
A possible neurotransmitter role for CGRP in a hair-cell sensory organ
5
Brain Research, 419 (1987) 347-351 347 Elsevier BRE 22458 A possible neurotransmitter role for CGRP in a hair-cell sensory organ J.C. Adams 1, E.A. Mroz 2'3 and W.F. Sewell 2'4 1Department of Otolaryngology and Communicative Sciences, Medical University of South Carolina, Charleston, SC 29425 (U.S.A.), 2Eaton-Peabody Laboratory of Auditory Physiology, MassachusettsEye and Ear Infirmary, and Department of Otolaryngology, Harvard Medical School, 3Departmentof Physiology, Harvard Medical School, 4programin Neuroscience, Harvard Medical School, Boston, MA 02114 (U.S.A.) (Accepted 26 May 1987) Key words: Calcitonin gene-related peptide; Lateral line organ; Hair cell; Neurotransmitter; Efferent nerve We report that calcitonin gene-related peptide (CGRP) increases the discharge rate of afferent fibers innervating hair cells in the lateral line organ of Xenopus laevis. We have localized CGRP-like immunoreactivityin small, presumably efferent, fibers innervating the lateral line organ. In addition to providing evidence for a neurotransmitter role for CGRP in a sensory system, these results may help explain the non-cholinergic excitatory effect seen with efferent stimulation in this and other hair cell organs such as the inner ear. Calcitonin gene-related peptide (CGRP) is a re- cently discovered, 37-amino-acid peptide 3'5'19'21 that has potent effects on the heart 27 and on smooth mus- cle 4,18. The distribution of CGRP and its binding sites in the nervous system2,2°'28,3°,31 suggests that it may be important in sensory function. Potassium-stimu- lated, calcium-dependent release of CGRP from cul- tured rat trigeminal ganglion cells has been found 15. There have been, however, no reports of effects of CGRP on neuronal firing rates in sensory systems. We examined the effects of CGRP and related peptides on the firing rates of afferent fibers inner- vating hair cells of the lateral line organs of 18 Afri- can clawed frogs (Xenopus laevis). Frogs were anes- thetized by cooling and a portion of the skin contain- ing the middle-lateral row of stitches were removed. The inner surface of the skin was continually super- fused with a salt solution (120 mM NaC1, 3.5 mM KCI, 1.5 mM CaCI2, 5.5 mM glucose, 20 mM Hepes, pH adjusted to 7.5 with NaOH). The activity of up to 6 afferent fibers was recorded simultaneously with a wire electrode in contact with the whole nerve trunk. All branches of the trunk except those to 3 stitches were disrupted. Peptides (obtained from Sigma) were dissolved in the fluid used to bathe the prepara- tion, and were applied by diverting the flow of super- fusion fluid through a loop containing 100 pl of pep- tide solution. For each applied sample, the percen- tage change in discharge rate was calculated by com- paring the rate during a 70-s control period immedia- tely preceding the application to that during a 40-s period during peak effects of sample application. De- tails of these methods have been presented else- where 29. Synthetic CGRP, whether of rat or human se- quence, increased the discharge rates of afferent nerve fibers of the lateral line organ (Fig. 1). Re- sponses to rat CGRP were seen at doses as low as 40 nM in some preparations, but most preparations re- sponded over concentration ranges of 100 nM to 10 ~M (Fig. 2). Synthetic human CGRP generally pro- duced a response that was about two-thirds that of the rat CGRP (data not shown). Unlike CGRP, calcitonin did not increase firing rates of lateral line afferents. Human calcitonin, a 32- amino-acid peptide, was inactive at doses up to 20 /~M (Fig. 3). Salmon calcitonin, which may share some of the biological activities of CGRP in mam- mals 8'32, had no effect on discharge rate at low con- centrations, and at higher concentrations (10/tM) de- 0006-8993/87/$03.50 © 1987 Elsevier Science Publishers B.V. (Biomedical Division)
Transcript of A possible neurotransmitter role for CGRP in a hair-cell sensory organ

Brain Research, 419 (1987) 347-351 347 Elsevier
BRE 22458
A possible neurotransmitter role for CGRP in a hair-cell sensory organ
J.C. Adams 1, E .A. Mroz 2'3 and W.F. Sewell 2'4
1Department of Otolaryngology and Communicative Sciences, Medical University of South Carolina, Charleston, SC 29425 (U.S.A.), 2Eaton-Peabody Laboratory of Auditory Physiology, Massachusetts Eye and Ear Infirmary, and Department of Otolaryngology,
Harvard Medical School, 3Department of Physiology, Harvard Medical School, 4program in Neuroscience, Harvard Medical School, Boston, MA 02114 (U.S.A.)
(Accepted 26 May 1987)
Key words: Calcitonin gene-related peptide; Lateral line organ; Hair cell; Neurotransmitter; Efferent nerve
We report that calcitonin gene-related peptide (CGRP) increases the discharge rate of afferent fibers innervating hair cells in the lateral line organ of Xenopus laevis. We have localized CGRP-like immunoreactivity in small, presumably efferent, fibers innervating the lateral line organ. In addition to providing evidence for a neurotransmitter role for CGRP in a sensory system, these results may help explain the non-cholinergic excitatory effect seen with efferent stimulation in this and other hair cell organs such as the inner ear.
Calcitonin gene-related peptide (CGRP) is a re- cently discovered, 37-amino-acid peptide 3'5'19'21 that
has potent effects on the heart 27 and on smooth mus- cle 4,18. The distribution of CGRP and its binding sites in the nervous system 2,2°'28,3°,31 suggests that it may
be important in sensory function. Potassium-stimu-
lated, calcium-dependent release of CGRP from cul- tured rat trigeminal ganglion cells has been found 15.
There have been, however, no reports of effects of
CGRP on neuronal firing rates in sensory systems. We examined the effects of CGRP and related
peptides on the firing rates of afferent fibers inner- vating hair cells of the lateral line organs of 18 Afri- can clawed frogs (Xenopus laevis). Frogs were anes- thetized by cooling and a portion of the skin contain- ing the middle-lateral row of stitches were removed. The inner surface of the skin was continually super- fused with a salt solution (120 mM NaC1, 3.5 mM
KCI, 1.5 mM CaCI2, 5.5 mM glucose, 20 mM Hepes, pH adjusted to 7.5 with NaOH). The activity of up to 6 afferent fibers was recorded simultaneously with a wire electrode in contact with the whole nerve trunk. All branches of the trunk except those to 3 stitches were disrupted. Peptides (obtained from Sigma) were dissolved in the fluid used to bathe the prepara-
tion, and were applied by diverting the flow of super- fusion fluid through a loop containing 100 pl of pep-
tide solution. For each applied sample, the percen- tage change in discharge rate was calculated by com-
paring the rate during a 70-s control period immedia- tely preceding the application to that during a 40-s
period during peak effects of sample application. De- tails of these methods have been presented else- where 29.
Synthetic CGRP, whether of rat or human se-
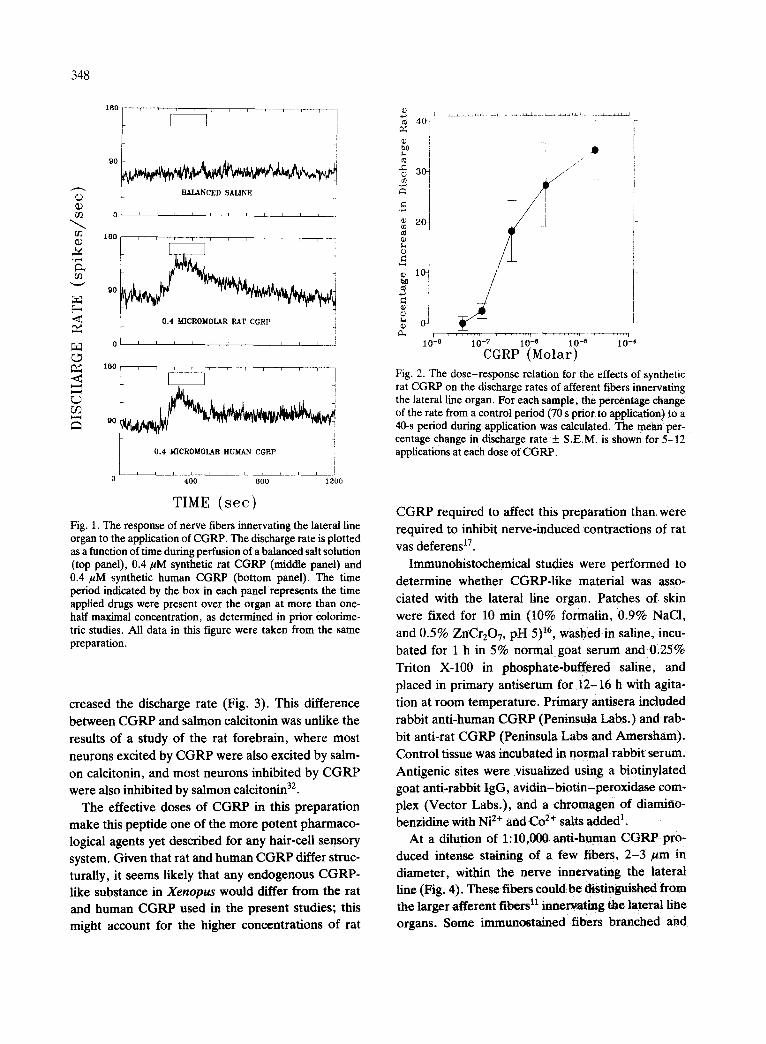
quence, increased the discharge rates of afferent nerve fibers of the lateral line organ (Fig. 1). Re-
sponses to rat CGRP were seen at doses as low as 40 nM in some preparations, but most preparations re- sponded over concentration ranges of 100 nM to 10 ~M (Fig. 2). Synthetic human CGRP generally pro- duced a response that was about two-thirds that of the rat CGRP (data not shown).
Unlike CGRP, calcitonin did not increase firing rates of lateral line afferents. Human calcitonin, a 32- amino-acid peptide, was inactive at doses up to 20 /~M (Fig. 3). Salmon calcitonin, which may share some of the biological activities of CGRP in mam- mals 8'32, had no effect on discharge rate at low con- centrations, and at higher concentrations (10/tM) de-
0006-8993/87/$03.50 © 1987 Elsevier Science Publishers B.V. (Biomedical Division)
348
C) 0)
1 B O
G~ 9O
180
~3 90
1°0 t E . . . . . . . -i!
J
o l l l ' l l l l l I , _ .
0.4 MICROMOLAR RAT C G R P i i
I I t I I ¢ I ~ r I I i
I , I I I t ] { [ I ] - - -
I ]
0 . 4 M I C R O M O L A R H U M A N C G R P
I I i I i I i J i _ . ~ . . . . .
0 4 0 0 8 0 0 1 2 0 0
TIME ( see ) Fig. 1. The response of nerve fibers innervating the lateral line organ to the application of CGRP. The discharge rate is plotted as a function of time during perfusion of a balanced salt solution (top panel), 0.4/~M synthetic rat CGRP (middle panel) and 0.4 /~M synthetic human CGRP (bottom panel). The time period indicated by the box in each panel represents the time applied drugs were present over the organ at more than one- half maximal concentration, as determined in prior colorime- tric studies. All data in this figure were taken from the same preparation.
creased the discharge rate (Fig. 3). This difference between CGRP and salmon calcitonin was unlike the results of a study of the rat forebrain, where most neurons excited by CGRP were also excited by salm- on calcitonin, and most neurons inhibited by CGRP were also inhibited by salmon calcitonin 32.
The effective doses of CGRP in this preparation make this peptide one of the more potent pharmaco- logical agents yet described for any hair-cell sensory system. Given that rat and human CGRP differ struc- turally, it seems likely that any endogenous CGRP- like substance in Xenopus would differ from the rat and human CGRP used in the present studies; this might account for the higher concentrations of rat
.~ 3 f /
/ " I
1
[
1 , V ~ . . . . . . . . r . . . . . . . t - ' . . . . . . . q . . . . . . . . I
10-8 10-7 10-e 10-5 i0-4 CGRP ( M o l a r )
Fig. 2. The dose-response relation for the effects of synthetic rat CGRP on the discharge rates of afferent fibers innervating the lateral line organ. For each sample, the percentage change: of the rate from a control period (70 s prior to application) to a 40-s period during application was calculated. R e mehn per- centage change in discharge rate _+ S.E.M. is shown for 5-12 applications at each dose of CGRP.
CGRP required to affect this preparation than were required to inhibit nerve-induced contractions of rat vas deferens 17.
Immunohistochemical studies were performed to determine whether CGRP-Iike material was asso- ciated with the lateral line organ. Patches of skin were fixed for 10 min (10% formalin, 0.9% NaC1.
and 0.5% ZnCr207, pH 5) 16, washed in saline, incu- bated for 1 h in 5% normal goat serum and 0.25% Triton X-100 in phosphate-buffered saline, and placed in primary antiserum for 12-16 h with agita- tion at room temperature. Primary antisera included rabbit anti-human CGRP (Peninsula Labs.) and rab- bit anti-rat CGRP (Peninsula Labs and Amersham). Control tissue was incubated in normal rabbit serum. Antigenic sites were visualized using a biotinylated goat anti-rabbit IgG, avidin-biotin-peroxidase com- plex (Vector Labs.), and a chromagen of diamino- benzidine with Ni 2+ and Co 2+ salts added 1.
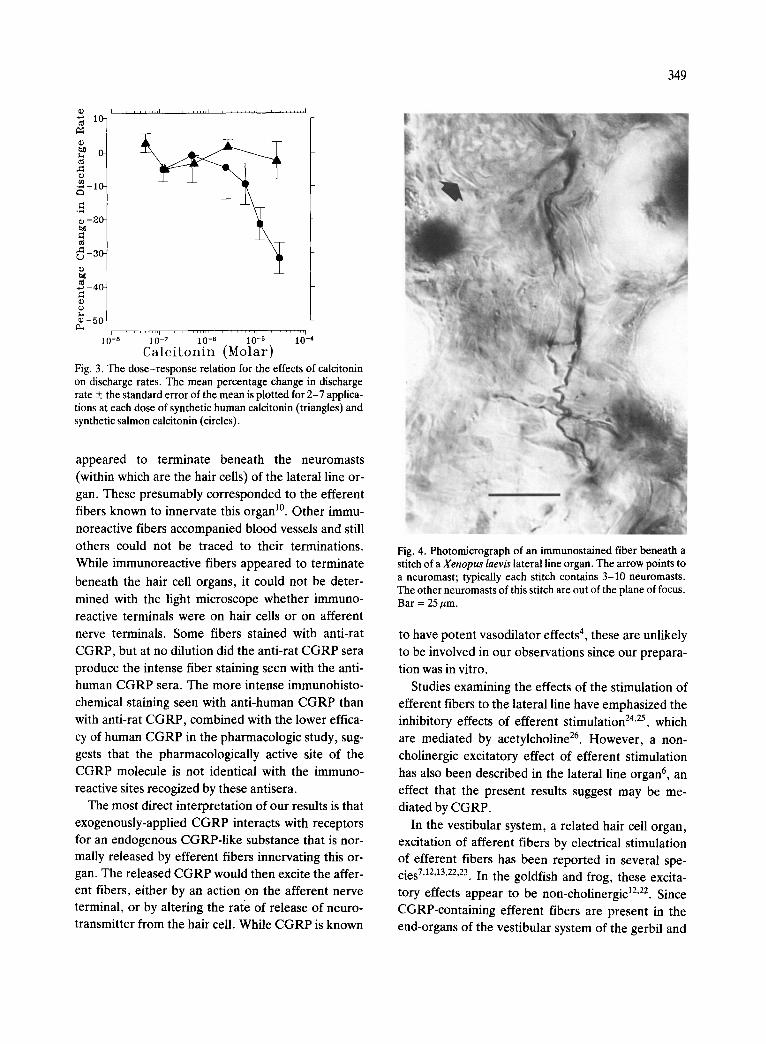
At a dilution of 1:10,000 anti-human CGRP pro- duced intense staining of a few fibers, 2-3 am in diameter, within the nerve innervating the lateral line (Fig. 4). These fibers could be distinguished from the larger afferent fibers 11 innervating the lateral line organs. Some immunostained fibers branched and
. . . . . . I . . . . . . . . I . . . . . . . . I . . . . . . . . I Q) 10.
0-
¢9 .~-10.
-20 .
t~
-30-
-~ -40 .
o~ - 5 0 .
349
i0r_8 . . . . . . . . [ . . . . . . . . ~ . . . . . . . . r . . . . . . . . 10-7 10-6 10-5 10-4
C a l c i t o n i n ( M o l a r ) Fig. 3. The dose-response relation for the effects of calcitonin on discharge rates. The mean percentage change in discharge rate + the standard error of the mean is plotted for 2-7 applica- tions at each dose of synthetic human calcitonin (triangles) and synthetic salmon calcitonin (circles).
appeared to terminate beneath the neuromasts
(within which are the hair cells) of the lateral line or-
gan. These presumably corresponded to the efferent fibers known to innervate this organ 1°. Other immu-
noreactive fibers accompanied blood vessels and still
others could not be traced to their terminations.
While immunoreactive fibers appeared to terminate
beneath the hair cell organs, it could not be deter-
mined with the light microscope whether immuno-
reactive terminals were on hair cells or on afferent
nerve terminals. Some fibers stained with anti-rat
CGRP, but at no dilution did the anti-rat C G R P sera
produce the intense fiber staining seen with the anti-
human C G R P sera. The more intense immunohisto-
chemical staining seen with anti-human C G R P than
with anti-rat CGRP, combined with the lower effica-
cy of human C G R P in the pharmacologic study, sug-
gests that the pharmacologically active site of the
CGRP molecule is not identical with the immuno- reactive sites recogized by these antisera.
The most direct interpretation of our results is that
exogenously-applied C G R P interacts with receptors for an endogenous CGRP-like substance that is nor- mally released by efferent fibers innervating this or- gan. The released C G R P would then excite the affer- ent fibers, either by an action on the afferent nerve terminal, or by altering the rate of release of neuro- transmitter from the hair cell. While C G R P is known
Fig. 4. Photomicrograph of an immunostained fiber beneath a stitch of a Xenopus laevis lateral line organ. The arrow points to a neuromast; typically each stitch contains 3-10 neuromasts. The other neuromasts of this stitch are out of the plane of focus. Bar = 25/~m.
to have potent vasodilator effects 4, these are unlikely
to be involved in our observations since our prepara-
tion was in vitro.
Studies examining the effects of the stimulation of efferent fibers to the lateral line have emphasized the inhibitory effects of efferent stimulation 24,25, which
are mediated by acetylcholine 26. However , a non-
cholinergic excitatory effect of efferent stimulation
has also been described in the lateral line organ 6 , an
effect that the present results suggest may be me- diated by CGRP.
In the vestibular system, a related hair cell organ, excitation of afferent fibers by electrical stimulation of efferent fibers has been reported in several spe- cies 7'12'13'22'23, In the goldfish and frog, these excita-
t o r y effects appear to be non-cholinergic 12,22. Since
CGRP-containing efferent fibers are present in the end-organs of the vestibular system of the gerbil and

350
rat (J. Adams , unpubl ished observat ions) , it is possi-
ble that these exci tatory effects may be media ted by
CGRP.
In the audi tory system, also a re la ted hair cell or-
gan, CGRP-I ike immunosta in ing has been repor ted in the central 2'28 and per iphera l t4,33 processes of the
efferent fibers of the lateral ol ivocochlear system 9.
The lack of reports describing excitat ion of audi tory
afferent fibers by st imulat ion of audi tory efferents
may be due to the ex t reme technical difficulty of
s t imulat ing the lateral efferent system while exclud-
ing the possibili ty of s imultaneously s t imulat ing the
inhibi tory ipsi lateral media l ol ivocochlear efferent fi-
bers. The excitatory effects of C G R P in the hair cell
system of the lateral line suggests that a t tent ion
should be paid to possible excitatory effects of this
lateral audi tory efferent system.
We wish to thank A. Mendez for technical assist-
ance. This work was suppor ted by Grants NS 21307,
NS 19097, and NS 13126 from the Nat ional Inst i tutes
of Heal th .
1 Adams, J.C., Heavy metal intensification of DAB-based HRP reaction product, J. Histochem. Cytochem., 29 (1981) 775.
2 Adams, J., Cells of origin of cochlear efferents in human, Abstr. Midwinter Meeting Assoc. for Research in Otolaryn- gology, 9 (1986) 5.
3 Amara, S.G., Jonas, V., Rosenfeld, M.G., Ong, E.S. and Evans, R.M., Alternative RNA processing in calcitonin gene expression generates mRNA encoding different poly- peptide products, Nature (London), 298 (1982) 240-244.
4 Brain, S.D., Williams, T.J., Tippins, J.R., Morris, H.R. and Maclntyre, I., Calcitonin gene-related peptide is a po- tent vasodilator, Nature (London), 313 (1985) 54-56.
5 Craig, R.K., Hall, L., Edbrooke, M.R., Allison, J. and Maclntyre, I., Partial nucleotide sequence of human calci- tonin precursor mRNA identifies flanking cryptic peptides, Nature (London), 295 (1982) 345-347.
6 Flock, A. and Russell, I.J., The postsynaptic action of ef- ferent fibres in the lateral line organ of the burbot Lota lota, J. Physiol. (London), 235 (1973) 591-605.
7 Goldberg J.M. and Fernandez, C., Efferent vestibular sys- tem in the squirrel monkey: anatomical location and influ- ence on afferent activity, J. Neurophysiol., 43 (1980) 986-1025.
8 Goltzman, D. and Mitchel, J., Interactions of calcitonin and calcitonin gene-related peptide at receptor sites in tar- get tissues, Science, 227 (1985) 1343-1345.
9 Guinan, J.J., Warr, W.B. and Norris, B.A., Topographic organization of the olivocochlear projections from the lat- eral and medial zones of the superior olivary complex, J. Comp. Neurol., 221 (1983) 358-370.
10 Harris, G.G. and Flock, A., Spontaneous and evoked ac- tivity from the Xenopus laevis lateral line. In P. Cahn (Ed.), Lateral Line Detectors, Indiana University Press, Bloom- ington, 1967, pp. 135-162.
11 Harris, G.G. and Milne, D.C., Input-output characteristics of the lateral line sense organs of Xenopus laevis, J. Acoust. Soc. Am., 40 (1%5) 32-42.
12 Hartmann, R. and Klinke, R., Efferent activity in the gold- fish vestibular nerve and its influence on afferent activity, Pfliigers Arch., 388 (1980) 123-128.
13 Highstein, S.M. and Baker, R., Action of the efferent ves- tibular system on primary afferents in the toadfish Opsanus tau, J. Neurophysiol., 54 (1985) 370-384.
14 Kita#ri, M., Yamashita, T., Tohyama, Y., Kumazawa, T., Takeda, Y., Kawasaki, Y., Matsunaga, T., Grigis, S., Hill-
yard, C.J., Maclntyre, I., Emson, P.C., Shiosaka, S. and Tohyama, M., Localization of calcitonin gene-related pep- tide in the organ of Corti of the rat: an immunohistochemi- cal study, Brain Research, 358 (1985) 394-397.
15 Mason, R.T., Peterfreund, R.A., Sawchenko, P.E., Corri- gan, A.Z., Rivier, J.E. and Vale, W.W., Release of the predicted calcitonin gene-related peptide from cultured rat trigeminal ganglion cells, Nature (London), 308 (1984) 653-655,
16 Mugnaini, E. and Dahl, A., Zinc-aldehyde fixation for light-microscopic immunocytochemistry of nervous tissues, J. Histochem. Cytochem., 31 (1983) 1435-1438.
17 Ohhashi, T. and Jacobowitz, D.M., Effect of calcitonin gene-related peptide on the neuroeffeetor mechanism of sympathetic nerve terminals in rat vas deferens, Peptides, 4 (1985) 987-991.
18 Palmer, J.M., Schemann. K., Tamura, K. and Wood. J.D.. Calcitonin gene-related peptide mimics slow synaptic exci- tation in myenterie neurons of guinea pig small intestine_ Soc. Neurosci. Abstr., 12 (19861 1495.
19 Rosenfeld, M.G., Amara, S.G.. Roos, B.A., Ong, E.S. and Evans, R. M., Altered expression of the calcitonin gene associated with RNA polymorphism. Nature (Lon- don), 290 (1981) 63-65.
20 Rosenfeld, M.G., Mermod, J.J., Amara, S.G., Swanson. L.W., Sawchenko, P.E., Rivier, J.. Vale, W.W. and Evans, R.M., Production of novel neuropeptide encoded by the caleitonin gene via tissue specific RNA processing, Nature (London), 304 (1983) 129-135
21 Rosenfeld. M.G., Lin, C.R., Amara, S.G., Stolarsky, L.o Roos, B.A., Ong, E.S. and Evans, R.M., Calcitonin mRNA polymorphism: peptide switching associated with alternative RNA splicing events, Proc. Natl. Acad. Sci. U.S.A., 79 (1982) 1717-1721
22 Rossi, M.L., Prigioni, I., VaUi, P. and Caselli, C., Activa- tion of the efferent system in the isolated frog labyrinth: ef- fects on the afferent EPSPs and spike discharge recorded from single fibers of the posterior nerve, Brain Research. 185 (1980) 125-137.
23 Rossi. M.L., Valti, P. and Caselli, C., Postsynaptic poten- tials recorded from afferent nerve fibres of the posterior semicircular canal in the frog, Brain Research, 135 (1977) 67-75.
24 Russell, I.J., Influence of efferent fibres on a receptor, Na- ture (London), 219 (1968) 177-178. ~
25 Russell, I.J., The role of the lateral line efferent system in

Xenopus laevis, J. Exp. Biol., 54 (1971) 621-641. 26 Russell, I.J., The pharmacology of efferent synapses in the
lateral-line system of Xenopus laevis, J. Exp. Biol., 54 (1971) 643-658.
27 Saito, A., Kimura, S. and Goto, K., Calcitonin gene-re- lated peptide as potential neurotransmitter in guinea pig right atrium, Am. J. Physiol., 250 (1986) H693-698.
28 Schweitzer, L.F., Lu, S.M., Dawbarn, D. and Cant, N.B., Calcitonin gene-related peptide immunoreactivity in the superior olivary complex of the cat and rat: a specific label for the lateral olivocochlear system, Soc. Neurosci. Abstr., 11 (1985) 1051.
29 Sewell, W. and Mroz, E., Neuroactive substances in inner ear extracts, J. Neurosci., in press.
30 Skofitsch, G. and Jacobowitz, D.M., Autoradiographic dis-
351
tribution of 125I-calcitonin gene-related peptide binding sites in the rat central nervous system, Peptides, 4 (1985) 975-986.
31 Tschopp, F.A., Henke, H., Petermann, J., Tobler, P., Jan- zer, R., H6kfelt, T., Lundberg, J.M., Cuello, C. and Fisch- er, J., Calcitonin gene-related peptide and its binding sites in the human central nervous system and pituitary, Proc. Natl. Acad. Sci. U.S.A., 82 (1985) 248-252.
32 Twery, M.J. and Moss, R.L., Calcitonin and calcitonin gene-related peptide alter the excitability of neurons in rat forebrain, Peptides, 6 (1985) 373-378.
33 Vetter, D.E., Adams, J.C. and Mugnaini, E., A dual effer- ent GABAergic projection to the rat cochlea, Soc. Neuro- sci. Abstr., 12 (1986) 779.


















