Oxf), 201, 151-6.
HELLSTRÖM, P. M., HEIN, J., BYTZER, P., BJÖRNSSÖN, E., KRISTENSEN, J. &
SCHAMBYE, H. 2009. Clinical trial: the glucagon-like peptide-1 analogue ROSE-
010 for management of acute pain in patients with irritable bowel syndrome: a
randomized, placebo-controlled, double-blind study. Aliment Pharmacol Ther, 29,
198-206.
HELLSTRÖM, P. M., NÄSLUND, E., EDHOLM, T., SCHMIDT, P. T., KRISTENSEN,
J., THEODORSSON, E., HOLST, J. J. & EFENDIC, S. 2008. GLP-1 suppresses
gastrointestinal motility and inhibits the migrating motor complex in healthy
subjects and patients with irritable bowel syndrome. Neurogastroenterol Motil, 20,
649-59.
KELLER, C., STEENSBERG, A., PILEGAARD, H., OSADA, T., SALTIN, B.,
PEDERSEN, B. K. & NEUFER, P. D. 2001. Transcriptional activation of the IL-6
gene in human contracting skeletal muscle: influence of muscle glycogen content.
FASEB J, 15, 2748-50.

34
KIANK, C., TACHÉ, Y. & LARAUCHE, M. 2010. Stress-related modulation of
inflammation in experimental models of bowel disease and post-infectious irritable
bowel syndrome: role of corticotropin-releasing factor receptors. Brain Behav
Immun, 24, 41-8.
MARTINI, F., NATH, J. L. & BARTHOLOMEW, E. F. 2012. Fundamentals of anatomy
& physiology, San Francisco, Benjamin Cummings.
MARTÍNEZ, V., RYTTINGER, M., KJERLING, M. & ASTIN-NIELSEN, M. 2007.
Characterisation of colonic accommodation in Wistar Kyoto rats with impaired
gastric accommodation. Naunyn Schmiedebergs Arch Pharmacol, 376, 205-16.
MCKENDRICK, M. W. 1996. Post Salmonella irritable bowel syndrome--5 year review. J
Infect, 32, 170-1.
MCKENDRICK, M. W. & READ, N. W. 1994. Irritable bowel syndrome--post salmonella
infection. J Infect, 29, 1-3.
MCKERNAN, D. P., GASZNER, G., QUIGLEY, E. M., CRYAN, J. F. & DINAN, T. G.
2011. Altered peripheral toll-like receptor responses in the irritable bowel
syndrome. Aliment Pharmacol Ther, 33, 1045-52.
MURPHY, D. L., LERNER, A., RUDNICK, G. & LESCH, K. P. 2004. Serotonin
transporter: gene, genetic disorders, and pharmacogenetics. Mol Interv, 4, 109-23.
NAKADE, Y., TSUKAMOTO, K., IWA, M., PAPPAS, T. N. & TAKAHASHI, T. 2007.
Glucagon like peptide-1 accelerates colonic transit via central CRF and peripheral
vagal pathways in conscious rats. Auton Neurosci, 131, 50-6.
O' MAHONY, S. M., COELHO, A. M., FITZGERALD, P., LEE, K., WINCHESTER, W.,
DINAN, T. G. & CRYAN, J. F. 2011. The effects of gabapentin in two animal
models of co-morbid anxiety and visceral hypersensitivity. Eur J Pharmacol, 667,
169-74.
O'MAHONY, S., CHUA, A. S., QUIGLEY, E. M., CLARKE, G., SHANAHAN, F.,
KEELING, P. W. & DINAN, T. G. 2008. Evidence of an enhanced central 5HT
response in irritable bowel syndrome and in the rat maternal separation model.
Neurogastroenterol Motil, 20, 680-8.
O'MAHONY, S. M., BULMER, D. C., COELHO, A. M., FITZGERALD, P.,
BONGIOVANNI, C., LEE, K., WINCHESTER, W., DINAN, T. G. & CRYAN, J.
F. 2010. 5-HT(2B) receptors modulate visceral hypersensitivity in a stress-sensitive
animal model of brain-gut axis dysfunction. Neurogastroenterol Motil, 22, 573-8,
e124.
O'MAHONY, S. M., MARCHESI, J. R., SCULLY, P., CODLING, C., CEOLHO, A. M.,
QUIGLEY, E. M., CRYAN, J. F. & DINAN, T. G. 2009. Early life stress alters
behavior, immunity, and microbiota in rats: implications for irritable bowel
syndrome and psychiatric illnesses. Biol Psychiatry, 65, 263-7.

35
O'MALLEY, D., DINAN, T. G. & CRYAN, J. F. 2011a. Altered expression and secretion
of colonic interleukin-6 in a stress-sensitive animal model of brain-gut axis
dysfunction. J Neuroimmunol, 235, 48-55.
O'MALLEY, D., DINAN, T. G. & CRYAN, J. F. 2012. Interleukin-6 modulates colonic
transepithelial ion transport in the stress-sensitive wistar kyoto rat. Front
Pharmacol, 3, 190.
O'MALLEY, D., JULIO-PIEPER, M., GIBNEY, S. M., DINAN, T. G. & CRYAN, J. F.
2010. Distinct alterations in colonic morphology and physiology in two rat models
of enhanced stress-induced anxiety and depression-like behaviour. Stress, 13, 114-
22.
O'MALLEY, D., LISTON, M., HYLAND, N. P., DINAN, T. G. & CRYAN, J. F. 2011b.
Colonic soluble mediators from the maternal separation model of irritable bowel
syndrome activate submucosal neurons via an interleukin-6-dependent mechanism.
Am J Physiol Gastrointest Liver Physiol, 300, G241-52.
OLDEN, K. W. 2003. Approach to the Patient with Severe, Refractory Irritable Bowel
Syndrome. Curr Treat Options Gastroenterol, 6, 311-317.
ORTIZ-LUCAS, M., SAZ-PEIRÓ, P. & SEBASTIÁN-DOMINGO, J. J. 2010. Irritable
bowel syndrome immune hypothesis. Part two: the role of cytokines. Rev Esp
Enferm Dig, 102, 711-7.
REINDERS, C. I., HERULF, M., LJUNG, T., HOLLENBERG, J., WEITZBERG, E.,
LUNDBERG, J. O. & HELLSTRÖM, P. M. 2005. Rectal mucosal nitric oxide in
differentiation of inflammatory bowel disease and irritable bowel syndrome. Clin
Gastroenterol Hepatol, 3, 777-83.
SAITO, Y. A., MITRA, N. & MAYER, E. A. 2010. Genetic approaches to functional
gastrointestinal disorders. Gastroenterology, 138, 1276-85.
SAULNIER, D. M., RINGEL, Y., HEYMAN, M. B., FOSTER, J. A., BERCIK, P.,
SHULMAN, R. J., VERSALOVIC, J., VERDU, E. F., DINAN, T. G., HECHT, G.
& GUARNER, F. 2013. The intestinal microbiome, probiotics and prebiotics in
neurogastroenterology. Gut Microbes, 4, 17-27.
SCHELLER, J., CHALARIS, A., SCHMIDT-ARRAS, D. & ROSE-JOHN, S. 2011. The
pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim
Biophys Acta, 1813, 878-88.
SCHIRRA, J., WANK, U., ARNOLD, R., GÖKE, B. & KATSCHINSKI, M. 2002. Effects
of glucagon-like peptide-1(7-36)amide on motility and sensation of the proximal
stomach in humans. Gut, 50, 341-8.
SCHJOLDAGER, B. T., MORTENSEN, P. E., CHRISTIANSEN, J., ORSKOV, C. &
HOLST, J. J. 1989. GLP-1 (glucagon-like peptide 1) and truncated GLP-1,
fragments of human proglucagon, inhibit gastric acid secretion in humans. Dig Dis
Sci, 34, 703-8.

36
SEELEY, R. R., STEPHENS, T. D. & TATE, P. 2003. Anatomy & physiology, Boston,
McGraw-Hill.
SHAH, V., LYFORD, G., GORES, G. & FARRUGIA, G. 2004. Nitric oxide in
gastrointestinal health and disease. Gastroenterology, 126, 903-13.
THARAKAN, A., NORTON, I. T., FRYER, P. J. & BAKALIS, S. 2010. Mass transfer
and nutrient absorption in a simulated model of small intestine. J Food Sci, 75,
E339-46.
TJONG, Y. W., IP, S. P., LAO, L., WU, J., FONG, H. H., SUNG, J. J., BERMAN, B. &
CHE, C. T. 2011. Role of neuronal nitric oxide synthase in colonic distension-
induced hyperalgesia in distal colon of neonatal maternal separated male rats.
Neurogastroenterol Motil, 23, 666-e278.
TOLESSA, T., GUTNIAK, M., HOLST, J. J., EFENDIC, S. & HELLSTRÖM, P. M.
1998a. Glucagon-like peptide-1 retards gastric emptying and small bowel transit in
the rat: effect mediated through central or enteric nervous mechanisms. Dig Dis Sci,
43, 2284-90.
TOLESSA, T., GUTNIAK, M., HOLST, J. J., EFENDIC, S. & HELLSTRÖM, P. M.
1998b. Inhibitory effect of glucagon-like peptide-1 on small bowel motility. Fasting
but not fed motility inhibited via nitric oxide independently of insulin and
somatostatin. J Clin Invest, 102, 764-74.
TURNBULL, A. V. & RIVIER, C. L. 1999. Regulation of the hypothalamic-pituitary-
adrenal axis by cytokines: actions and mechanisms of action. Physiol Rev, 79, 1-71.
VAN DER VEEK, P. P., VAN DEN BERG, M., DE KROON, Y. E., VERSPAGET, H.
W. & MASCLEE, A. A. 2005. Role of tumor necrosis factor-alpha and interleukin-
10 gene polymorphisms in irritable bowel syndrome. Am J Gastroenterol, 100,
2510-6.
XIA, Y., HU, H. Z., LIU, S., REN, J., ZAFIROV, D. H. & WOOD, J. D. 1999. IL-1beta
and IL-6 excite neurons and suppress nicotinic and noradrenergic
neurotransmission in guinea pig enteric nervous system. J Clin Invest, 103, 1309-
16.
YALE, S. H., MUSANA, A. K., KIEKE, A., HAYES, J., GLURICH, I. & CHYOU, P. H.
2008. Applying case definition criteria to irritable bowel syndrome. Clin Med Res,
6, 9-16.
YAZAR, A., BÜYÜKAFPAR, K., POLAT, G., PATA, C., KANÝK, A., TIFTIK, E. N. &
BAĞDATOĞLU, O. 2005. The urinary 5-hydroxyindole acetic acid and plasma
nitric oxide levels in irritable bowel syndrome: a preliminary study. Scott Med J,
50, 27-9.
YUAN, P. Q., WU, S. V., WANG, L. & TACHÉ, Y. 2010. Corticotropin releasing factor
in the rat colon: expression, localization and upregulation by endotoxin. Peptides,
31, 322-31.

37


















