A preliminary study of functional connectivity of medication naïve children with...
8
A preliminary study of functional connectivity of medication naïve children with obsessive–compulsive disorder Alexander Mark Weber a , Noam Soreni a,b,c, ⁎, Michael David Noseworthy a,d,e,f a School of Biomedical Engineering, McMaster University, Hamilton, Ontario, Canada b Department of Psychiatry and Behavioural Neuroscience, McMaster University, Hamilton, Ontario, Canada c Offord Centre for Child Studies, McMaster Children's Hospital, Hamilton, Ontario, Canada d Electrical & Computer Engineering, McMaster University, Hamilton, Ontario, Canada e Medical Physics & Applied Radiation Sciences, McMaster University, Hamilton, Ontario, Canada f Department of Radiology, McMaster University, Hamilton, Ontario, Canada abstract article info Article history: Received 3 September 2013 Received in revised form 7 March 2014 Accepted 1 April 2014 Available online 12 April 2014 Keywords: Cingulate network Cortico-striatal–thalamic–cortical (CSTC) Independent component analysis (ICA) Obsessive–compulsive disorder Pediatric Resting state networks (RSN) Background: Evidence suggests that obsessive–compulsive disorder (OCD) is associated with a dysfunction in the cortico-striatal–thalamic–cortical (CSTC) circuitry. Resting state functional connectivity magnetic resonance imaging (rs-fcMRI) allows measurements of resting state networks (RSNs), brain networks that are present at ‘rest’. However, although OCD has a typical onset during childhood or adolescence, only two other studies have performed rs-fcMRI comparisons of RSNs in children and adolescents with OCD against healthy controls. Methods: In the present study, we performed resting state functional magnetic resonance imaging using a 3 Tesla MRI, in 11 medication-naïve children and adolescents with OCD and 9 healthy controls. In contrast to previous studies that relied on a priori determination of RSNs, we determined resting state functional connectivity with a data-driven independent component analysis (ICA). Results: Consistent with previous reports in healthy adults, we identified 13 RSNs. Case–control un-adjusted statistical significance (p b 0.05) was found for two networks. Firstly, increased connectivity (OCD N control) in the right section of Brodmann area 43 of the auditory network; Secondly, decreased connectivity in the right section of Brodmann area 8 and Brodmann area 40 in the cingulate network. Conclusions: Our preliminary findings of case–control differences in RSNs lend further support to the CSTC hypothesis of OCD, as well as implicating other regions of the brain outside of the CSTC. © 2014 Elsevier Inc. All rights reserved. 1. Introduction Obsessive–compulsive disorder (OCD) is a common neuropsychiatric disorder with a lifetime prevalence of between 1 and 2.5% (Bebbington, 1998; Horwath and Weissman, 2000; Ruscio et al., 2010). Lifetime prev- alence of OCD for children and adolescents has been estimated to be around 0.25–2.7% (Heyman et al., 2003; Rapoport et al., 2000), which in- creases exponentially with increasing age (up to 18) (Heyman et al., 2003). OCD in children and adolescents is associated with functional im- pairments in home, school and social settings (Valderhaug and Ivarsson, 2005). Structural and functional neuroimaging studies of youth and adults with OCD suggest an impairment of cortico-striatal–thalamic–cortical (CSTC) circuits (Graybiel and Rauch, 2000; Saxena et al., 1998, 2001). Recently, a growing number of functional magnetic resonance imaging (fMRI) studies have been used to probe CSTC activation in OCD using an experimental ‘resting’ state, in which the subject is asked to simply relax, remain awake and not to think of anything in particular (Biswal et al., 1995). Resting state fMRI is often analyzed using a functional connectivity approach termed resting state functional connectivity MRI (rs-fcMRI) (Damoiseaux et al., 2006). rs-fcMRI analysis usually targets low frequency (b 0.1 Hz), synchronized activations (also known as low-frequency blood oxygen level dependent (BOLD) fluctuations) in spatially separated areas of the brain (Friston et al., 1993). These synchronized neuro-physiological events, active at rest, represent structurally and functionally connected Progress in Neuro-Psychopharmacology & Biological Psychiatry 53 (2014) 129–136 Abbreviations: OCD, obsessive–compulsive disorder; CSTC, cortico-striatal–thalamic– cortical; MRI, Magnetic Resonance Imaging; rs-fcMRI, resting state functional connectivity magnetic resonance imaging; RSN, resting state network; ICA, independent component analysis; BOLD, blood oxygen level dependent; CCA, cross-correlation-analysis; ROI, region of interest; ACC, anterior cingulate cortex; PCC, posterior cingulate cortex; DSM- IV, Diagnostic and Statistical Manual of Mental Disorders IV; K-SADS-PL, Kiddie-Sads- Present and Lifetime; REB, Research Ethics Board; CY-BOCS, Child Yale–Brown Obsessive–Compulsive Scale; MELODIC, Multivariate Exploratory Linear Optimized Decomposition into Independent Components; FWHM, full-width-at-half-maximum; BA, Brodmann area; DMN, default mode network; mPFC, medial prefrontal cortex; DLPFC, dorsolateral prefrontal cortex. ⁎ Corresponding author at: Pediatric OCD Consultation Team, Anxiety Treatment and Research Center, St. Joseph's Healthcare, 50 Charlton Ave. East, Hamilton, Ontario L8N 4A6, Canada. Tel.: +1 905 522 1155x33139. E-mail address: [email protected] (N. Soreni). http://dx.doi.org/10.1016/j.pnpbp.2014.04.001 0278-5846/© 2014 Elsevier Inc. All rights reserved. Contents lists available at ScienceDirect Progress in Neuro-Psychopharmacology & Biological Psychiatry journal homepage: www.elsevier.com/locate/pnp
-
Upload
michael-david -
Category
Documents
-
view
214 -
download
2
Transcript of A preliminary study of functional connectivity of medication naïve children with...

Progress in Neuro-Psychopharmacology & Biological Psychiatry 53 (2014) 129–136
Contents lists available at ScienceDirect
Progress in Neuro-Psychopharmacology & BiologicalPsychiatry
j ourna l homepage: www.e lsev ie r .com/ locate /pnp
A preliminary study of functional connectivity of medication naïvechildren with obsessive–compulsive disorder
Alexander Mark Weber a, Noam Soreni a,b,c,⁎, Michael David Noseworthy a,d,e,f
a School of Biomedical Engineering, McMaster University, Hamilton, Ontario, Canadab Department of Psychiatry and Behavioural Neuroscience, McMaster University, Hamilton, Ontario, Canadac Offord Centre for Child Studies, McMaster Children's Hospital, Hamilton, Ontario, Canadad Electrical & Computer Engineering, McMaster University, Hamilton, Ontario, Canadae Medical Physics & Applied Radiation Sciences, McMaster University, Hamilton, Ontario, Canadaf Department of Radiology, McMaster University, Hamilton, Ontario, Canada
Abbreviations: OCD, obsessive–compulsive disorder; Ccortical; MRI, Magnetic Resonance Imaging; rs-fcMRI, restmagnetic resonance imaging; RSN, resting state networkanalysis; BOLD, blood oxygen level dependent; CCA, cregion of interest; ACC, anterior cingulate cortex; PCC, poIV, Diagnostic and Statistical Manual of Mental DisordePresent and Lifetime; REB, Research Ethics Board;Obsessive–Compulsive Scale; MELODIC, MultivariateDecomposition into Independent Components; FWHM,BA, Brodmann area; DMN, default mode network; mPDLPFC, dorsolateral prefrontal cortex.⁎ Corresponding author at: Pediatric OCD Consultation
Research Center, St. Joseph's Healthcare, 50 Charlton Av4A6, Canada. Tel.: +1 905 522 1155x33139.
E-mail address: [email protected] (N. Soreni).
http://dx.doi.org/10.1016/j.pnpbp.2014.04.0010278-5846/© 2014 Elsevier Inc. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:
Received 3 September 2013Received in revised form 7 March 2014Accepted 1 April 2014Available online 12 April 2014Keywords:Cingulate networkCortico-striatal–thalamic–cortical (CSTC)Independent component analysis (ICA)Obsessive–compulsive disorderPediatricResting state networks (RSN)
Background: Evidence suggests that obsessive–compulsive disorder (OCD) is associatedwith a dysfunction in thecortico-striatal–thalamic–cortical (CSTC) circuitry. Resting state functional connectivity magnetic resonanceimaging (rs-fcMRI) allows measurements of resting state networks (RSNs), brain networks that are present at‘rest’. However, although OCD has a typical onset during childhood or adolescence, only two other studieshave performed rs-fcMRI comparisons of RSNs in children and adolescents with OCD against healthy controls.Methods: In the present study,we performed resting state functionalmagnetic resonance imaging using a 3 TeslaMRI, in 11 medication-naïve children and adolescents with OCD and 9 healthy controls. In contrast to previousstudies that relied on a priori determination of RSNs, we determined resting state functional connectivity witha data-driven independent component analysis (ICA).Results: Consistent with previous reports in healthy adults, we identified 13 RSNs. Case–control un-adjustedstatistical significance (p b 0.05) was found for two networks. Firstly, increased connectivity (OCD N control)in the right section of Brodmann area 43 of the auditory network; Secondly, decreased connectivity in the
right section of Brodmann area 8 and Brodmann area 40 in the cingulate network.Conclusions: Our preliminary findings of case–control differences in RSNs lend further support to the CSTChypothesis of OCD, as well as implicating other regions of the brain outside of the CSTC.© 2014 Elsevier Inc. All rights reserved.
1. Introduction
Obsessive–compulsive disorder (OCD) is a common neuropsychiatricdisorder with a lifetime prevalence of between 1 and 2.5% (Bebbington,1998; Horwath andWeissman, 2000; Ruscio et al., 2010). Lifetime prev-alence of OCD for children and adolescents has been estimated to be
STC, cortico-striatal–thalamic–ing state functional connectivity; ICA, independent componentross-correlation-analysis; ROI,sterior cingulate cortex; DSM-rs IV; K-SADS-PL, Kiddie-Sads-CY-BOCS, Child Yale–BrownExploratory Linear Optimizedfull-width-at-half-maximum;FC, medial prefrontal cortex;
Team, Anxiety Treatment ande. East, Hamilton, Ontario L8N
around 0.25–2.7% (Heyman et al., 2003; Rapoport et al., 2000), which in-creases exponentially with increasing age (up to 18) (Heyman et al.,2003). OCD in children and adolescents is associated with functional im-pairments in home, school and social settings (Valderhaug and Ivarsson,2005).
Structural and functional neuroimaging studies of youth and adultswith OCD suggest an impairment of cortico-striatal–thalamic–cortical(CSTC) circuits (Graybiel and Rauch, 2000; Saxena et al., 1998, 2001).Recently, a growing number of functional magnetic resonance imaging(fMRI) studies have been used to probe CSTC activation in OCD usingan experimental ‘resting’ state, in which the subject is asked to simplyrelax, remain awake and not to think of anything in particular (Biswalet al., 1995).
Resting state fMRI is often analyzed using a functional connectivityapproach termed resting state functional connectivity MRI (rs-fcMRI)(Damoiseaux et al., 2006). rs-fcMRI analysis usually targets low frequency(b0.1 Hz), synchronized activations (also known as low-frequency bloodoxygen level dependent (BOLD) fluctuations) in spatially separated areasof the brain (Friston et al., 1993). These synchronized neuro-physiologicalevents, active at rest, represent structurally and functionally connected

130 A.M. Weber et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 53 (2014) 129–136
networks, termed resting state networks (RSNs). rs-fcMRI can be ana-lyzed by two different approaches: one that uses strong a priori knowl-edge, and the other which is almost exclusively data driven (Ma et al.,2007). Most rs-fcMRI analyses to date have used cross-correlation-analysis (CCA), an a priori driven method, which looks at correlationsbetween each voxel and a pre-determined function (Biswal et al., 1995;Ma et al., 2007). This pre-determined function is often taken to be apreselected region of interest (ROI), or seed-voxel. In contrast, indepen-dent component analysis (ICA; McKeown et al., 1998), lacks an ROI apriori assumption. Instead, ICA is based on the assumption that activa-tions are independent to other signal variations (motion, cardiac andrespiratory fluctuations, etc.), and extract components (RSNs) basedon enforcing orthogonality spatially.
To date, there have been 11 published rs-fcMRI studies inOCD thatwehave found (Fitzgerald et al., 2010, 2011; Fontenelle et al., 2012; Harrisonet al., 2009; Jang et al., 2010; Kang et al., 2013; Li et al., 2012; Meunieret al., 2012; Sakai et al., 2010; Stern et al., 2012a; Zhang, 2011), all ofwhich had relied on a CCA approach with seed-voxels placed in variousregions of the brain and reported statistically significant differencesbetween patients and controls.
Although OCD has a high rate of onset during childhood and adoles-cence, only two rs-fcMRI studies (Fitzgerald et al., 2010, 2011) havefocused on children and adolescents with the disorder. This is of partic-ular importance given that RSNs in healthy subjects go through manydevelopmental changes during the transition from childhood to adult-hood (Fair et al., 2008, 2009; Stevens et al., 2009; Supekar et al.,2010). The first rs-fcMRI study that looked at children and adolescents(Fitzgerald et al., 2010) reported decreased connectivity between thedorsal anterior cingulate cortex (ACC) and right anterior operculum,as well as between the ventral medial frontal cortex and the posteriorcingulate cortex (PCC), in OCDpatients as compared to healthy controls.The second study (Fitzgerald et al., 2011) reported decreased connec-tivity between the dorsal striatum and rostral ACC, two structures thatwere not the focus of their initial report (Fitzgerald et al., 2010). Thus,preliminary RSN findings in youth with OCD support the hypothesisthat differences in RSN activation exist between children and adoles-cents with OCD and their healthy counterparts. However, evidence iscurrently limited by the small number of studies and the reliance on aCCA analysis approach in existing studies.
The present fMRI pilot study compared resting state activations be-tweenmedication naïve children and adolescentswithOCD and healthycontrols. Unlike previous reports, we opted to use ICA as it provides adata-driven analysis of the resting state fMRI data andminimizes depen-dency on seed-voxel location and inter-subject anatomical variability. Asthis was the first study to use ICA in children and adolescents with OCD,we did not limit ourselves to pre-determined RSNs, but instead decidedto compare all identifiable RSNs between patients with OCD and healthycontrols. We hypothesized, based on the study by Fitzgerald et al. (2011)that we would find decreased connectivity between the cingulate cortexand caudate during ‘rest’ activation.
2. Methods and materials
2.1. Subjects
15 psychotropic naïve children and adolescents with OCD (age8–16 years old) were recruited from the Pediatric OCD ConsultationTeamat the Anxiety Treatment and Research Center, Hamilton, Ontario.Patients had a primary diagnosis of OCD according to DSM-IV criteria(American Psychiatric Association, 1994). All DSM-IV diagnoses (OCDand co-morbid conditions) were made by a child psychiatrist (N.S.)using the K-SADS-PL (Kaufman et al., 1997) A total of 13 healthy com-parison subjects (age 8–16 years old) were recruited from the generalcommunity through advertisements. Control subjects did not haveany personal history of psychiatric illness, treatment with psychotropicmedications or a diagnosed learning disability.
Exclusion criteria for patients and controls included lifetime historyof psychosis, bipolar disorder, conduct disorder, substance abuse/dependence, an eating disorder, significant medical or neurologic disor-ders or a previously diagnosed learning disability. The St. Joseph'sHealthcare Research Ethics Board (REB) approved the study. Before initi-ating all studies, legal guardians provided written informed consent, andall children 16 years of age and younger gavewritten assent. Adolescents16 years of age and older and their parents gave written informed con-sent before initiating all studies.
2.2. Clinical measures
All patientswere assessed for OCD symptom severity using the ChildYale–Brown Obsessive–Compulsive Scale (CY-BOCS) (Goodman et al.,1991). The CY-BOCS is a clinician-rated semi-structured instrumentwith good inter-rater reliability and validity for children and adolescents(Storch, 2006). To ensure that patients had at least mild OCD symptoms,only patients with CY-BOCS total scores N11 were recruited to theimaging study (Scahill et al., 1997).
2.3. Imaging
All scanning was done on a GE Signa 3 T HDx twinspeed short boreMRI system using an 8-channel phased array receive-only head RF coil(GE Healthcare, Milwaukee, WI). Each session involved a localizerscan (30 s) and a high-resolution 3D IR-prepped fSPGR T1-weightedimaging sequence (24 cm field of view, TE/TR/TI = 2.1/7.5/450 ms,flip angle=12°, 512 × 512matrix, 1mmthick/0mmskip, 148 acquiredslices, voxels were reconstructed to 0.469 × 0.469 × 1 mm). Restingstate BOLD data was acquired with a T2*-weighted gradient echo planarimaging (GE-EPI) sequence, with the following parameters: 64 × 64matrix, 28 axial slices (5 mm thick, no skip), TR = 2000 ms, TE =35 ms, flip angle = 90°, 240 images per slice, and angled to AC/PCalignment. Subjects were asked to close their eyes, lay still and thinkof nothing in particular.
2.4. Pre-processing
Data from each subject was corrected for interleaved slice acquisitionand 3D rigid body motion, and was aligned to anatomical data using thesoftware application AFNI (Cox, 1996). Of an original participant group of13 controls and 15 OCD subjects, 4 controls and 4 OCD subjects wereexcluded from analysis due to motions greater than 2 mm or degrees insingle scan session. Next, using the FSL software package MELODIC(Multivariate Exploratory Linear Optimized Decomposition into Inde-pendent Components) (Beckmann and Smith, 2004), all functionaldata were first registered to brain-extracted high resolution T1-weighted anatomical scans, then registered to MNI152 standard spaceat 2 mm resampling resolution. Images were then filtered with ahigh-pass temporal cutoff of 0.009 Hz, and were spatially smoothedwith a Gaussian kernel with full-width-at-half-maximum (FWHM) of5 mm.
2.5. rs-fcMRI data analysis
Using MELODIC, probabilistic independent component analysis wasperformed on all subjects, at a single-group level, to decompose the 4Ddata sets into separate spatial maps. This was accomplished by using amulti-session temporal concatenation approach,whichworks by stackingall 2D (space(voxels) ∗ time) data matrices of every data-set on top ofeach other. The concatenation method was chosen due to the fact thatRSNs between subjects are not expected to have the same time-courses.The present study implemented the following options in MELODIC:time-courses were variance-normalized in order to stress voxel-wisetemporal dynamics over mean signal; multi-session temporal concatena-tion (as already mentioned); the number of components (RSNs) was

131A.M. Weber et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 53 (2014) 129–136
manually chosen at 25; a level of 0.5 was used as the threshold level forindependent components (a voxel will be reported if the probability ofit belonging to non-backgroundmixtures exceeds the probability of it be-longing to the background Gaussian noise is exceeded (Beckmann andSmith, 2004)).
Subject-specific RSNs (ones found from the single-group level anal-ysis) were then mapped onto each subject through the FSL software‘Dual Regression’ (Beckmann et al., 2009). This software package firstuses the spatial maps found from MELODIC to find temporal dynamicsassociated with each map. These time-courses are then used as a setof temporal regressors in a general linear model to find the samemaps for each subject.
2.6. Statistics
Group differences were obtained from the Dual Regression program,where the different sets of spatial maps were collected across subjectsinto single 4D files (one per RSN) and analyzed across groups (OCD vs.Control) using non-parametric statistical analysis involving permuta-tion testing (Beckmann et al., 2009). The number of permutations wasset at 10,000 to maximize alpha level calculations (Webster, 2012). Asthis study is thefirst ICA analysis ofOCDwithno specific RSNbeing tested,it was treated as exploratory in nature (i.e. as a pilot study); thus, correc-tion for multiple-comparisons (Bonferroni) was not performed.
3. Results
3.1. Subjects
The final OCD sample size consisted of 11medication-naïve childrenand adolescents with OCD (mean age 13.0 years, SD= 2.9). OCD symp-tomswere in themoderate range. The control group included 9 healthychildren and adolescents (mean age 12.7 years, SD = 3.2) (Table 1,sample characteristics). No significant sex or age differences betweenpatients and controls were found.
3.2. Resting state networks
Of the 25 component maps that were created through MELODIC, 13were identified (Fig. 1) as being physiologically relevant, based onknown neuroanatomical RSNs (Table 2) from previous ICA literaturereports in adults, as there are no known ICA studies of children andadolescents. The other 12 networks are believed to be either artifactualor currently un-recognized.
3.3. Statistics
Of these 13 RSNs, un-adjusted statistical significance (p b 0.05) wasfound for two networks (Fig. 2). Increased connectivity (OCD N control)in the auditory network (RSNE; Fig. 2a)was found in the right section ofBrodmann area (BA) 43, which is responsible for sensorimotor repre-sentation of themouth (Bernal and Perdomo, 2008). Decreased connec-tivity was found in RSNM (cingulate network; Fig. 2b) in the right
Table 1Sample demographics and OCD symptom severity.
OCD Control Significance
Sample size 11 9Sex Girls—6
Boys—6Girls—4Boys—5
0.66a
Mean age (years) 13.0 ± 2.9 12.7 ± 3.2 0.81b
Age range 9–16 8–16CY-BOCS total score 22.7 ± 5.2 (range 16–30) N/A
CY-BOCS: Child Yale–Brown Obsessive–Compulsive Scale.a Chi-square results.b Independent sample t-test results.
section of BA 8, which is responsible for uncertainty (Volz et al.,2005), or alternatively for hope, a higher-order expectation positivelycorrelated with uncertainty (Chew and Ho, 1994). Decreased connec-tivity in RSNM was also found in BA 40, which is involved in spatial ori-entation and semantic representation (reading, both in regard tomeaning and phonology) (Stoeckel et al., 2009).
4. Discussion
The present pilot study is the first RSN analysis in OCD subjects (andone of the first in children in general) that used an ICA approach for adata-driven separation of spatial maps. Our preliminary results identified13 RSNs based on previously reported networks in the literature. Of these13 networks, un-adjusted statistical significance was discovered in twonetworks: increased connectivity (OCD N control) in the auditorynetwork in the right section of BA 43; and decreased connectivity wasfound in the cingulate network in the right section of BA 8 and BA 40.
The first finding of the present study is the identification of 13 rest-ing networks that are in line with previous literature reports in healthyadults (Damoiseaux et al., 2006; Heine et al., 2012). Several seed-basedRSN studies done on children and adolescents (Fair et al., 2008, 2009;Supekar et al., 2010), as well as an ICA study looking at RSN changesfrom adolescence to adulthood (Stevens et al., 2009), have shown thatin some cases a single RSN in the adult brain is actually two or severalindependent RSNs in children and adolescents. RSNs in children are typ-ically anatomically localized, with more globally distributed architec-ture appearing in late adolescence or young adulthood (Fair et al.,2009). In line with this report, the present study identified three inde-pendent spatial maps (RSNA, RSNB and RSNC) that may represent thedeveloping default mode network (DMN) in children and adolescents(Fair et al., 2008, 2009; Supekar et al., 2010). Indeed, the link betweenthe PCC and the medial prefrontal cortex (mPFC) could be significantlyweaker in children compared with adults (Fair et al., 2008, 2009;Supekar et al., 2010). Furthermore, although this finding has not beenreported before, our findings suggest that the sensorimotor networkin children and adolescents may consist of two separate spatial maps:RSNI and RSNJ.
Significant between-group differences were found for RSNM
(cingulate network) and RSNE (auditory network). First, decreasedconnectivity (OCD b Control; p b 0.05)was found in the RSNM cingulatenetwork (Fig. 2b). Connectivity differences in the cingulate networkbetween subjects with OCD and healthy controls support the CSTC the-ory of OCD, as the cingulate network is composed of the frontal cortex(medial frontal/ACC, bilateral dorsolateral prefrontal cortex (DLPFC))and the striatum, both of which are vital components that make upthe CSTC. The finding of decreased connectivity is in line with the soleprevious study (Fitzgerald et al., 2011) that looked at caudate/striatumto frontal lobe connectivity in children. In this study, Fitzgerald andcolleagues reported reduced connectivity between the dorsal striatumand rostral ACC. This finding is in contrast with previous seed-basedRSN studies in adults with OCD that looked at the caudate/striatum(Fontenelle et al., 2012; Harrison et al., 2009; Kang et al., 2013).
Specifically, decreased connectivity in the cingulate network wasfound in the right section of BA 8 and in BA 40 (Fig. 2b). BA 8 has beenshown previously to be activated during increased uncertainty in nor-mal healthy adults (Volz et al., 2005). Indeed, the role of the cingulatenetwork in monitoring and managing conflicts is well described in thecontext of OCD (Melloni et al., 2012). For example, during check/re-check rituals, it would be expected that the patient is ‘re-checking’a lock (for example) because they are uncertain if it is indeed locked(Stern et al., 2012b). Because the present resting-state study did notaim to simulate an OCD-related conflict (i.e. door closed or shut), ourfindings of abnormal cingulate network activation during rest comple-ment rather than replicate previous fMRI studies of executive functiontask in OCD (Huyser et al., 2009; Kang et al., 2013; Melloni et al., 2012).

Fig. 1. Probabilistic ICA estimated RSNmaps of group analysis: axial, coronal, and sagittal views of spatialmaps for each component. Images are thresholded z-statistics overlaid on the average high-resolution scan transformed into standard (MNI152)space. Red to blue are z-values, ranging from 5 to 10. The left side of the image refers to the right hemisphere of the brain. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
132A.M
.Weber
etal./Progressin
Neuro-Psychopharm
acology&
BiologicalPsychiatry53
(2014)129
–136

Table 2The 13 resting state networks identified (Fig. 1) with anatomical locations, function and references.
RSN Anatomical locations Function References
A DMN Precuneus/PCC; mediofrontal/ACC;inferior parietal lobe; and medialtemporal lobe
Network when there are no goal-directed tasks. Buckner et al. (2008) and Raichle et al. (2001)Associated with introspective mental processes,episodic memories, and imagining future events.
B Emotionalregulation/anteriorDMN
Perigenual ACC; and mPFC Mentalizing and self-reflection. Amodio and Frith (2006), Fair et al. (2008), Frith and Frith(2003), Johnson et al. (2002), Kelly et al. (2009), andOchsner et al. (2005)
Identified as the front half of the developing DMN
C Socialprocessing/AnteriorDMN
Rostral supragenual ACC Evaluative functions such as monitoring, signaling ofconflict or interference, response to errors and decisionmaking.
Botvinick et al. (1999), Botvinick et al. (2004), Garavan et al.(2003), Kelly et al. (2009), Kiehl et al. (2000), Kroger et al.(2002), Luo et al. (2003), and Paulus and Frank (2006)
May be another front section of the developing DMN(Kelly et al., 2009)
D Saliencenetwork
Fronto-insular; ACC;subcortical; and limbic areas
Separates relevant from irrelevant information. Beckmann et al. (2005) and Menon and Uddin (2010)
E Auditorynetwork
Primary and secondary auditorycortices
Important for tone/pitch discrimination, music, and speech. Heine et al. (2012) and Laird et al. (2011)
F Visualoccipitalnetwork
Lateral and superior occipital gyrus Important in higher-order visual stimuli (e.g., orthography). Damoiseaux et al. (2006), Heine et al. (2012), and Laird et al.(2011)
G Visual lateralnetwork
Superior parietal cortex;occipitotemporal; and precentalareas
Processing of complex (emotional) visual stimuli. Damoiseaux et al. (2006), Heine et al. (2012), and Laird et al.(2011)
H Visualmedialnetwork
Part of striate and parastriate Processing of simple visual stimuli(e.g., a flickering checkerboard).
Damoiseaux et al. (2006), Heine et al. (2012), and Laird et al.(2011)
I Posterior halfofsensorimotornetwork
Pre-central gyrus Activation resembles the activations seen in motor tasks. Biswal et al. (1995), Damoiseaux et al. (2006), and Heineet al. (2012)
J Lateral half ofsensorimotornetwork
Post-central gyrus Activation resembles the activations seen in motor tasks. Biswal et al. (1995), Damoiseaux et al. (2006), and Heineet al. (2012)
K Rightexecutivecontrolnetwork
Right hemisphere: middle frontal;orbital; superior parietal; middletemporal gyrus; and posteriorcingulate
The fronto-parietal component has been associated withmemory, language, attention and visual processes.Damoiseaux et al. suggests that both executive controlnetworks areinvolved in memory function, while Smith et al. have gonefurther by suggesting that the right network is involved inpain and the left network is involved in language.
Damoiseaux et al. (2006), Heine et al. (2012),Rosazza and Minati (2011), and Smith et al. (2009)
L Leftexecutivecontrolnetwork
Left hemisphere: middle frontal;orbital; superior parietal; middletemporal gyrus; and posteriorcingulate
See right executive control network (above) Damoiseaux et al. (2006) and Heine et al. (2012)
M Cingulatenetwork
Medial frontal/ACC; bilateral DLPFC;bilateral temporal cortex; andstriatum
Emotion formation and processing. Bush et al. (2000), Bush et al. (2002), Chen et al. (2008),and Nielsen et al. (2005)Linking behavioral outcomes with motivation.
133A.M. Weber et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 53 (2014) 129–136
BA 40 has been shown to be involved in spatial orientation andsemantic representation (reading, both in regard to meaning andphonology) (Stoeckel et al., 2009). Thus, it may be that decreasedspatial orientation and semantic representation (symbol manipula-tion) could contribute to reduced cognitive abilities to perform organi-zational strategies in patients with OCD. This is in line with theoreticalapproaches that consider OCD to be associated with specific executivefailure of organizational strategies during encoding (Olley et al.,2007).
Increased connectivity (OCD N control; p b 0.05) was found in theRSNE auditory network in the right section of BA 43 (Fig. 2a). One possibleexplanation for this finding may lie in the difference in how the twocomparison groups react to the loud MRI setting. As OCD is typicallyclassified as an anxiety disorder (Stein et al., 2010), the extremelyloud and potentially frightening sounds that the MRI system makescould activate the auditory network differently in children with OCDthan in healthy matched children. The specific location of increasedactivity, BA 43, is believed to be responsible for the sensorimotor repre-sentation of the mouth (Bernal and Perdomo, 2008). One possible ex-planation for this is that OCD is often co-morbid with Tourette'ssyndrome (Mol Debes, 2012), with 20–30% of subjectswith OCDhavinga current or past history of facial tics (Franklin et al., 2012; Pauls et al.,1986), that are usually exacerbated by stress. It is thus plausible that
the increased stress in the fMRI environment is associated with morefrequent tics in the OCD group that was subsequently detected by in-creased BA 43 activation. Looking at sample demographics, however,revealed that only one subject from the OCD group presented withtics, while no subject from the control group did.
The difference between our findings and the 11 previous existingrs-fcMRI studies in adults and youth with OCD (Fitzgerald et al., 2010,2011; Fontenelle et al., 2012; Harrison et al., 2009; Jang et al., 2010;Kang et al., 2013; Li et al., 2012; Meunier et al., 2012; Sakai et al.,2010; Stern et al., 2012a, 2012b; Zhang, 2011) may be accounted forby the ICA approach taken by our group. For example, a CCA study ofthe DMN requires placement of a seed voxel or ROI in the PCC/precuneus (Buckner et al., 2008). Indeed, this approachwould probablynot introduce any issues in studies of adult subjects. However, whenstudying children and adolescents, there is the risk of only looking atpart of the DMN, as this network is broken into several smaller net-works in this age group (Fair et al., 2009). In contrast, the ICA methodallowed us to identify the RSNA, RSNB and RSNC (Fair et al., 2008;Kelly et al., 2009) as possible precursors to the adult DMN. CCAmethodsare also limited by the lack of accepted standards of selecting seedROIs for the DMN and issues of variability in inter-subject anatomyand anatomy–function correspondence (Ma et al., 2007). Furthermore,in many correlation studies, the global signal is often regressed out,
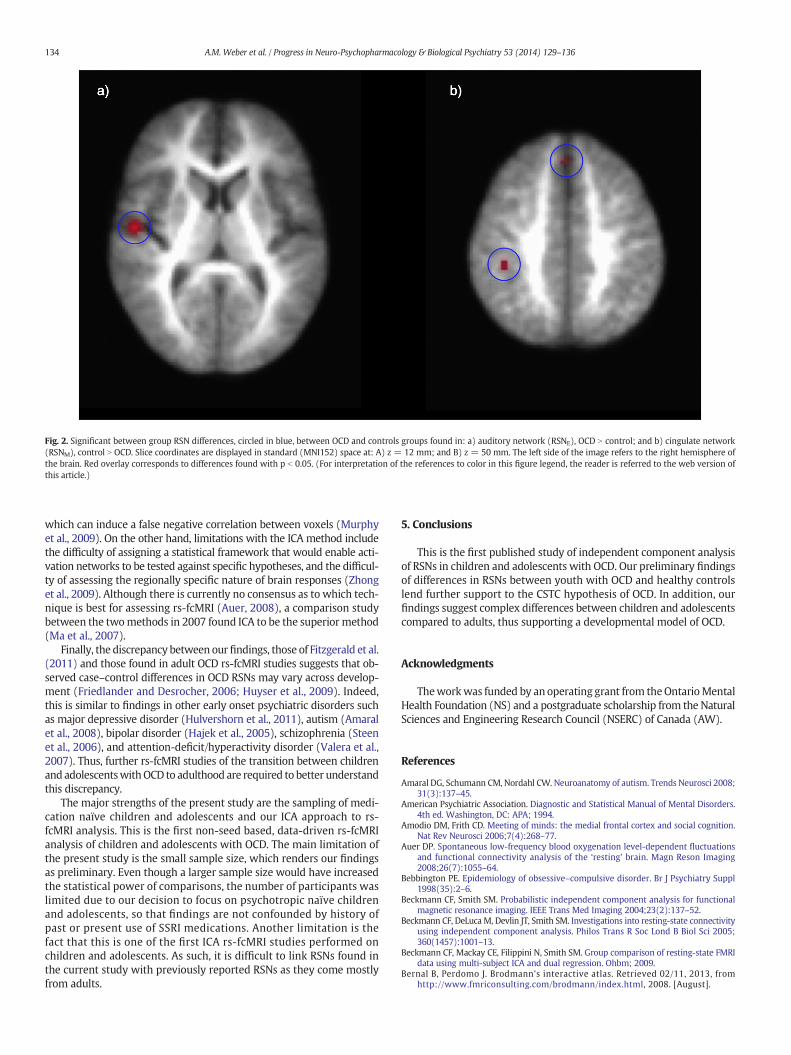
Fig. 2. Significant between group RSN differences, circled in blue, between OCD and controls groups found in: a) auditory network (RSNE), OCD N control; and b) cingulate network(RSNM), control N OCD. Slice coordinates are displayed in standard (MNI152) space at: A) z = 12 mm; and B) z = 50 mm. The left side of the image refers to the right hemisphere ofthe brain. Red overlay corresponds to differences found with p b 0.05. (For interpretation of the references to color in this figure legend, the reader is referred to the web version ofthis article.)
134 A.M. Weber et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 53 (2014) 129–136
which can induce a false negative correlation between voxels (Murphyet al., 2009). On the other hand, limitations with the ICA method includethe difficulty of assigning a statistical framework that would enable acti-vation networks to be tested against specific hypotheses, and the difficul-ty of assessing the regionally specific nature of brain responses (Zhonget al., 2009). Although there is currently no consensus as to which tech-nique is best for assessing rs-fcMRI (Auer, 2008), a comparison studybetween the twomethods in 2007 found ICA to be the superior method(Ma et al., 2007).
Finally, the discrepancy between ourfindings, those of Fitzgerald et al.(2011) and those found in adult OCD rs-fcMRI studies suggests that ob-served case–control differences in OCD RSNs may vary across develop-ment (Friedlander and Desrocher, 2006; Huyser et al., 2009). Indeed,this is similar to findings in other early onset psychiatric disorders suchas major depressive disorder (Hulvershorn et al., 2011), autism (Amaralet al., 2008), bipolar disorder (Hajek et al., 2005), schizophrenia (Steenet al., 2006), and attention-deficit/hyperactivity disorder (Valera et al.,2007). Thus, further rs-fcMRI studies of the transition between childrenand adolescentswithOCD to adulthood are required to better understandthis discrepancy.
The major strengths of the present study are the sampling of medi-cation naïve children and adolescents and our ICA approach to rs-fcMRI analysis. This is the first non-seed based, data-driven rs-fcMRIanalysis of children and adolescents with OCD. The main limitation ofthe present study is the small sample size, which renders our findingsas preliminary. Even though a larger sample size would have increasedthe statistical power of comparisons, the number of participants waslimited due to our decision to focus on psychotropic naïve childrenand adolescents, so that findings are not confounded by history ofpast or present use of SSRI medications. Another limitation is thefact that this is one of the first ICA rs-fcMRI studies performed onchildren and adolescents. As such, it is difficult to link RSNs found inthe current study with previously reported RSNs as they come mostlyfrom adults.
5. Conclusions
This is the first published study of independent component analysisof RSNs in children and adolescents with OCD. Our preliminary findingsof differences in RSNs between youth with OCD and healthy controlslend further support to the CSTC hypothesis of OCD. In addition, ourfindings suggest complex differences between children and adolescentscompared to adults, thus supporting a developmental model of OCD.
Acknowledgments
Theworkwas funded by an operating grant from theOntarioMentalHealth Foundation (NS) and a postgraduate scholarship from the NaturalSciences and Engineering Research Council (NSERC) of Canada (AW).
References
Amaral DG, Schumann CM, Nordahl CW. Neuroanatomy of autism. Trends Neurosci 2008;31(3):137–45.
American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders.4th ed. Washington, DC: APA; 1994.
Amodio DM, Frith CD. Meeting of minds: the medial frontal cortex and social cognition.Nat Rev Neurosci 2006;7(4):268–77.
Auer DP. Spontaneous low-frequency blood oxygenation level-dependent fluctuationsand functional connectivity analysis of the ‘resting’ brain. Magn Reson Imaging2008;26(7):1055–64.
Bebbington PE. Epidemiology of obsessive–compulsive disorder. Br J Psychiatry Suppl1998(35):2–6.
Beckmann CF, Smith SM. Probabilistic independent component analysis for functionalmagnetic resonance imaging. IEEE Trans Med Imaging 2004;23(2):137–52.
Beckmann CF, DeLuca M, Devlin JT, Smith SM. Investigations into resting-state connectivityusing independent component analysis. Philos Trans R Soc Lond B Biol Sci 2005;360(1457):1001–13.
Beckmann CF, Mackay CE, Filippini N, Smith SM. Group comparison of resting-state FMRIdata using multi-subject ICA and dual regression. Ohbm; 2009.
Bernal B, Perdomo J. Brodmann's interactive atlas. Retrieved 02/11, 2013, fromhttp://www.fmriconsulting.com/brodmann/index.html, 2008. [August].

135A.M. Weber et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 53 (2014) 129–136
Biswal B, Yetkin FZ, Haughton VM, Hyde JS. Functional connectivity in the motorcortex of resting human brain using echo-planar MRI. Magn Reson Med 1995;34(4):537–41.
Botvinick M, Nystrom LE, Fissell K, Carter CS, Cohen JD. Conflict monitoring versusselection-for-action in anterior cingulate cortex. Nature 1999;402(6758):179–81.
Botvinick MM, Cohen JD, Carter CS. Conflict monitoring and anterior cingulate cortex: anupdate. Trends Cogn Sci 2004;8(12):539–46.
Buckner RL, Andrews-Hanna JR, Schacter DL. The brain's default network: anatomy, func-tion, and relevance to disease. Ann N Y Acad Sci 2008;1124:1–38.
Bush G, Luu P, Posner MI. Cognitive and emotional influences in anterior cingulate cortex.Trends Cogn Sci 2000;4(6):215–22.
Bush G, Vogt BA, Holmes J, Dale AM, Greve D, Jenike MA, et al. Dorsal anterior cingulatecortex: a role in reward-based decision making. Proc Natl Acad Sci U S A 2002;99(1):523–8.
Chen S, Ross TJ, Zhan W, Myers CS, Chuang KS, Heishman SJ, et al. Group independentcomponent analysis reveals consistent resting-state networks across multiplesessions. Brain Res 2008;1239:141–51.
Chew SH, Ho JL. Hope: an empirical study of attitude toward the timing of uncertaintyresolution. J Risk Uncertain 1994;8(3):267–88.
Cox RW. AFNI: software for analysis and visualization of functional magnetic resonanceneuroimages. Comput Biomed Res Int J 1996;29(3):162–73.
Damoiseaux JS, Rombouts SA, Barkhof F, Scheltens P, Stam CJ, Smith SM, et al. Consistentresting-state networks across healthy subjects. Proc Natl Acad Sci U S A 2006;103(37):13848–53.
Fair DA, Cohen AL, Dosenbach NU, Church JA, Miezin FM, Barch DM, et al. The maturingarchitecture of the brain's default network. Proc Natl Acad Sci U S A 2008;105(10):4028–32.
Fair DA, Cohen AL, Power JD, Dosenbach NU, Church JA, Miezin FM, et al. Functional brainnetworks develop from a “local to distributed” organization. PLoS Comput Biol 2009;5(5):e1000381.
Fitzgerald KD, Stern ER, Angstadt M, Nicholson-Muth KC, Maynor MR, Welsh RC, et al.Altered function and connectivity of the medial frontal cortex in pediatric obsessive–compulsive disorder. Biol Psychiatry 2010;68(11):1039–47.
Fitzgerald KD,Welsh RC, Stern ER, AngstadtM, Hanna GL, Abelson JL, et al. Developmentalalterations of frontal–striatal–thalamic connectivity in obsessive–compulsive disor-der. J Am Acad Child Adolesc Psychiatry 2011;50(9):938–48. [e3].
Fontenelle L, Harrison B, Pujol J, Davey C, Fornito A, Bora E, et al. Brain functional connectivityduring induced sadness in patients with obsessive–compulsive disorder. J PsychiatryNeurosci 2012;37(4):231–40.
Franklin ME, Harrison JP, Benavides KL. Obsessive–compulsive and tic-related disorders.Child Adolesc Psychiatr Clin N Am 2012;21(3):555–71.
Friedlander L, Desrocher M. Neuroimaging studies of obsessive–compulsive disorder inadults and children. Clin Psychol Rev 2006;26(1):32–49.
Friston KJ, Frith CD, Liddle PF, Frackowiak RS. Functional connectivity: the principal-component analysis of large (PET) data sets. J Cereb Blood Flow Metab 1993;13(1):5–14.
Frith U, Frith CD. Development and neurophysiology of mentalizing. Philos Trans R SocLond B Biol Sci 2003;358(1431):459–73.
Garavan H, Ross TJ, Kaufman J, Stein EA. A midline dissociation between error-processingand response-conflict monitoring. Neuroimage 2003;20(2):1132–9.
Goodman WK, Price LH, Rasmussen SA, Riddle MA, Rapoport JL. Children's Yale–BrownObsessive Compulsive Scale (CY-BOCS); 1991.
Graybiel AM, Rauch SL. Toward a neurobiology of obsessive–compulsive disorder. Neuron2000;28(2):343–7.
Hajek T, Carrey N, Alda M. Neuroanatomical abnormalities as risk factors for bipolardisorder. Bipolar Disord 2005;7(5):393–403.
Harrison BJ, Soriano-Mas C, Pujol J, Ortiz H, Lopez-Sola M, Hernandez-Ribas R, et al.Altered corticostriatal functional connectivity in obsessive–compulsive disorder.Arch Gen Psychiatry 2009;66(11):1189–200.
Heine L, Soddu A, Gomez F, Vanhaudenhuyse A, Tshibanda L, Thonnard M, et al. Restingstate networks and consciousness: alterations of multiple resting state network con-nectivity in physiological, pharmacological, and pathological consciousness states.Front Psychol 2012;3:295.
Heyman I, Fombonne E, Simmons H, Ford T, Meltzer H, Goodman R. Prevalence ofobsessive–compulsive disorder in the British nationwide survey of child mentalhealth. Int Rev Psychiatry 2003;15(1–2):178–84.
Horwath E,WeissmanMM. The epidemiology and cross-national presentation of obsessive–compulsive disorder. Psychiatr Clin North Am 2000;23(3):493–507.
Hulvershorn LA, Cullen K, Anand A. Toward dysfunctional connectivity: a review of neu-roimaging findings in pediatric major depressive disorder. Brain Imaging Behav2011;5(4):307–28.
Huyser C, Veltman DJ, de Haan E, Boer F. Paediatric obsessive–compulsive disorder, aneurodevelopmental disorder? evidence from neuroimaging. Neurosci BiobehavRev 2009;33(6):818–30.
Jang JH, Kim JH, Jung WH, Choi JS, Jung MH, Lee JM, et al. Functional connectivity infronto-subcortical circuitry during the resting state in obsessive–compulsive disorder.Neurosci Lett 2010;474(3):158–62.
Johnson SC, Baxter LC,Wilder LS, Pipe JG, Heiserman JE, Prigatano GP. Neural correlates ofself-reflection. Brain 2002;125(Pt 8):1808–14.
Kang D, Jang JH, Han JY, Kim J, JungWH, Choi J, et al. Neural correlates of altered responseinhibition and dysfunctional connectivity at rest in obsessive–compulsive disorder.Prog Neuropsychopharmacol Biol Psychiatry 2013;40(C):340–6.
Kaufman J, Birmaher B, Brent D, Rao U, Flynn C, Moreci P, et al. Schedule for affectivedisorders and schizophrenia for school-age children-present and lifetime version(K-SADS-PL): initial reliability and validity data. J Am Acad Child Adolesc Psychiatry1997;36(7):980–8.
Kelly AM, Di Martino A, Uddin LQ, Shehzad Z, Gee DG, Reiss PT, et al. Development ofanterior cingulate functional connectivity from late childhood to early adulthood.Cereb Cortex 2009;19(3):640–57.
Kiehl KA, Liddle PF, Hopfinger JB. Error processing and the rostral anterior cingulate: anevent-related fMRI study. Psychophysiology 2000;37(2):216–23.
Kroger JK, Sabb FW, Fales CL, Bookheimer SY, Cohen MS, Holyoak KJ. Recruitment ofanterior dorsolateral prefrontal cortex in human reasoning: a parametric study of re-lational complexity. Cereb Cortex 2002;12(5):477–85.
Laird AR, Fox PM, Eickhoff SB, Turner JA, Ray KL, McKay DR, et al. Behavioral interpreta-tions of intrinsic connectivity networks. J Cogn Neurosci 2011;23(12):4022–37.
Li P, Li P, Li S, Li S, Dong Z, Dong Z, et al. Altered resting state functional connectivity patternsof the anterior prefrontal cortex in obsessive–compulsive disorder. Neuroreport 2012;23(11):681–6.
Luo Q, Perry C, Peng D, Jin Z, Xu D, Ding G, et al. The neural substrate of analogical reasoning:an fMRI study. Brain Res Cogn Brain Res 2003;17(3):527–34.
Ma L, Wang B, Chen X, Xiong J. Detecting functional connectivity in the resting brain: acomparison between ICA and CCA. Magn Reson Imaging 2007;25(1):47–56.
McKeownMJ, Makeig S, Brown GG, Jung TP, Kindermann SS, Bell AJ, et al. Analysis of fMRIdata by blind separation into independent spatial components. Hum Brain Mapp1998;6(3):160–88.
Melloni M, Urbistondo C, Sedeno L, Gelormini C, Kichic R, Ibanez A. The extended fronto-striatal model of obsessive compulsive disorder: convergence from event-related po-tentials, neuropsychology and neuroimaging. Front Hum Neurosci 2012;6:259.
Menon V, Uddin LQ. Saliency, switching, attention and control: a network model of insulafunction. Brain Struct Funct 2010;214(5–6):655–67.
Meunier D, Ersche KD, Craig KJ, Fornito A, Merlo-Pich E, Fineberg NA, et al. Brain functionalconnectivity in stimulant drug dependence and obsessive–compulsive disorder.Neuroimage 2012;59(2):1461–8.
Mol Debes NM. Co-morbid disorders in Tourette syndrome. Behav Neurol 2012;26(1):7–14.
Murphy K, Birn RM, Handwerker DA, Jones TB, Bandettini PA. The impact of global signalregression on resting state correlations: are anti-correlated networks introduced?Neuroimage 2009;44(3):893–905.
Nielsen FA, Balslev D, Hansen LK. Mining the posterior cingulate: segregation betweenmemory and pain components. Neuroimage 2005;27(3):520–32.
Ochsner KN, Beer JS, Robertson ER, Cooper JC, Gabrieli JD, Kihsltrom JF, et al. The neuralcorrelates of direct and reflected self-knowledge. Neuroimage 2005;28(4):797–814.
Olley A, Malhi G, Sachdev P. Memory and executive functioning in obsessive–compulsivedisorder: a selective review. J Affect Disord 2007;104(1–3):15–23.
Pauls DL, Towbin KE, Leckman JF, Zahner GE, Cohen DJ. Gilles de la Tourette's syndromeand obsessive–compulsive disorder. Evidence supporting a genetic relationship.Arch Gen Psychiatry 1986;43(12):1180–2.
Paulus MP, Frank LR. Anterior cingulate activity modulates nonlinear decision weightfunction of uncertain prospects. Neuroimage 2006;30(2):668–77.
Raichle ME, MacLeod AM, Snyder AZ, Powers WJ, Gusnard DA, Shulman GL. A defaultmode of brain function. Proc Natl Acad Sci U S A 2001;98(2):676–82.
Rapoport JL, Inoff-Germain G, Weissman MM, Greenwald S, Narrow WE, Jensen PS, et al.Childhood obsessive–compulsive disorder in the NIMH MECA study: parent versuschild identification of cases. Methods Epidemiol Child Adolesc Ment Disord J AnxietyDisord 2000;14(6):535–48.
Rosazza C, Minati L. Resting-state brain networks: literature review and clinical applica-tions. Neurol Sci 2011;32(5):773–85.
Ruscio AM, Stein DJ, ChiuWT, Kessler RC. The epidemiology of obsessive–compulsive dis-order in the national comorbidity survey replication. Mol Psychiatry 2010;15(1):53–63.
Sakai Y, Narumoto J, Nishida S, Nakamae T, Yamada K, Nishimura T, et al. Corticostriatalfunctional connectivity in non-medicated patients with obsessive–compulsive disor-der. Eur Psychiatry 2010;1–7.
Saxena S, Brody AL, Schwartz JM, Baxter LR. Neuroimaging and frontal–subcortical circuitryin obsessive–compulsive disorder. Br J Psychiatry Suppl 1998(35):26–37.
Saxena S, Bota RG, Brody AL. Brain–behavior relationships in obsessive–compulsive disorder.Semin Clin Neuropsychiatry 2001;6(2):82–101.
Scahill L, Riddle MA, McSwiggin-Hardin M, Ort SI, King RA, GoodmanWK, et al. Children'sYale–Brown Obsessive Compulsive Scale: reliability and validity. J Am Acad ChildAdolesc Psychiatry 1997;36(6):844–52.
Smith SM, Fox PT, Miller KL, Glahn DC, Fox PM, Mackay CE, et al. Correspondence of thebrain's functional architecture during activation and rest. Proc Natl Acad Sci U S A2009;106(31):13040–5.
Steen RG, Mull C, McClure R, Hamer RM, Lieberman JA. Brain volume in first-episodeschizophrenia: systematic review and meta-analysis of magnetic resonance imagingstudies. Br J Psychiatry J Ment Sci 2006;188:510–8.
Stein DJ, Fineberg NA, Bienvenu OJ, Denys D, Lochner C, Nestadt G, et al. Should OCD beclassified as an anxiety disorder in DSM-V? Depress Anxiety 2010;27(6):495–506.
Stern ER, Fitzgerald KD, Welsh RC, Abelson JL, Taylor SF. Resting-state functional connec-tivity between fronto-parietal and default mode networks in obsessive–compulsivedisorder. PLoS One 2012a;7(5):e36356.
Stern ER, Welsh RC, Gonzalez R, Fitzgerald KD, Abelson JL, Taylor SF. Subjective uncertaintyand limbic hyperactivation in obsessive–compulsive disorder. Hum Brain Mapp2012b;34(8):1956–70.
Stevens MC, Pearlson GD, Calhoun VD. Changes in the interaction of resting-state neuralnetworks from adolescence to adulthood. Hum Brain Mapp 2009;30(8):2356–66.
Stoeckel C, Gough PM, Watkins KE, Devlin JT. Supramarginal gyrus involvement in visualword recognition. Cortex 2009;45(9):1091–6.
Storch EA. The clinician rated CY-BOCS scale is more sensitive than the self-reported LOI-CVin children and adolescents with obsessive compulsive disorder. Evid Based MentHealth 2006;9(3):67.

136 A.M. Weber et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 53 (2014) 129–136
Supekar K, Uddin LQ, Prater K, Amin H, Greicius MD, Menon V. Development of functionaland structural connectivity within the default mode network in young children.Neuroimage 2010;52(1):290–301.
Valderhaug R, Ivarsson T. Functional impairment in clinical samples of Norwegian andSwedish children and adolescents with obsessive–compulsive disorder. Eur ChildAdolesc Psychiatry 2005;14(3):164–73.
Valera EM, Faraone SV, Murray KE, Seidman LJ. Meta-analysis of structural imagingfindings in attention-deficit/hyperactivity disorder. Biol Psychiatry 2007;61(12):1361–9.
Volz KG, Schubotz RI, von Cramon DY. Variants of uncertainty in decision-making andtheir neural correlates. Brain Res Bull 2005;67(5):403–12.
Webster M. Randomise/theory. Retrieved 01/03, 2013, from http://fsl.fmrib.ox.ac.uk/fsl/fslwiki/Randomise/Theory, 2012.
Zhang T. Abnormal small-world architecture of top-down control networks in obsessive–compulsive disorder. J Psychiatry Neurosci 2011;36(1):23–31.
Zhong Y, Wang H, Lu G, Zhang Z, Jiao Q, Liu Y. Detecting functional connectivity in fMRIusing PCA and regression analysis. Brain Topogr 2009;22(2):134–44.


















