
Uncovering Higher-Taxon Diversification Dynamics from Clade ...

Novel Clade of Alphaproteobacterial Endosymbionts Associated withStinkbugs and Other Arthropods
Yu Matsuura,a,b Yoshitomo Kikuchi,c Xian Ying Meng,a Ryuichi Koga,a and Takema Fukatsua
National Institute of Advanced Industrial Science and Technology (AIST), Tsukuba, Japana; Graduate School of Life and Environmental Sciences, University of Tsukuba,Tsukuba, Japanb; and National Institute of Advanced Industrial Science and Technology (AIST), Hokkaido, Japanc
Here we report a novel clade of secondary endosymbionts associated with insects and other arthropods. Seed bugs of the genusNysius (Hemiptera: Lygaeidae) harbor the primary gammaproteobacterial symbiont Schneideria nysicola within a pair of bacte-riomes in the abdomen. Our survey of Nysius species for their facultative bacterial associates consistently yielded a novel type ofalphaproteobacterial 16S rRNA gene sequence in addition to those of Wolbachia. Diagnostic PCR survey of 343 individuals rep-resenting 24 populations of four Nysius species revealed overall detection rates of the alphaproteobacteria at 77.6% in Nysiusplebeius, 87.7% in Nysius sp. 1, 81.0% in Nysius sp. 2, and 100% in Nysius expressus. Further survey of diverse stinkbugs repre-senting 24 families, 191 species, and 582 individuals detected the alphaproteobacteria from an additional 12 species representingsix families. Molecular phylogenetic analysis showed that the alphaproteobacteria from the stinkbugs form a distinct and coher-ent monophyletic group in the order Rickettsiales together with several uncharacterized endosymbionts from fleas and ticks. Thealphaproteobacterial symbiont clade was allied to bacterial clades such as the endosymbionts of acanthamoebae, the endosymbi-onts of cnidarians, and Midichloria spp., the mitochondrion-associated endosymbionts of ticks. In situ hybridization and elec-tron microscopy identified small filamentous bacterial cells in various tissues of N. plebeius, including the bacteriome and ovary.The concentrated localization of the symbiont cells at the anterior pole of oocytes indicated its vertical transmission routethrough host insect generations. The designation “Candidatus Lariskella arthropodarum” is proposed for the endosymbiontclade.
Diverse organisms are in symbiotic association with multiplebacterial species. Many insects are associated with an obligate
symbiotic bacterium, which is referred to as the “primary” sym-biont. The primary symbiont generally plays important biologicalroles for growth, survival, and/or reproduction of its host, andtherefore its infection is fixed in the host populations. Frequently,these insects also possess one or a few additional facultative bac-terial associates, which are referred to as “secondary” symbionts.The secondary symbionts are generally not essential for their hostand usually exhibit partial infection frequencies in the host pop-ulations (46, 49).
Plant-sucking stinkbugs (Hemiptera: Heteroptera: Pentato-momorpha) are also frequently associated with symbiotic bacte-ria. While several atypical stinkbugs of the family Lygaeidae areassociated with a primary endocellular bacterial symbiont in thebacteriome (39, 40, 41, 43), the majority of stinkbugs are associ-ated with a primary extracellular bacterial symbiont within spe-cialized midgut portions called crypts or ceca (19, 24, 25, 28–30,32–37, 51–53, 60). When experimentally deprived of the gut sym-biont, these stinkbugs suffer retarded growth, sterility, and/orhigh mortality (1, 19, 24, 27, 33, 35, 37, 51, 60), indicating impor-tant biological roles of the symbiont for the host. In contrast,facultative secondary symbionts of stinkbugs have been poorlyinvestigated despite their potential ecological relevance: except foran extensive survey of Wolbachia among diverse stinkbugs (31),only sporadic detections of Wolbachia and Sodalis have been re-ported (28, 29, 43).
Seed bugs of the genus Nysius, belonging to the family Lygaei-dae, comprise an exceptional stinkbug group in association withan endocellular primary symbiont. They possess a pair of bacteri-omes in the abdomen, wherein a gammaproteobacterial symbiontSchneideria nysicola is harbored (43). Although biological roles of
Schneideria have not been examined experimentally, nutritionalsupplementation for the host insects is suspected. Meanwhile,their facultative bacterial associates have been poorly character-ized, except for sporadic detection of 16S rRNA gene sequences ofWolbachia and other bacteria (43). Here we report a novel clade ofalphaproteobacterial secondary symbionts associated with Nysiusseed bugs. We demonstrate that this symbiont is not restricted toNysius spp. but widely found in other stinkbugs and also in somefleas and ticks, identifying a novel arthropod-associated symbiontgroup belonging to the order Rickettsiales.
MATERIALS AND METHODSMaterials. Table S1 in the supplemental material lists the stinkbug sam-ples examined in this study. Some of the samples were used in previousstudies (31, 43). Some samples were freshly dissected and subjected toDNA extraction, while other samples were immediately put in glass vialsfilled with acetone and preserved at room temperature until use (18).Several strains of Nysius plebeius and Kleidocerys resedae were maintainedin the laboratory at 25°C under a long-day regime (16 h of light and 8 h ofdark) on sunflower seeds, whole wheat, and distilled water supplementedwith 0.05% ascorbic acid, and these insects were subjected to histologicalanalyses.
Dissection and DNA extraction. The insects were dissected in phos-phate-buffered saline (PBS) (137 mM NaCl, 8.1 mM Na2HPO4, 2.7 mM
Received 2 March 2012 Accepted 5 April 2012
Published ahead of print 13 April 2012
Address correspondence to Yu Matsuura, [email protected].
Supplemental material for this article may be found at http://aem.asm.org/.
Copyright © 2012, American Society for Microbiology. All Rights Reserved.
doi:10.1128/AEM.00673-12
June 2012 Volume 78 Number 12 Applied and Environmental Microbiology p. 4149–4156 aem.asm.org 4149
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 03
Dec
embe
r 20
21 b
y 11
2.19
7.12
0.80
.

KCl, 1.5 mM KH2PO4 [pH 7.4]) with fine forceps under a dissectionmicroscope; the symbiotic organs (either the bacteriome or the midgutfourth section with crypts), the reproductive organs, and the alimentarytracts (excluding the symbiotic section if present) of the insects were iso-lated for DNA extraction. For small insects whose body length is less than1 cm, the whole body or the whole abdomen was used instead. Each of thetissue samples was homogenized in a 1.5-ml plastic tube and digested withlysis buffer (10 mM Tris-HCl [pH 8.0], 1 mM EDTA, 0.1 M NaCl, 0.5%sodium dodecyl sulfate, 0.2 mg/ml proteinase K) at 56°C overnight. Thelysate was extracted with phenol-chloroform and subjected to ethanolprecipitation, and all the precipitated nucleic acid was dried and dissolvedin TE buffer (10 mM Tris-HCl [pH 8.0], 1 mM EDTA).
PCR, cloning, and DNA sequencing. A 1.5-kb fragment of bacterial16S rRNA gene was amplified with primers 16SA1 and 16SB1 (see TableS2 in the supplemental material). The PCR was conducted with Ampli-TaqGold DNA polymerase (Applied Biosystems) and its supplementedbuffer system under the following temperature profile: (i) 95°C for 10min; (ii) 35 cycles, with 1 cycle consisting of 95°C for 30 s, 52°C for 30 s,and 72°C for 2 min; and (iii) a final extension step at 72°C for 10 min. ThePCR products were subjected to cloning, restriction fragment length poly-morphism (RFLP) genotyping, and DNA sequencing with BigDye termi-nator v3.1 cycle sequencing kit and 3130/3130xl genetic analyzer (AppliedBiosystems) as described previously (35). A 1.2-kb fragment of the 16SrRNA gene of the alphaproteobacterial symbiont was amplified with theprimers 16SA1 and Alp1203R (Table S2), and the PCR products werecleaned using exonuclease I (New England BioLabs) and alkaline phos-phatase (shrimp) (TaKaRa) and subjected to direct cycle sequencing. Ar-tifactual chimeric sequences were detected by Mallard software (3).
Diagnostic PCR. A 0.25-kb region of the 16S rRNA gene of the alpha-proteobacterial symbiont was amplified with the primers Alp976F (Alpstands for alphaproteobacterial, and F stands for forward) and Alp1203R(R stands for reverse) (see Table S2 in the supplemental material). A0.6-kb fragment of the wsp gene of the Wolbachia symbiont was amplifiedwith the primers wspF and wspR (Table S2). The PCRs were performedunder the following temperature profile: 95°C for 10 min, followed by 35cycles, with 1 cycle consisting of 95°C for 30 s, 55°C for 45 s, and 72°C for1 min. Negative- and positive-control samples were included. To confirmthe quality of the DNA samples, a 0.65-kb fragment of insect 18S rRNAgene was amplified with primers 2880 and B (Table S2) under the follow-ing temperature profile: 95°C for 10 min, followed by 30 cycles, with 1cycle consisting of 95°C for 15 s, 55°C for 15 s, and 72°C for 1 min.
Molecular phylogenetic analysis. Multiple alignments of nucleotidesequences were generated by the program MUSCLE (10) within a soft-ware package of MEGA5.05 (61). Ambiguously aligned nucleotide siteswere corrected manually or removed. Phylogenetic analyses were con-ducted by three methods: maximum likelihood (ML), maximum parsi-mony (MP), and neighbor joining (NJ). ML trees were created by thePhyML 3.0 program (20), while MP and NJ trees were constructed byMEGA 5.0.5. In the ML analyses, the GTR � I � G (general time revers-ible with invariable sites allowed and the rate variation among sites)model (for Fig. 1 and for Fig. S2 in the supplemental material) and theHKY (Hasegawa-Kishino-Yano) � I � G model (for Fig. S1) were chosenfor the 16S rRNA gene data set on the basis of the Akaike informationcriterion using the program Modeltest 3.7 (50) in conjunction with theprogram PAUP* (59). Bootstrap tests were conducted with 100 replica-tions in the ML analyses and with 1,000 replications in the MP and NJanalyses.
Fluorescent in situ hybridization. Table S2 in the supplemental ma-terial lists the fluorochrome-labeled oligonucleotide probes used for 16SrRNA-targeted fluorescent in situ hybridization. The insects were dis-sected, thoroughly washed in PBS, and fixed in Carnoy’s solution (etha-nol-chloroform-acetic acid [6:3:1]). After the tissues were fixed overnight,they were treated with 6% hydrogen peroxide in 80% ethanol for severalweeks for quenching autofluorescence of the tissues (38). After thoroughwashing with absolute ethanol and phosphate-buffered saline with Tween
20 (PBST) buffer (137 mM NaCl, 8.1 mM Na2HPO4, 2.7 mM KCl, 1.5mM KH2PO4 [pH 7.4], 0.2% Tween 20), the samples were incubated withhybridization buffer (20 mM Tris-HCl [pH 8.0], 0.9 M NaCl, 0.01% so-dium dodecyl sulfate, 30% formamide) three times for 5 min each time.Then, the samples were hybridized with hybridization buffer containingthe probes (100 nM each) and SYTOX green (0.25 �M) overnight. Afterthorough washing with PBST, the samples were mounted with Slowfadeantifade solution (Invitrogen) and observed with a laser confocal micro-scope (Pascal 5; Carl Zeiss).
Electron microscopy. Insects were dissected with fine forceps in 0.1 Msodium phosphate buffer (pH 7.4) containing 2.5% glutaraldehyde, andisolated bacteriomes and ovaries were prefixed in the fixative at 4°C over-night and postfixed in 2% osmium tetroxide at 4°C for 60 min. Afterdehydration through an ethanol series, the materials were embedded inSpurr resin. Ultrathin sections were made on an ultramicrotome (Ultra-cat-N; Leichert-Nissei), mounted on copper meshes, stained with uranylacetate and lead citrate, and observed with a transmission electron micro-scope (model H-7600; Hitachi).
Nucleotide sequence accession numbers. The nucleotide sequencesdetermined in this study have been deposited in the DDBJ/EMBL/GenBank nucleotide sequence database under the accession numbersJQ726711 to JQ726831 (see Tables S1 and S3 and Fig. S1 and S2 in thesupplemental material).
RESULTSBacterial 16S rRNA gene sequences from Nysius plebeius. Thebacteriomes, ovaries, and midguts were dissected from five adultfemales of N. plebeius collected in Tsukuba, Japan, and subjectedto DNA extraction. The DNA samples were subjected to PCR,cloning, and RFLP genotyping of a 1.5-kb region of the bacterial16S rRNA gene. A total of 173 clones were analyzed, and almost allof them, except for three clones with unique RFLP patterns, wereclassified into three RFLP genotypes, tentatively named type A (98clones), type B (43 clones), and type C (29 clones) (see Table S3 inthe supplemental material). The type A clones, 1,479 bp in size,were predominant in the bacteriome (48/55 clones) and the ovary(38/59 clones) but minor in the midgut (12/57 clones) and repre-sented Schneideria nysicola, the bacteriome-associated gamma-proteobacterial primary symbiont of Nysius spp. (43). The type Bclones, 1,427 bp in size, were obtained from all the tissues at lowerfrequencies (4/55 clones from the bacteriome, 12/59 clones fromthe ovary, and 27/57 clones from the midgut), which representedan alphaproteobacterial Wolbachia symbiont of N. plebeius (43).The type C clones, 1,420 bp in size, were identified from all thetissues at the lowest frequencies (3/55 clones from the bacteriome,9/59 clones from the ovary, and 17/57 clones from the midgut),with the highest BLAST hit to an alphaproteobacterial 16S rRNAgene sequence belonging to the Rickettsiales from the tick Ixodesovatus (99.1% [1,147/1,157] identity; GenBank accession numberAB297807) (17). Hereafter, we refer to the type C bacterium as thealpha-symbiont of N. plebeius. Of three exceptional clones, twoturned out to be chimeras of the different symbiont sequences,whereas a clone obtained from the midgut exhibited the highestBLAST hit to a 16S rRNA gene sequence of Phyllobacterium myr-sinacearum (99.9% [1,335/1,337] identity; GenBank accessionnumber AB681132).
Ubiquitous occurrences of the alpha-symbiont and Wolba-chia symbiont in Nysius spp. and Kleidocerys resedae. Similarly,we analyzed the bacterial 16S rRNA sequences obtained fromeight additional populations of N. plebeius, four populations ofNysius sp. 1, a population of Nysius sp. 2, and a population of N.expressus. In addition to the primary symbiont Schneideria, the
Matsuura et al.
4150 aem.asm.org Applied and Environmental Microbiology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 03
Dec
embe
r 20
21 b
y 11
2.19
7.12
0.80
.

alpha-symbiont was consistently detected from all the species andpopulations. The 16S rRNA gene sequences of the alpha-symbi-ont, 1,421 bp in size, were completely or nearly identical (99.9 to100% identities) to the sequence from N. plebeius. The Wolbachiasymbiont was also detected from all the species and populationsbut the population of N. plebeius from Iriomote-jima, Japan.From two populations of Nysius sp. 1, a population of Nysius sp. 2,and a population of N. expressus, a different type of alphaproteo-bacterial 16S rRNA gene sequences was also detected, which ex-hibited the highest BLAST hit to the sequence of Rickettsia belliifrom the tick Dermacentor variabilis (99.6% [1,418 or 1,419/1,424] identity; GenBank accession number CP000087). Further-more, a number of minor bacterial 16S rRNA gene sequences wereidentified from the midgut and whole-body DNA samples, mostof which are probably derived from gut microbes (see Table S3 inthe supplemental material).
We also analyzed bacterial 16S rRNA sequences from Klei-docerys resedae, a lygaeid species related to but distinct fromNysius spp., which possesses a bacteriome-associated primarysymbiont Kleidoceria schneideri (39). Of two insect populationsexamined, the alpha-symbiont and a Rickettsia symbiont weredetected at low frequencies in the insect population fromOkunikko, Japan, whereas Wolbachia symbionts were found inboth populations (see Table S3 in the supplemental material).The 16S rRNA gene sequences of the alpha-symbiont, 1,421 bpin size, were almost identical (99.4 to 99.5% identities) to thesequences from Nysius spp.
Next we performed a wider diagnostic PCR survey of the al-pha-symbiont and the Wolbachia symbiont across many popula-tions of Nysius spp. and K. resedae. A total of 183 individuals from13 populations of Nysius plebeius, 106 individuals from seven pop-ulations of Nysius sp. 1, 21 individuals from four populations ofNysius sp. 2, 33 individuals from a population of N. expressus, and46 individuals from two populations of K. resedae were analyzed.Both the alpha-symbiont and the Wolbachia symbiont were de-tected from almost all the insect species and populations at highfrequencies. The overall infection frequencies were 77.6% (142/183) in N. plebeius, 87.7% (93/106) in Nysius sp. 1, 81.0% (17/21)in Nysius sp. 2, 100% (33/33) in N. expressus, and 87.0% (40/46) inK. resedae for the alpha-symbiont and 86.9% (159/183) in N. ple-beius, 88.7% (94/106) in Nysius sp. 1, 95.2% (20/21) in Nysius sp.2, 100% (33/33) in N. expressus, and 100% (46/46) in K. resedae forthe Wolbachia symbiont (see Table S1 in the supplemental mate-rial).
Occurrences of the alpha-symbiont in diverse stinkbugs.Furthermore, we performed a diagnostic PCR survey of the alpha-symbiont across diverse heteroptaran bugs representing 24 fami-lies, 190 species, and 536 individuals. Among these bugs, thealpha-symbiont was detected from 11 species representing sixfamilies, including Arocatus melanostomus (2/2) (Lygaeidae), Di-morphopterus pallipes (1/1) (Blissidae), Horridipamera incon-spicua (1/2) and Paromius exguus (3/3) (Rhyparochromidae),Physopelta gutta (2/2) and Physopelta cincticollis (1/3) (Largidae),Neuroctenus castaneus (1/1) (Aradidae), and Dolycoris baccarum(1/15), Eysarcoris guttiger (1/8), Eysarcoris annamita (12/12), andPiezodorus hybneri (1/7) (Pentatomidae) (see Table S1 in the sup-plemental material).
For each of the alpha-symbiont-positive species, a 1.2-kb re-gion of the 16S rRNA gene was sequenced. All the sequences, 1,129
to 1,150 bp in size, exhibited very high sequence identities (99.6 to100%) to the alpha-symbiont sequences from Nysius spp.
Phylogenetic placement of alpha-symbionts from diversestinkbugs. Molecular phylogenetic analyses based on the 16SrRNA gene sequences revealed that the alpha-symbionts of Nysiusspp., K. resedae, and other stinkbugs form a very compact mono-phyletic group, wherein only a few nucleotide variations are pres-ent among them. Notably, the alpha-symbiont clade also con-tained 16S rRNA gene sequences of uncultured bacteria derivedfrom fleas (13) and ticks (17, 44). In the Alphaproteobacteria, thealpha-symbiont clade was placed within the Rickettsiales, whosemembers are endocellular symbionts/parasites/pathogens such asRickettsia, Wolbachia, Ehrlichia, Anaplasma, etc. (6, 9). The alpha-symbiont clade was allied to the following bacterial groups: theendosymbionts of acanthamoebae (15), the endosymbionts ofcnidarians (14, 58), and Midichloria spp., the mitochondrion-associated endosymbionts of ticks (12, 54) (Fig. 1).
Phylogenetic placement of Wolbachia symbionts and Rick-ettsia symbionts of Nysius spp. and Kleidecerys resedae. Most ofthe 16S rRNA gene sequences of the Wolbachia symbionts fromNysius spp. and K. resedae were placed in the Wolbachia super-group B, except for a sequence from K. resedae in the Wolbachiasupergroup A (Fig. 1; see also Fig. S1 in the supplemental mate-rial). In the supergroup B, the Wolbachia sequences from differentpopulations of the same Nysius species were almost identical toeach other, except for the two distinct lineages of the Wolbachiasequences from Nysius sp. 1. The 16S rRNA gene sequences of theRickettsia symbionts from N. expressus, Nysius sp. 1, and Nysius sp.2 were all allied to the sequence of Rickettsia bellii, while the Rick-ettsia sequence from K. resedae clustered with Rickettsia felis and R.akari (Fig. 1; see also Fig. S2 in the supplemental material).
Localization and fine structure of the alpha-symbiont inNysius plebeius and Kleidocerys resedae. The following labora-tory-maintained strains of N. plebeius were subjected to fluores-cent in situ hybridization of the symbionts. The NpFky002 (Npstands for N. plebeius, and Fky stands for Fukuyama) strain wasestablished from an adult female collected at Fukuyama, Hiro-shima, Japan, and infected with the alpha-symbiont only in addi-tion to the Schneideria symbiont. The NpTsk002 (Tsk stands forTsukuba) strain was derived from an adult female collected inTsukuba, Ibaraki, Japan, and infected with both the alpha-symbi-ont and the Wolbachia symbiont in addition to the Scineideriasymbiont. In the disymbiotic strain NpFky002, the alpha-symbi-ont signals were detected in various tissues, including bacteri-omes, ovaries, and midguts at low densities (Fig. 2A to D). In thebacteriomes, the alpha-symbiont signals were sporadically foundin the cytoplasm of bacteriocytes together with intense signals ofthe Schneideria symbiont (Fig. 2A). In the ovaries, the alpha-sym-biont signals were sparsely detected in nurse cells, follicular cells,and Schneideria-harboring ovarial bacteriocytes (Fig. 2B). At theanterior pole of oocytes, the alpha-symbiont signals were seen asfilamentous bacterial cells smaller than the Schneideria symbiontcells, where the alpha-symbiont cells formed a cluster adjacent tobut distinct from a larger cluster of tubular cells of the Schneideriasymbiont (Fig. 2C). Midgut epithelial cells also exhibited sparsesignals of the alpha-symbiont (Fig. 2D). In the trisymbiotic strainNpTsk002, the alpha-symbiont signals generally exhibited local-ization patterns similar to those in strain NpFky002, together withsignals of the Wolbachia symbiont. The alpha-symbiont signalscooccurred in bacteriocytes, nurse cells, follicular cells, and ovarial
Novel Facultative Symbiont of Stinkbugs
June 2012 Volume 78 Number 12 aem.asm.org 4151
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 03
Dec
embe
r 20
21 b
y 11
2.19
7.12
0.80
.

FIG 1 Phylogenetic placement of the alpha-symbionts, the Wolbachia symbionts, and the Rickettsia symbionts from Nysius spp., Kleidocerys resedae, and otherstinkbugs on the basis of 16S rRNA gene sequences. A maximum likelihood phylogeny inferred from 1,118 aligned nucleotide sites is shown, while maximumparsimony and neighbor-joining analyses gave substantially the same results. Bootstrap support values of 50% or higher are shown at the nodes in the followingorder: maximum likelihood/maximum parsimony/neighbor joining. Asterisks indicate support values lower than 50%. Sequences identified from stinkbugs areindicated in boldface type, while the collection localities in Japan are indicated in parentheses. Sequence accession numbers are shown in brackets. str., strain. Thescale bar indicates the branch length measured by the number of substitutions per site. Accession numbers starting with AB were from DDBJ, those beginningwith FM and FN were from EMBL, those beginning with JQ, FJ, AF, GU, EF, CP, GQ, AE, DQ, AY, and U were from Genbank, and those beginning with NC werefrom NCBI RefSeq.
Matsuura et al.
4152 aem.asm.org Applied and Environmental Microbiology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 03
Dec
embe
r 20
21 b
y 11
2.19
7.12
0.80
.
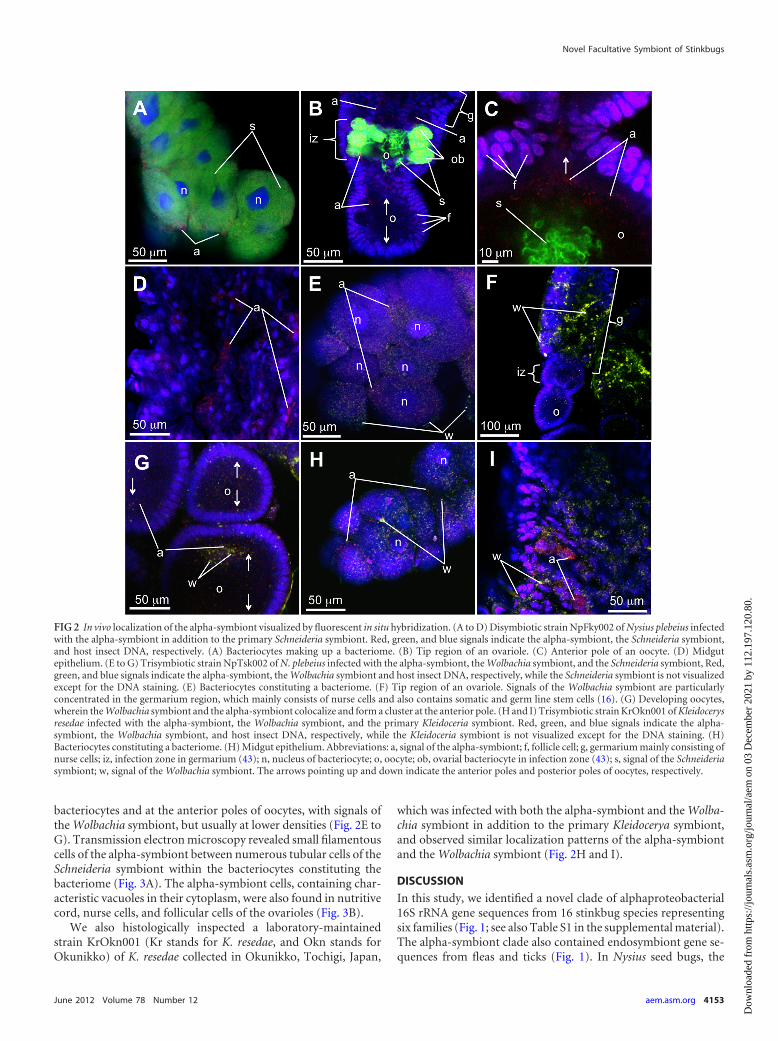
bacteriocytes and at the anterior poles of oocytes, with signals ofthe Wolbachia symbiont, but usually at lower densities (Fig. 2E toG). Transmission electron microscopy revealed small filamentouscells of the alpha-symbiont between numerous tubular cells of theSchneideria symbiont within the bacteriocytes constituting thebacteriome (Fig. 3A). The alpha-symbiont cells, containing char-acteristic vacuoles in their cytoplasm, were also found in nutritivecord, nurse cells, and follicular cells of the ovarioles (Fig. 3B).
We also histologically inspected a laboratory-maintainedstrain KrOkn001 (Kr stands for K. resedae, and Okn stands forOkunikko) of K. resedae collected in Okunikko, Tochigi, Japan,
which was infected with both the alpha-symbiont and the Wolba-chia symbiont in addition to the primary Kleidocerya symbiont,and observed similar localization patterns of the alpha-symbiontand the Wolbachia symbiont (Fig. 2H and I).
DISCUSSION
In this study, we identified a novel clade of alphaproteobacterial16S rRNA gene sequences from 16 stinkbug species representingsix families (Fig. 1; see also Table S1 in the supplemental material).The alpha-symbiont clade also contained endosymbiont gene se-quences from fleas and ticks (Fig. 1). In Nysius seed bugs, the
FIG 2 In vivo localization of the alpha-symbiont visualized by fluorescent in situ hybridization. (A to D) Disymbiotic strain NpFky002 of Nysius plebeius infectedwith the alpha-symbiont in addition to the primary Schneideria symbiont. Red, green, and blue signals indicate the alpha-symbiont, the Schneideria symbiont,and host insect DNA, respectively. (A) Bacteriocytes making up a bacteriome. (B) Tip region of an ovariole. (C) Anterior pole of an oocyte. (D) Midgutepithelium. (E to G) Trisymbiotic strain NpTsk002 of N. plebeius infected with the alpha-symbiont, the Wolbachia symbiont, and the Schneideria symbiont, Red,green, and blue signals indicate the alpha-symbiont, the Wolbachia symbiont and host insect DNA, respectively, while the Schneideria symbiont is not visualizedexcept for the DNA staining. (E) Bacteriocytes constituting a bacteriome. (F) Tip region of an ovariole. Signals of the Wolbachia symbiont are particularlyconcentrated in the germarium region, which mainly consists of nurse cells and also contains somatic and germ line stem cells (16). (G) Developing oocytes,wherein the Wolbachia symbiont and the alpha-symbiont colocalize and form a cluster at the anterior pole. (H and I) Trisymbiotic strain KrOkn001 of Kleidocerysresedae infected with the alpha-symbiont, the Wolbachia symbiont, and the primary Kleidoceria symbiont. Red, green, and blue signals indicate the alpha-symbiont, the Wolbachia symbiont, and host insect DNA, respectively, while the Kleidoceria symbiont is not visualized except for the DNA staining. (H)Bacteriocytes constituting a bacteriome. (H) Midgut epithelium. Abbreviations: a, signal of the alpha-symbiont; f, follicle cell; g, germarium mainly consisting ofnurse cells; iz, infection zone in germarium (43); n, nucleus of bacteriocyte; o, oocyte; ob, ovarial bacteriocyte in infection zone (43); s, signal of the Schneideriasymbiont; w, signal of the Wolbachia symbiont. The arrows pointing up and down indicate the anterior poles and posterior poles of oocytes, respectively.
Novel Facultative Symbiont of Stinkbugs
June 2012 Volume 78 Number 12 aem.asm.org 4153
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 03
Dec
embe
r 20
21 b
y 11
2.19
7.12
0.80
.

alpha-symbionts were at high infection frequencies ranging from76% to 100% within the species (Table S1) and observed as fila-mentous bacteria in various cells and tissues (Fig. 2 and 3). Thealpha-symbiont cells were particularly concentrated at the ante-rior poles of oocytes in the ovaries of adult females, indicating thevertical transmission route of the symbiont through the host gen-erations (Fig. 2 and 3). On the basis of these results, we concludethat the alpha-symbionts comprise a facultative symbiont cladebelonging to the Rickettsiales, whose hosts are stinkbugs and otherarthropods.
The alphaproteobacterial order Rickettsiales contains well-known bacterial groups, such as Wolbachia, Rickettsia, Ehrlichia,and Anaplasma, whose members are associated with a diverse setof host/vector species with various relationships ranging fromparasitic/pathogenic through commensal to mutualistic (6, 9).The alpha-symbiont clade was allied not only to an endosymbiontclade associated with ticks (Midichloria spp.) but also to endosym-biont clades associated with cnidarians, acanthamoebae, and cili-ates (Fig. 1), highlighting the diversity of host-symbiont associa-tions in the Rickettsiales.
Currently, infections with the alpha-symbiont have been
detected only from stinkbugs (Hemiptera), fleas (Sipho-naptera), and ticks (Acari). Biological or ecological connec-tions between the disparate host taxa are obscure, but theirnutritionally unbalanced food sources, such as plant sap defi-cient in essential amino acids for stinkbugs and vertebrateblood lacking in B vitamins for fleas and ticks (8), might be ofsome relevance. Meanwhile, it seems likely that further surveysacross diverse insects and other arthropods would reveal amuch wider host range of the alpha-symbiont.
The biological effects of the alpha-symbiont infection on thehost stinkbugs and other arthropods deserve future study. Con-sidering the high infection frequencies in Nysius spp. and K. rese-dae (see Table S1 in the supplemental material), it seems plausiblethat the alpha-symbiont is conditionally beneficial for the hosts, asfacultative symbionts Serratia, Hamiltonella, and Regiella are fortheir host aphids (45, 48, 49, 56, 63). Alternatively, the possibilitythat the alpha-symbiont may induce cytoplasmic incompatibilityor other reproductive phenotypes in the hosts, thereby spreadingin the host populations, as known for facultative symbiontsWolbachia and Cardinium (11, 66), should be taken into account.Since no remarkable bias in the sex ratio is observed in naturalpopulations of Nysius spp. and K. resedae (Y. Matsuura, personalobservation), it seems unlikely that the alpha-symbiont is causingeither parthenogenesis, feminization, or the male killing. Com-parative genomic analyses of the alpha-symbiont with other sym-biotic bacteria in the Alphaproteobacteria will shed further light onits functional and evolutionary aspects (55).
Among facultative microbial associates of insects and otherarthropods, Wolbachia is the most prevalent one. Many strainsinduce cytoplasmic incompatibility or other reproductive pheno-types, thereby driving their own infections into the host popula-tions (11, 66). Some strains conditionally benefit the fitness oftheir hosts (4, 5, 21, 62, 64). Some strains are indispensable fortheir hosts (7, 23, 26, 42). Wolbachia infections are estimated to beprevalent in more than 60% of insect species (22). Similarly, Rick-ettsia (65), Spiroplasma (2), Arsenophonus (47), Cardinium (67),and Sodalis (57) have been recognized as comprising ecologicallywidespread and phylogenetically coherent facultative symbiontgroups. Although our current data are still limited, we expect thatthe alpha-symbiont clade identified in this study would constitutea new group of such widespread symbionts associated with insectsand other arthropods.
On account of the distinct and coherent phylogenetic and mi-crobiological traits described in this study, we propose the desig-nation “Candidatus Lariskella arthropodarum” for the novel Rick-ettsiales clade associated with stinkbugs, fleas, and ticks. Thegeneric name is after a Russian animation character Lariska, along, black, and rat-like creature in the classic movie series Che-burashka, reflecting the first molecular identification of the bacte-rium from Russia (44) and the morphology of the bacteriumreminiscent of the character. The specific epithet refers to its as-sociation with arthropod hosts.
The order Rickettsiales is known for a number of animal andhuman pathogens, including Rickettsia prowazekii for epidemictyphus, Rickettsia typhi for murine typhus, Rickettsia rickettsiifor Rocky Mountain spotted fever, Rickettsia felis for flea-bornespotted fever, Orientia tsutsugamushi for scrub typhus, Ehrlichiachaffeensis for monocytic ehrlichiosis, Ehrlichia ruminantium forcattle heartwater, etc. (6). Notably, Mediannikov et al. (44) detecteda 16S rRNA gene sequence of “Candidatus Lariskella arthro-
FIG 3 Transmission electron microscopy of the alpha-symbiont in Nysiusplebeius. (A) In the bacteriocyte, filamentous alpha-symbiont cells are seenbetween numerous Schneideria cells of larger size. (B) An enlarged image ofalpha-symbiont cells in the ovariole. Abbreviations: a, alpha-symbiont cell; s,Schneideria symbiont cell.
Matsuura et al.
4154 aem.asm.org Applied and Environmental Microbiology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 03
Dec
embe
r 20
21 b
y 11
2.19
7.12
0.80
.

podarum” from blood and biopsy samples of patients with acutefevers who had been bitten by Ixodes ticks. Hence, potential med-ical relevance of the bacterial group should be a focal issue infuture studies.
ACKNOWLEDGMENTS
We thank T. Hosokawa, Y. Ayabe, Y. G. Baba, M. Baba, E. Hara, H.Higuchi, C. Himuro, H. Hirayama, M. Hironaka, N. Ijichi, K. Inadomi, Y.Ishii, M. Ishizaki, K. Ito, S. Kada, N. Kaiwa, T. Kashima, A. Kikuchi, K.Kouno, F. Kuchiki, S. Kudo, N. Kumano, M. Moriyama, S. Ohno,M. Sakakibara, G. Sakurai, M. Tadenuma, M. Takai, T. Takemoto, M.Tanahashi, K. Tanaka, H. Toju, K. Tsuji, N. Tsurusaki, and T. Yasuda forinsect samples and Y. Kamagata for logistic support.
This study was supported by the Program for Promotion of Basic andApplied Research for Innovations in Bio-oriented Industry (BRAIN).Y.M. was supported by the Japan Society for the Promotion of Science(JSPS) Predoctoral Fellowship for Young Scientists.
REFERENCES1. Abe Y, Mishiro K, Takanashi M. 1995. Symbiont of brown-winged green
bug, Plautia stali Scott. Jpn. J. Appl. Entomol. Zool. 39:109 –115.2. Anbutsu H, Fukatsu T. 2011. Spiroplasma as a model insect endosymbi-
ont. Environ. Microbiol. Rep. 3:144 –153.3. Ashelford KE, Chuzhanova NA, Fry JC, Jones AJ, Weightman AJ. 2006.
New screening software shows that most recent large 16S rRNA gene clonelibraries contain chimeras. Appl. Environ. Microbiol. 72:5734 –5741.
4. Brownlie JC, et al. 2009. Evidence for metabolic provisioning by a commoninvertebrate endosymbiont, Wolbachia pipientis, during periods of nutritionalstress. PLoS Pathog. 5:e1000368. doi:10.1371/journal.ppat.1000368.
5. Casiraghi M, et al. 2002. Tetracycline treatment and sex-ratio distortion:a role for Wolbachia in the moulting of filarial nematodes? Int. J. Parasitol.32:1457–1468.
6. Darby AC, Cho NH, Fuxelius HH, Westberg J, Andersson SG. 2007.Intracellular pathogens go extreme: genome evolution in the Rickettsiales.Trends Genet. 23:511–520.
7. Dedeine F, et al. 2001. Removing symbiotic Wolbachia bacteria specifi-cally inhibits oogenesis in a parasitic wasp. Proc. Natl. Acad. Sci. U. S. A.98:6247– 6252.
8. Douglas AE. 2009. The microbial dimension in insect nutritional ecology.Funct. Ecol. 23:38 – 47.
9. Dumler JS, et al. 2001. Reorganization of genera in the families Rickett-siaceae and Anaplasmataceae in the order Rickettsiales: unification of somespecies of Ehrlichia with Anaplasma, Cowdria with Ehrlichia and Ehrlichiawith Neorickettsia, descriptions of six new species combinations and des-ignation of Ehrlichia equi and ‘HGE agent’ as subjective synonyms ofEhrlichia phagocytophila. Int. J. Syst. Evol. Microbiol. 51:2145–2165.
10. Edgar RC. 2004. MUSCLE: a multiple sequence alignment method withreduced time and space complexity. BMC Bioinformatics 19:113. doi:10.1186/1471-2105-5-113.
11. Engelstädter J, Telschow A. 2009. Cytoplasmic incompatibility and hostpopulation structure. Heredity 103:196 –207.
12. Epis S, et al. 2008. Midichloria mitochondrii is widespread in hard ticks(Ixodidae) and resides in the mitochondria of phylogenetically diversespecies. Parasitology 135:485– 494.
13. Erickson DL, Anderson NE, Cromar LM, Jolley A. 2009. Bacterialcommunities associated with flea vectors of plague. J. Med. Entomol. 46:1532–1536.
14. Fraune S, Bosch TC. 2007. Long-term maintenance of species-specificbacterial microbiota in the basal metazoan Hydra. Proc. Natl. Acad. Sci.U. S. A. 104:13146 –13151.
15. Fritsche TR, et al. 1999. In situ detection of novel bacterial endosymbi-onts of Acanthamoeba spp. phylogenetically related to members of theorder Rickettsiales. Appl. Environ. Microbiol. 65:206 –212.
16. Frydman HM, Li JM, Robson DN, Wieschaus E. 2006. Somatic stem cellniche tropism in Wolbachia. Nature 441:509 –512.
17. Fujita H, et al. 2007. Some suggestive records of rickettsiae isolated fromticks in Korea and central China. Annu. Rep. Ohara Hosp. 47:21–24.
18. Fukatsu T. 1999. Acetone preservation: a practical technique for molec-ular analysis. Mol. Ecol. 8:1935–1945.
19. Fukatsu T, Hosokawa T. 2002. Capsule-transmitted gut symbiotic bac-
terium of the Japanese common plataspid stinkbug, Megacopta punctatis-sima. Appl. Environ. Microbiol. 68:389 –396.
20. Guindon S, et al. 2010. New algorithms and methods to estimate maxi-mum-likelihood phylogenies: assessing the performance of PhyML 3.0.Syst. Biol. 59:307–321.
21. Hedges LM, Brownlie JC, O’Neill SL, Johnson KN. 2008. Wolbachia andvirus protection in insects. Science 322:702.
22. Hilgenboecker K, Hammerstein P, Schlattmann P, Telschow A, WerrenJH. 2008. How many species are infected with Wolbachia? A statisticalanalysis of current data. FEMS Microbiol. Lett. 281:215–220.
23. Hoerauf A, et al. 1999. Tetracycline therapy targets intracellular bacteriain the filarial nematode Litomosoides sigmodontis and results in filarialinfertility. J. Clin. Invest. 103:11–18.
24. Hosokawa T, Kikuchi Y, Nikoh N, Shimada M, Fukatsu T. 2006. Stricthost-symbiont cospeciation and reductive genome evolution in insect gutbacteria. PLoS Biol. 4:e337. doi:10.1371/journal.pbio.0040337.
25. Hosokawa T, et al. 2010. Phylogenetic position and peculiar genetic traitsof a midgut bacterial symbiont of the stinkbug Parastrachia japonensis.Appl. Environ. Microbiol. 76:4130 – 4135.
26. Hosokawa T, Koga R, Kikuchi Y, Meng XY, Fukatsu TT. 2010. Wolba-chia as a bacteriocyte-associated nutritional mutualist. Proc. Natl. Acad.Sci. U. S. A. 107:769 –774.
27. Hosokawa T, et al. 2012. Mothers never miss the moment: a fine-tunedmechanism for vertical symbiont transmission in a subsocial insect. Anim.Behav. 83:293–300.
28. Kaiwa N, et al. 2010. Primary gut symbiont and secondary, Sodalis-alliedsymbiont of the scutellerid stinkbug Cantao ocellatus. Appl. Environ. Mi-crobiol. 76:3486 –3494.
29. Kaiwa N, et al. 2011. Bacterial symbionts of the giant jewel stinkbugEucorysses grandis (Hemiptera: Scutelleridae). Zool. Sci. 28:169 –174.
30. Kaltenpoth M, Winter SA, Kleinhammer A. 2009. Localization andtransmission route of Coriobacterium glomerans, the endosymbiont ofpyrrhocorid bugs. FEMS Microbiol. Ecol. 69:373–383.
31. Kikuchi Y, Fukatsu T. 2003. Diversity of Wolbachia endosymbionts inheteropteran bugs. Appl. Environ. Microbiol. 69:6082– 6090.
32. Kikuchi Y, Meng XY, Fukatsu T. 2005. Gut symbiotic bacteria of thegenus Burkholderia in the broad-headed bugs Riptortus clavatus and Lep-tocorisa chinensis (Heteroptera: Alydidae). Appl. Environ. Microbiol. 71:4035– 4043.
33. Kikuchi Y, Hosokawa T, Fukatsu T. 2007. Insect-microbe mutualismwithout vertical transmission: a stinkbug acquires a beneficial gut symbi-ont from the environment every generation. Appl. Environ. Microbiol.73:4308 – 4316.
34. Kikuchi Y, Hosokawa T, Fukatsu T. 2008. Diversity of bacterial symbi-osis in stinkbugs, p 39 – 63. In Dijk TV (ed), Microbial ecology researchtrends. Nova Science Publishers Inc, Hauppauge, NY.
35. Kikuchi Y, et al. 2009. Host-symbiont co-speciation and reductive ge-nome evolution in gut symbiotic bacteria of acanthosomatid stinkbugs.BMC Biol. 7:2. doi:10.1186/1741-7007-7-2.
36. Kikuchi Y, Hosokawa T, Fukatsu T. 2011. An ancient but promiscuoushost-symbiont association between Burkholderia gut symbionts and theirheteropteran hosts. ISME J. 5:446 – 460.
37. Kikuchi Y, Hosokawa T, Nikoh N, Fukatsu T. 2012. Gut symbioticbacteria in the cabbage bugs Eurydema rugosa and Eurydema dominulus(Heteroptera: Pentatomidae). Appl. Entomol. Zool. 47:1– 8.
38. Koga R, Tsuchida T, Fukatsu T. 2009. Quenching autofluorescence ofinsect tissues for in situ detection of endosymbionts. Appl. Entomol. Zool.44:281–291.
39. Küchler S, Dettner MK, Kehl S. 2010. Molecular characterization andlocalization of the obligate endosymbiotic bacterium in the birch catkinbug Kleidocerys resedae (Heteroptera: Lygaeidae, Ischnorhynchinae).FEMS Microbiol. Ecol. 73:408 – 418.
40. Kuechler S, Dettner MK, Kehl S. 2011. Characterization of an obligateintracellular bacterium in the midgut epithelium of the bulrush bug Chi-lacis typhae (Heteroptera, Lygaeidae, Artheneinae). Appl. Environ. Micro-biol. 77:2869 –2876.
41. Kuechler S, Renz MP, Dettner K, Kehl S. 2012. Diversity of symbioticorgans and bacterial endosymbionts among lygaeoid bugs of the familiesBlissidae and Lygaeidae (Hemiptera: Heteroptera: Lygaeoidea). Appl. En-viron. Microbiol. 78:2648 –2659.
42. Langworthy NG, et al. 2000. Macrofilaricidal activity of tetracyclineagainst the filarial nematode Onchocerca ochengi: elimination of Wolba-
Novel Facultative Symbiont of Stinkbugs
June 2012 Volume 78 Number 12 aem.asm.org 4155
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 03
Dec
embe
r 20
21 b
y 11
2.19
7.12
0.80
.

chia precedes worm death and suggests a dependent relationship. Proc. R.Soc. B 267:1063–1069.
43. Matsuura Y, et al. 2012. Evolution of symbiotic organs and endosymbi-onts in lygaeid stinkbugs. ISME J. 6:397– 409.
44. Mediannikov O, et al. 2004. Microorganism “Montezuma” of the orderRickettsiales: the potential causative agent of tick-borne disease in the FarEast of Russia. Zh. Mikrobiol. Epidemiol. Immunobiol. Jan-Feb:7–13. (InRussian with English summary.)
45. Montllor CB, Maxmen A, Purcell AH. 2002. Facultative bacterial endo-symbionts benefit pea aphids Acyrthosiphon pisum under heat stress. Ecol.Entomol. 27:189 –195.
46. Moran NA, McCutcheon JP, Nakabachi A. 2008. Genomics and evolu-tion of heritable bacterial symbionts. Annu. Rev. Genet. 42:165–190.
47. Nováková E, Hypsa V, Moran NA. 2009. Arsenophonus, an emergingclade of intracellular symbionts with a broad host distribution. BMC Mi-crobiol. 9:143. doi:10.1186/1471-2180-9-143.
48. Oliver KM, Russell JA, Moran NA, Hunter MS. 2003. Facultative bac-terial symbionts in aphids confer resistance to parasitic wasps. Proc. Natl.Acad. Sci. U. S. A. 100:1803–1807.
49. Oliver KM, Degnan PH, Burke GR, Moran NA. 2010. Facultative sym-bionts in aphids and the horizontal transfer of ecologically importanttraits. Annu. Rev. Entomol. 55:247–266.
50. Posada D, Crandall KA. 1998. MODELTEST: testing the model of DNAsubstitution. Bioinformatics 14:817– 818.
51. Prado SS, Almeida RP. 2009. Phylogenetic placement of pentatomidstink bug gut symbionts. Curr. Microbiol. 58:64 – 69.
52. Prado SS, Rubinoff D, Rodrigo P, Almeida P. 2006. Vertical transmis-sion of a pentatomid caeca-associated symbiont. Ann. Entomol. Soc. Am.99:577–585.
53. Prado SS, Hung KY, Daugherty MP, Almeida RP. 2010. Indirect effectsof temperature on stink bug fitness, via maintenance of gut-associatedsymbionts. Appl. Environ. Microbiol. 76:1261–1266.
54. Sassera D, et al. 2006. ‘Candidatus Midichloria mitochondri’, an endo-symbiont of the tick Ixodes ricinus with a unique intramitochondrial life-style. Int. J. Syst. Evol. Microbiol. 56:2535–2540.
55. Sassera D, et al. 2011. Phylogenomic evidence for the presence of aflagellum and cbb3 oxidase in the free-living mitochondrial ancestor. Mol.Biol. Evol. 28:3285–3296.
56. Scarborough CL, Ferrari J, Godfray HC. 2005. Aphid protected frompathogen by endosymbiont. Science 310:1781.
57. Snyder AK, McMillen CM, Wallenhorst P, Rio RV. 2011. The phylogenyof Sodalis-like symbionts as reconstructed using surface-encoding loci.FEMS Microbiol. Lett. 317:143–151.
58. Sunagawa S, Woodley CM, Medina M. 2010. Threatened corals provideunderexplored microbial habitats. PLoS One 5:e9554. doi:10.1371/journal.pone.0009554.
59. Swofford DL. 2001. PAUP* version 4.0b10. Sinauer, Sunderland, MA.60. Tada A, et al. 2011. Obligate association with gut bacterial symbiont in
Japanese populations of the southern green stinkbug Nezara viridula(Heteroptera: Pentatomidae). Appl. Entomol. Zool. 46:483– 488.
61. Tamura K, et al. 2011. MEGA5: molecular evolutionary genetics analysisusing maximum likelihood, evolutionary distance, and maximum parsi-mony methods. Mol. Biol. Evol. 28:2731–2739.
62. Teixeira L, Ferreira A, Ashburner M. 2008. The bacterial symbiontWolbachia induces resistance to RNA viral infections in Drosophila mela-nogaster. PLoS Biol. 6(12):e2. doi:10.1371/journal.pbio.1000002.
63. Tsuchida T, Koga R, Fukatsu T. 2004. Host plant specialization governedby facultative symbiont. Science 303:1989.
64. Weeks AR, Turelli M, Harcombe WR, Reynolds KT, Hoffmann AA.2007. From parasite to mutualist: rapid evolution of Wolbachia in naturalpopulations of Drosophila . PLoS Biol. 5 :e114. doi:10.1371/journal.pbio.0050114.
65. Weinert LA, Werren JH, Aebi A, Stone GN, Jiggins FM. 2009. Evolutionand diversity of Rickettsia bacteria. BMC Biol. 7:6. doi:10.1186/1741-7007-7-6.
66. Werren JH, Baldo L, Clark ME. 2008. Wolbachia: master manipulators ofinvertebrate biology. Nat. Rev. Microbiol. 6:741–751.
67. Zchori-Fein E, Perlman SJ. 2004. Distribution of the bacterial symbiontCardinium in arthropods. Mol. Ecol. 13:2009 –2016.
Matsuura et al.
4156 aem.asm.org Applied and Environmental Microbiology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 03
Dec
embe
r 20
21 b
y 11
2.19
7.12
0.80
.


















