
A Mechanochemical Model for Adult Dermal Wound …cbsg/Materials/OlsenSherrattMaini95.pdf · A...
16
J . theor . Biol . (1995) 177, 113–128 0022–5193/95/220113+16 $12.00/0 7 1995 Academic Press Limited A Mechanochemical Model for Adult Dermal Wound Contraction and the Permanence of the Contracted Tissue Displacement Profile L O†, J A. S‡ P K. M† †Centre for Mathematical Biology , Mathematical Institute 24-29 St Giles ’, Oxford , OX13LB, U.K. ‡Nonlinear Systems Laboratory , Mathematics Institute University of Warwick , Coventry , CV47AL, U.K. (Received on 19 July 1994, Accepted in revised form on 7 June 1995) The healing of adult mammalian skin wounds involves a complex sequence of spatially and temporally coordinated processes. Wound contraction, by reducing the size of the injury, is an intrinsic component of full-thickness excisional dermal wound healing. The underlying biomechanics of wound contraction, however, are not fully understood, and little is known about the pathogenesis of severe medical conditions known as fibrocontractive diseases. The aim of this work is to investigate a deterministic mathematical model in order to obtain insight into the mechanistic relationships between wound contraction and associated normal and pathological healing processes. The model describes the essential roles of fibroblast and myofibroblast cells, a chemical growth factor and the extracellular matrix which includes type I collagen. The model results are qualitatively consistent with the biology of fibroplasia and wound contraction. It is shown that a contracted state evolves during a (long) transient phase of healing known as ‘‘proliferation’’, while collagen kinetics are fundamental to the considerably longer ‘‘remodelling’’ phase. Some quantitative results, notably on the evolution of wound contraction, compare favourably with experimental data. Application of the model to adult human dermal wound healing in vivo, with a greater understanding of the underlying biological mechanisms involved, may suggest strategies for controlling contraction and fibrocontractive diseases. 7 1995 Academic Press Limited 1. Introduction The healing of full-thickness skin wounds in adult mammal involves a highly complex and inter- dependent series of repair processes, operating within characteristic spatio-temporal intervals (Clarke, 1988). Wound contraction, the biomechanical phenomenon in which the wound boundaries are drawn inwards, is a ubiquitous and essential feature in the healing of these wounds and, together with tissue formation, normally effects full wound closure (Rudolph, 1980; Rudolph et al ., 1992). The rate and extent of contraction depends upon species, age, wound dimensions and other parameters, such as infection and anatomical location (McGrath & Hundahl, 1982; McGrath & Simon, 1983; Rudolph, 1979; Skalli & Gabbiani, 1988; Skalli et al ., 1989). Fetal wounds, for example, are believed to heal without significant contraction or scarring (Skalli & Gabbiani, 1988; Shah et al ., 1992; Longaker et al ., 1992). In adult humans, a typical excisional wound may undergo a 20–30% reduction in wound area because of contraction over a period of several weeks, while in other mammals the rate and extent of 113
-
Upload
hoangxuyen -
Category
Documents
-
view
218 -
download
0
Transcript of A Mechanochemical Model for Adult Dermal Wound …cbsg/Materials/OlsenSherrattMaini95.pdf · A...

J. theor. Biol. (1995) 177, 113–128
0022–5193/95/220113+16 $12.00/0 7 1995 Academic Press Limited
A Mechanochemical Model for Adult Dermal Wound Contraction
and the Permanence of the Contracted Tissue
Displacement Profile
L O†, J A. S‡ P K. M†
†Centre for Mathematical Biology, Mathematical Institute 24-29 St Giles’, Oxford,OX1 3LB, U.K.
‡Nonlinear Systems Laboratory, Mathematics Institute University of Warwick,Coventry, CV4 7AL, U.K.
(Received on 19 July 1994, Accepted in revised form on 7 June 1995)
The healing of adult mammalian skin wounds involves a complex sequence of spatially and temporallycoordinated processes. Wound contraction, by reducing the size of the injury, is an intrinsic componentof full-thickness excisional dermal wound healing. The underlying biomechanics of wound contraction,however, are not fully understood, and little is known about the pathogenesis of severe medicalconditions known as fibrocontractive diseases. The aim of this work is to investigate a deterministicmathematical model in order to obtain insight into the mechanistic relationships between woundcontraction and associated normal and pathological healing processes. The model describes the essentialroles of fibroblast and myofibroblast cells, a chemical growth factor and the extracellular matrix whichincludes type I collagen. The model results are qualitatively consistent with the biology of fibroplasiaand wound contraction. It is shown that a contracted state evolves during a (long) transient phase ofhealing known as ‘‘proliferation’’, while collagen kinetics are fundamental to the considerably longer‘‘remodelling’’ phase. Some quantitative results, notably on the evolution of wound contraction,compare favourably with experimental data. Application of the model to adult human dermal woundhealing in vivo, with a greater understanding of the underlying biological mechanisms involved, maysuggest strategies for controlling contraction and fibrocontractive diseases.
7 1995 Academic Press Limited
1. Introduction
The healing of full-thickness skin wounds in adultmammal involves a highly complex and inter-dependent series of repair processes, operating withincharacteristic spatio-temporal intervals (Clarke,1988). Wound contraction, the biomechanicalphenomenon in which the wound boundaries aredrawn inwards, is a ubiquitous and essential featurein the healing of these wounds and, together withtissue formation, normally effects full wound closure(Rudolph, 1980; Rudolph et al., 1992). The rate and
extent of contraction depends upon species, age,wound dimensions and other parameters, such asinfection and anatomical location (McGrath &Hundahl, 1982; McGrath & Simon, 1983; Rudolph,1979; Skalli & Gabbiani, 1988; Skalli et al., 1989).Fetal wounds, for example, are believed to healwithout significant contraction or scarring (Skalli &Gabbiani, 1988; Shah et al., 1992; Longaker et al.,1992). In adult humans, a typical excisional woundmay undergo a 20–30% reduction in wound areabecause of contraction over a period of several weeks,while in other mammals the rate and extent of
113

. ET AL .114
contraction is generally greater [up to 80–90% ofwound closure is attributed to contraction in rats;Rudolph (1979) McGrath & Simon (1983)].
By reducing the size of the defect, woundcontraction is usually beneficial to the overall repairprocess. However, insufficient contraction may causedelayed or impaired healing, whilst excessive contrac-tion often induces poor quality repair with substantialscarring (Skalli & Gabbiani, 1988; Jennings & Hunt,1992; Rudolph et al., 1992; Shah et al., 1992). Healingabnormalities occurring at the level of woundcontraction may also result in fibrocontractivediseases, many of which are clinically severe,physically undesirable and functionally debilitating(see, for review, Rudolph, 1980; Skalli & Gabbiani,1988; Rudolph & Vande Berg, 1991; Skalli et al.,1989). Despite intensive research, the underlyingbiological mechanisms of wound contraction and ofassociated pathologies remain incompletely under-stood (Gabbiani, 1992; Rudolph et al., 1992).
The aim of this work, therefore, is to develop amathematical model for the fundamental cell biologyand biomechanics of wound contraction, in order topredict the behaviour of experimental and clinicalwounds and to yield insight into the pathogenesis offibrocontractive diseases.
An effective deterministic model must predictsteady states representing normal skin, a contractedwound and fibrocontractive diseases. The initialinjury is a large perturbation of the normal,unwounded state (Skalli & Gabbiani, 1988), withthe ensuing healing process encapsulated bythe evolution of the model dynamical system. Theprecise sense, however, in which the contractedstate is a ‘‘steady solution’’ of the system, is vitalin elucidating the underlying biological mechanismsof normal and pathological wound contraction,and has attracted recent modelling debate (Murrayet al., 1988; Tranquillo & Murray, 1992). Thus,the emphasis of this paper is on investigating themodel to acquire a greater understanding of thecontracted wound state. A subsequent report willaddress pathological transitions of fibrocontractivedisease states as predicted by the model discussedbelow (L. Olsen et al., submitted).
2. A Biological Overview of Dermal Wound Healing
Adult mammalian skin is composed of a complexarray of local and systemic elements, each providingimportant contributions towards the regulatory andprotective functions of the skin (Odland, 1991; Mast,1992). Anatomically, the skin is partitioned into twolayers (Fig. 1). The outermost layer is the epidermis,
consisting of densely packed keratinocyte cells whichcontinually proliferate and differentiate (Dover &Wright, 1991). The underlying, thicker layer isknown as the dermis and is composed of fibroblastcells, extracellular matrix (ECM), blood capillariesand other circulatory, secretory and structuralcomponents (Odland, 1991; Mast, 1992). Fibroblastsperform many vital roles in dermal wound healingand are central to contraction (Clark, 1988, 1993;Rudolph et al., 1992). The various fibrous and solublemolecules which constitute the ECM include proteinssuch as collagen—the most abundant solid-phasecomponent of the skin, and enzymes which enable theECM to undergo constant turnover (Odland, 1991).Beneath the dermis, but not clearly demarcated fromit, are subcutaneous strata of fatty, muscular andfibrous tissues, interspersed with circulatory vessels(Mast, 1992).
The epidermal response to wounding is welldocumented, both biologically (Clark, 1989; Oldland& Ross, 1968) and mathematically (Sherratt &Murray, 1992; Sherratt et al., 1992). Full-thicknessdermal wound healing, however, is required whenepidermal and dermal tissues are excised (Fig. 1). Themultitude of interregulatory dermal repair processes,of which wound contraction is a prominent andimportant consequence, have attracted enormousbiomedical interest (see, for review, Clark, 1989;Jennings & Hunt, 1992; Mast, 1992; Rudolph et al.,1992). These processes may be grouped into threeoverlapping temporal phases: inflammation, prolifer-ation and remodelling (Clark, 1988).
Platelets from damaged blood vessels initiate theinflammatory phase within a few hours of woundingby aggregating to form a stable clot, thereby sealingthe wound from further damage or infection (Clark,1989). They also release a variety of biochemicalsubstances, including several types of growth factorswhich individually and synergistically regulate manyrepair processes (Sprugel et al., 1987; Clark, 1991;Grotendorst, 1992; Martin et al., 1992). Notably, theyattract inflammatory cells into the wound site,triggering further biochemical and enzymatic cas-cades which initiate the following phases of healing(Clark, 1989).
The proliferative phase usually begins several dayspost-wounding. Epidermal integrity is re-establishedas keratinocytes divide and migrate above the woundtissue (Odland & Ross, 1968). Simultaneously,angiogenesis takes place, whereby new blood capil-laries grow into the wound tissue from surroundingvessels, thus establishing a blood supply and hencemetabolic regulation in the wound (Arnold & West,1992; Whalen & Zetter, 1992). In addition, growth
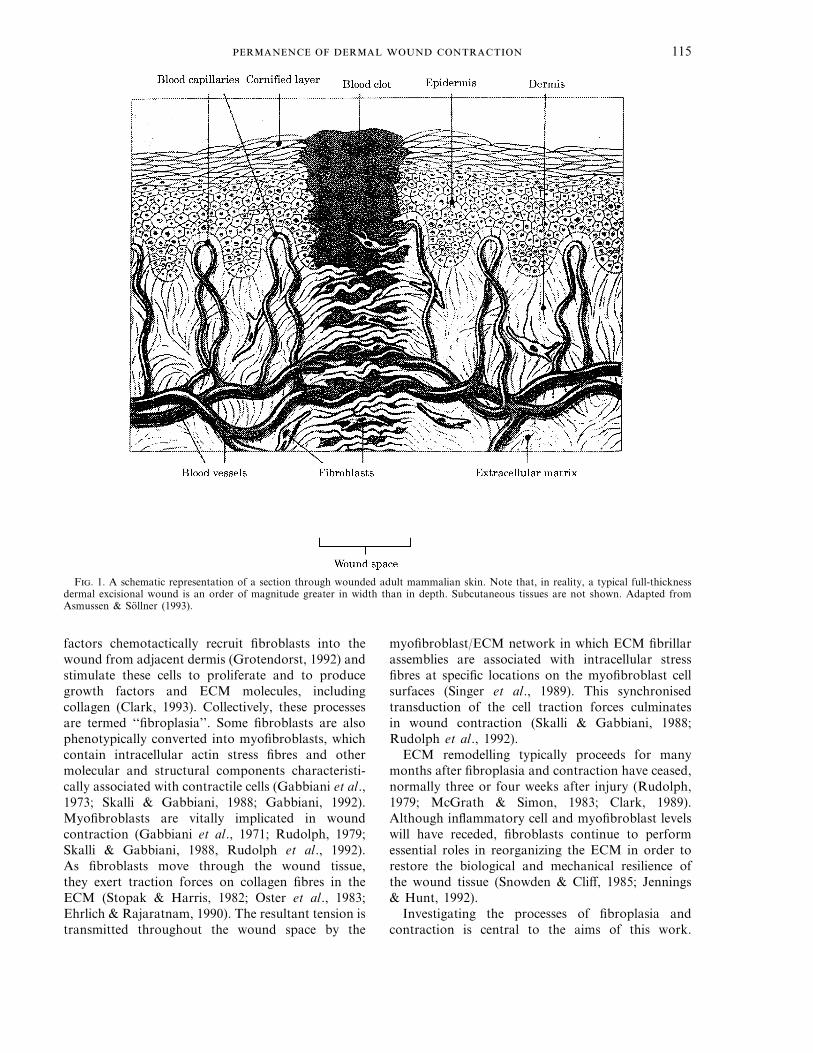
115
F. 1. A schematic representation of a section through wounded adult mammalian skin. Note that, in reality, a typical full-thicknessdermal excisional wound is an order of magnitude greater in width than in depth. Subcutaneous tissues are not shown. Adapted fromAsmussen & Sollner (1993).
factors chemotactically recruit fibroblasts into thewound from adjacent dermis (Grotendorst, 1992) andstimulate these cells to proliferate and to producegrowth factors and ECM molecules, includingcollagen (Clark, 1993). Collectively, these processesare termed ‘‘fibroplasia’’. Some fibroblasts are alsophenotypically converted into myofibroblasts, whichcontain intracellular actin stress fibres and othermolecular and structural components characteristi-cally associated with contractile cells (Gabbiani et al.,1973; Skalli & Gabbiani, 1988; Gabbiani, 1992).Myofibroblasts are vitally implicated in woundcontraction (Gabbiani et al., 1971; Rudolph, 1979;Skalli & Gabbiani, 1988, Rudolph et al., 1992).As fibroblasts move through the wound tissue,they exert traction forces on collagen fibres in theECM (Stopak & Harris, 1982; Oster et al., 1983;Ehrlich & Rajaratnam, 1990). The resultant tension istransmitted throughout the wound space by the
myofibroblast/ECM network in which ECM fibrillarassemblies are associated with intracellular stressfibres at specific locations on the myofibroblast cellsurfaces (Singer et al., 1989). This synchronisedtransduction of the cell traction forces culminatesin wound contraction (Skalli & Gabbiani, 1988;Rudolph et al., 1992).
ECM remodelling typically proceeds for manymonths after fibroplasia and contraction have ceased,normally three or four weeks after injury (Rudolph,1979; McGrath & Simon, 1983; Clark, 1989).Although inflammatory cell and myofibroblast levelswill have receded, fibroblasts continue to performessential roles in reorganizing the ECM in order torestore the biological and mechanical resilience ofthe wound tissue (Snowden & Cliff, 1985; Jennings& Hunt, 1992).
Investigating the processes of fibroplasia andcontraction is central to the aims of this work.

. ET AL .116
Providing a framework in which to model (fromexperimental data) these components of the prolifera-tive phase of healing is therefore the focus of thefollowing section.
3. Modelling Wound Contraction
The model is based upon a fundamental conserva-tion law for each space- and time-dependent quantity,Q=Q(x, t):
1Q1t
=−9 · JQ+fQ ,
where JQ is the flux of Q and fQ are the kinetic terms(rates of production and loss of Q).
The fibroblast density, myofibroblast density,collagen concentration and the concentration of a‘‘generic’’ growth factor are modelled as distinctvariables, thereby accounting for the important cellbiology of fibroplasia and wound contraction(Section 2). Multiple regulatory chemicals are knownto mediate these processes in reality (Clark, 1989,1991), but for simplicity we model a single growthfactor which promotes the healing response. Platelet-derived growth factor (PDGF) exhibits much of therequisite behaviour of the generic chemical (Rosset al., 1986; Huang et al., 1988; Raines et al., 1990;Grotendorst, 1992; Lepisto, 1992), and experimentaldata for PDGF are used to determine the relevantparameter values (see Appendix). The explicitinclusion of fibroblasts and myofibroblasts as distinctcell types is a new modelling approach, enabling thefundamental cellular aspects of wound contraction(Section 2) and of fibrocontractive diseases to be moreaccurately described (Skalli & Gabbiani; 1988; Skalliet al., 1989).
The complex biomechanics of wound contractionhas recently posed a considerable and importantmodelling challenge (McGrath & Simon, 1982;Murray et al., 1988, Tranquillo & Murray, 1992).Following previous mechanochemical models formorphogenesis, we consider the conservation of tis-sue momentum (Murray et al., 1983; Murray, 1989).This yields the ‘‘force-balance’’ equation whichgoverns the tissue displacement—an obvious quanti-tative measure of wound contraction (Murray et al.,1988).
The model variables to represent the fibroblastdensity, n, myofibroblast density, m, chemicalconcentration, c, collagen concentration, r, and tissuedisplacement, u, satisfy the following conservationequations. The fluxes and kinetics model the relevantprocesses involved in fibroplasia and wound contrac-tion (Section 2), with fundamental forms derived
from experimental and theoretical results (seereferences below).
, n(x, t)
1n1t
=9 · $Dn9n−an
(bn+c)2 n9c−n1u
1t%+0rn+
rnmaxcC1/2+c1n01−
nK1
−k1maxcCk+c
n+k2m−dnn. (1)
The contributions to the cell flux are: randomdispersal (Bard & Hay, 1975), modelled by linearFickian diffusion—a standard representation ofrandom cell motion (Murray, 1989); growth factor-mediated chemotaxis (Grotendorst, 1992; Seppa etal., 1992), where the chemotactic sensitivity functionoriginates from a biologically realistic cell surfacereceptor mechanism (Sherratt et al., 1993); andpassive convection by the moving tissue. For thekinetics, there is: mitosis, using a logistic form for thecell proliferation rate, in quantitative agreement withexperimental data on fibroblast growth curves (VandeBerg et al., 1989) with growth factor enhancementof the linear growth rate (Raines et al., 1990);phenotypic transformation into myofibroblasts,stimulated by growth factors (Skalli & Gabbiani,1988; Gabbiani; 1992); a source term from the reversephenotypic transformation; and natural cell death(Alberts et al., 1990).
, m(x, t)
1m1t
=9 · $−m1u
1t%+er0rn+rnmaxc
C1/2+c1m01−mK1
+k1maxcCk+c
n−k2m−dmm. (2)
Without evidence that myofibroblasts are activelymotile, passive convection is taken to be the only flux.The kinetics correspond to those for fibroblasts:namely mitosis, phenotypic transformation and celldeath.
, c(x, t)
1c1t
=9 · $Dc9c−c1u
1t%+(p−1)k'p (n+zpm)cGp+c
−k'c (n+zcm)c
Gc+c−dcc. (3)

117
The chemical diffuses through the ECM (assuminga constant diffusion coefficient) and is convected bythe tissue. The kinetics involve production andconsumption of the chemical by both fibroblasts andmyofibroblasts—the roles of cell surface receptormolecules (Raines et al., 1990; Sato et al., 1993) againsuggest the functional forms for these processes (seeAppendix for details). There is also natural decay ofthe chemical, with a tissue clearance half-life of onlya few minutes (Bowen-Pope et al., 1984; Huang et al.,1988).
, r(x, t)
1r
1t=9 · $−r
1u
1t%+0rr+rrmaxcCr+c1 n+hbm
R2r+r2
−dr (n+hdm)r. (4)
Collagen fibres are generally linked in a mesh-likenetwork (Clark, 1989; Jennings & Hunt, 1992), sopassive convection is the only contribution to thecollagen flux. For the kinetics, there is: synthesis byboth fibroblasts and myofibroblasts, with growthfactor enhancement (Clark, 1989; Pierce et al., 1991);and enzymatic degradation by both cell types (Uitto,1984; Skalli & Gabbiani, 1988; Jeffrey, 1992).
, u(x, t)
0=9 · $m11e
1t+m2
1u
1tI+E'(e+n'uI)
+t0(1+jm)nr
R2t+r2 I%−sru. (5)
Here, e=12(9u+9uT) is the strain tensor and u=9 · u
is the dilation. Following previous mechanochemicalmodels of tissue morphogenesis proposed by Murrayand colleagues (Murray & Oster, 1984; Murray et al.,1988), two contributions of the flux of momentum areincluded. First, there are traction stresses exerted oncollagen fibres in the ECM by fibroblasts at their cellsurfaces. These stresses are modelled as the unittensor multiplied by an isotropic traction coefficient,taken to be proportional to the fibroblast density andto the collagen concentration (Murray et al., 1988).Enhancement of the traction force by myofibroblastsand inhibition at high collagen densities (those typicalof unwounded dermis, for example,) are assumed.Second, there are intrinsic ECM stresses associatedwith tissue deformation. Using the simplifyingassumptions of tissue isotropy, a simple viscoelasticrheology and a linear stress-strain/strain rate depen-
dence (valid for small strains and strain rates), aconstitutive relation for the ECM stress tensor isderived, as in other mechanical models (Landau &Lifshitz, 1970; Fung, 1981; Murray, 1989; Tranquillo& Murray, 1992). The only ‘‘kinetics’’ are due to thebody forces exerted on the tissue by fibrous ECMattachments between the dermal tissue and subcu-taneous layers. These are modelled simply as linearrestoring forces acting against tissue displacementand are taken to be proportional to the displacement(per unit of fibrous ECM) (Murray et al., 1992;Tranquillo & Murray, 1992).
Together with negligible inertial effects owing to thehigh tissue viscosity, these simplifications yield the‘‘force balance’’ equation (5) above. Despite itsapparent complexity, no simpler description of thebiomechanics of wounded or normal dermal tissuehas been formulated. Indeed, the assumptions withrespect to the tissue rheology and to the constitutiverelation have provoked ongoing modelling debate onthe biomechanical detail necessary to derive a realisticforce balance equation.
1-
In reality, wound healing occurs in a three-dimen-sional domain. However, the thickness of the adultskin (approximately 1–2 mm) is much smaller thanthe typical dimensions of the excisional wounds fromwhich experimental data has been collated and towhich this model is applicable (generally between 1and 5 cm). Thus, the skin may be regarded astwo-dimensional (Fung, 1981). In this paper, werestrict attention to the simple cases of (long)rectangular and circular wounds, represented bylinear and axisymmetric geometries as illustrated inFig. 2, so that the problem is one-dimensional.
F. 2. Diagram of the linear and axisymmetric woundgeometries in the model (in the plane of the skin), with x denotingthe distance from the wound centre, x=0. The initial woundboundaries are at (normalized) distance unity from the woundcentre, as shown.

. ET AL .118
To clarify the roles of the parameters in the model,we nondimensionalize the variables and the par-ameters (see Appendix for details) to obtain the fivenondimensional model equations for the 1-D linearwound (omitting the tildes for notational simplicity):
1n1t
=1
1x $Dn1n1x
−a
(b+c)2 n1c1x
−n1u1t%
+s01+Ac
B+c1n(1−gn)
−k1c
C+cn+k2m−dn (6)
1m1t
=1
1x $−m1u1t%+ess01+
AcB+c1m(1−gm)
+k1c
C+cn−k2m−ldm (7)
1c1t
=1
1x $Dc1c1x
−c1u1t%+kc (n+zm)c
gc+c−dcc (8)
1r
1t=
1
1x $−r1u1t%+$v01+
PcQ+c1
×1
f2+r2−drr%(n+hm) (9)
0=m13u
1x2 1t+
12u1x2+n
1
1x $(1+jm)nr
c2+r2 %−sru (10)
Even after non-dimensionalization, the modelcontains 31 parameters. Accurate estimation of thevalues of these parameters is vital before the modelcan be usefully applied, and our methods ofparameters value estimation are discussed in detail inthe Appendix.
Equations (6–10) are solved in the 1-D lineardomain xq0, with symmetric boundary conditions atx=0. The spatial coordinate is rescaled so that theinitial wound space is defined as 0ExE1 (Fig. 2). Thewound is assumed to be sufficiently isolated fromother large perturbations of normal dermis, so that atinfinity, the model variables assume their constant,dermal values (which are calculated in the nextsection).
The initial conditions (t=0) are defined at the onsetof the proliferative phase of healing (Section 2). Thereare no cells inside the wound initially, and the dermallevel of fibroblasts outside, n=1 (see Appendix). Theinitial myofibroblast density is zero everywhere. Theinitial chemical profile, cinit , is due to the supply of
growth factors during the inflammatory phase (Clark,1988; Jennings & Hunt, 1992; Martin et al., 1992),and assumes a form that decreases smoothly to zeroaround the wound margins owing to diffusion.Specifically, we take cinit(x)=1/(1+x4); the modelresults, however, are not sensitive to minor alterationsof the form chosen. The collagen concentration is atsome small value, rinit�1, inside the wound (due tolow-level depositions of ECM molecules by infiltrat-ing cells such as macrophages during the early,inflammatory phase of healing), and the dermal value,r=1 outside. rinit must be positive, however, to avoiddifficulties with the numerical simulations. Finally,the displacement is zero everywhere.
4. Predictions of the Model
4.1.
The general problem of spatially inhomogeneoussteady states is analytically intractable. Motivatedhowever, by the biological requirement that the modelexhibits a locally stable spatially inhomogeneoussteady state which represents unwounded dermis, theuniform case is considered first, as given by the systemof algebraic equations obtained by setting thederivatives in (6–10) to zero.
From (10), either r=0 or u=0. In the case r=0,the kinetic terms of (8) and (9) imply that n, m andc are also zero, yielding a trivial solution. This is oflittle biological interest, since it is not relevant to anyrealistic steady state. If u=0, then there is no tissuedisplacement when the model variables are at auniform steady state, in agreement with the boundarycondition u(0, t)=0.
In addition, normal human dermis produces sig-nificant concentrations of active growth factors onlyin response to external stimuli such as wounding(Clark, 1991; Martin et al., 1992). Therefore,investigations are restricted here to steady states forwhich c=0. The residual, simplified, uniformsteady-state equations are solved explicitly, with thefollowing results (see Table 1). Here ‘‘existence’’means the requirement that the species variables n, m,c and r assume real, nonnegative values. Linearstability is deduced from the standard analysis whichyields specific parameter inequalities, with which theparameter values suggested by our estimates (seeAppendix) are compared.
The scalings chosen to nondimensionalize thefibroblast density, n, and the collagen concentration,r, (n0 and r0 respectively—see Appendix) set theunwounded (normal dermal) steady state values tounity. n+ and m+ may be calculated analytically from

119
(6) and (7). The finding that n+q1 and m+q0 isindicative of healing abnormalities with excessivecellular responses and, in particular, of fibrocontrac-tive diseases in which myofibroblasts persist (Skalli &Gabbiani, 1988; Skalli et al., 1989; Rudolph & VandeBerg, 1991).
4.2.
We assume that the phenomenon of normal woundcontraction is manifested in a stable steady state inwhich the species variables are uniform, namely[n*, m*, c*, r*]=[1, 0, 0, 1], whereas the displace-ment is spatially varying. Under these conditions, (10)implies that the steady displacement profile, u(x),satisfies the simple, linear boundary value problem:
d2udx2−su=0, u(0)=0,
dudx
(a)=0 (11)
The only solution to (11) is u00. Thus, at steadystate, no non-trivial displacement profile exists whenthe species variables are spatially uniform. With acorresponding result for the axisymmetric case [inwhich (11) is a modified Bessel equation], the modeldoes not appear to exhibit a contracted steady state.This problem is discussed further below.
4.3.
The above analyses suggest that the system shouldreturn to the dermal steady state, of which the initialstate is a large perturbation. This is confirmednumerically as shown in Fig. 3, noting that: (a) thefibroblast density, n, gradually reattains its dermalvalue; (b) myofibroblasts, m, appear quickly buttransiently, with highest densities around the woundboundary; (c) the chemical concentration, c, decaysrapidly to zero with diffusion from the wound site;(d) the collagen concentration, r, deviates onlyslightly from its initial profile over the 30-day timecourse of these simulations (passive convection
appears to be the dominant feature here), and isrestored to its dermal value over several years (notshown); (e, f) the displacement, u, becomes negative,most markedly just inside the wound margin. Over aconsiderably longer time scale of several years,however, the numerical simulations confirm that thedisplacement relaxes slowly back to zero (not shown).
4.4.
There is relatively little quantitative experimentaldata on the fundamental processes of fibroplasia andwound contraction in vivo. This is probably aconsequence of the multitude of interacting processesthat occur in real wound healing, which rendermeasurement of the individual variables a difficulttask. Although this is untrue of in vitro models, nosingle experimental system is at present regarded as arealistic simulator of the proliferative phase of woundhealing. Hence, we consider the few sources ofavailable experimental data which can be used toassess the qualitative predictive abilities of our model.
The spatiotemporal evolution of the tissue displace-ment is of particular interest. McGrath & Simon(1983) analysed the rate of change of the area offull-thickness dermal circular excisional wounds inrats (as a fraction of the wound area prior tocontraction). Their experimental data and thenumerical simulations of the axisymmetric model arecompared in Fig. 4; here we use the model withoutcollagen kinetics, since a ‘‘final contracted area’’ canbe calculated for this case (see Fig. 4 legend andSection 4.5). Quantitatively, the two sets of data aresimilar, but not perfectly correlated. Contractionappears to be slightly faster in the experimental databetween 1 and 2 weeks after wounding. As theauthors noted, however, inaccuracies in wound areameasurements are likely during this period becauseepidermal regeneration may hinder the precisecalculation of the positions of the dermal wound
T 1.Analysis of spatially uniform steady states (with c=0) listing the parametric constraints for existence and local
stability of each steady stateSpatially uniform steady states (with c=0)
[n, m, c, r, u]: [0, 0, 0, r, 0] [1, 0, 0, 1, 0] [n+, m+, 0, 1, 0]
Interpretation: ‘‘Trivial’’ ‘‘Normal dermal’’ ‘‘Pathological’’
‘‘Existence’’ conditions: Always exists dQs k2+ldQess
Stability conditions: sQd dQs dQs(deduced from linear essQk2+ld essQk2+ld k2+ldQessstability analysis) kcQgcdc kc (n++zm+)Qgcdc
Stability result: Unstable Stable Unstable(for parameter valuesas in Fig. 3 legend)

. ET AL .120
F. 3. Numerical simulations of the linear wound model, using the method of lines and Gear’s method, showing the evolution of (a)fibroblast cell density, (b) myofibroblast cell density, (c) chemical concentration, (d) collagen concentration, and (e) tissue displacementprofiles, plotted against the distance from the wound centre at sucessive time intervals of Dt=5 (days). In (f ), the simulated movementof tattoo marks located across the wound and adjacent dermis is shown. A finite domain approximation, xa=10, is used for thesesimulations, so zero flux conditions are imposed at this boundary rather than constant, dermal values at infinity. Parameter valuesare: Dn=0.02, a=0.5, b=0.2, s=0.02, A=44.5, B=1, g=0.01, k1=10, C=1, k2=1, d=0.0198, es=0.5, l=10, Dc=1.2, kc=0.4, gc=1,dc=0.5, z=1, v=0.008, P=10, Q=0.1, f=3, dr=0.0008, h=2, m=20, n=0.02, j=10, c=0.005, s=1, rinit=0.01.
margins. The results are very similar over the 30-daytime-course of the simulations in the case wherethe collagen kinetics are included (not shown). Other
experimental data on the area of the contractingwounds (Rudolph, 1979; McGrath & Hundahl, 1982;Rudolph et al., 1992) are also in close agreement
121
with the numerical simulations of the model (notshown).
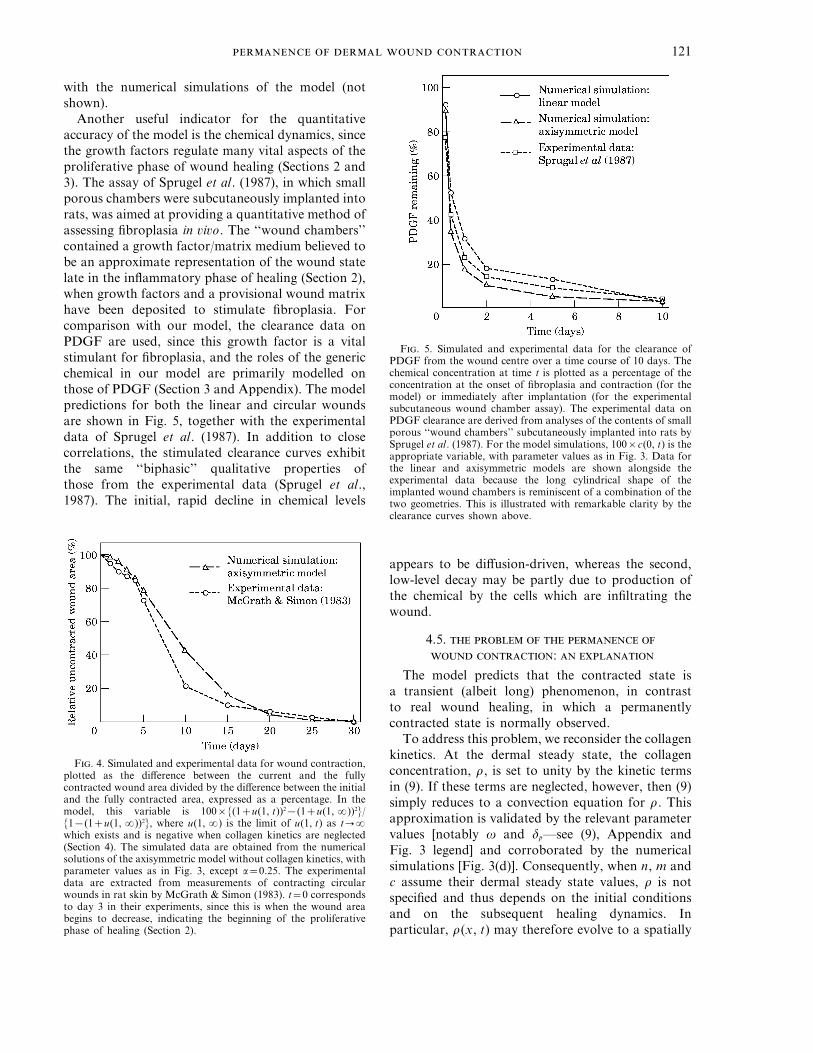
Another useful indicator for the quantitativeaccuracy of the model is the chemical dynamics, sincethe growth factors regulate many vital aspects of theproliferative phase of wound healing (Sections 2 and3). The assay of Sprugel et al. (1987), in which smallporous chambers were subcutaneously implanted intorats, was aimed at providing a quantitative method ofassessing fibroplasia in vivo. The ‘‘wound chambers’’contained a growth factor/matrix medium believed tobe an approximate representation of the wound statelate in the inflammatory phase of healing (Section 2),when growth factors and a provisional wound matrixhave been deposited to stimulate fibroplasia. Forcomparison with our model, the clearance data onPDGF are used, since this growth factor is a vitalstimulant for fibroplasia, and the roles of the genericchemical in our model are primarily modelled onthose of PDGF (Section 3 and Appendix). The modelpredictions for both the linear and circular woundsare shown in Fig. 5, together with the experimentaldata of Sprugel et al. (1987). In addition to closecorrelations, the stimulated clearance curves exhibitthe same ‘‘biphasic’’ qualitative properties ofthose from the experimental data (Sprugel et al.,1987). The initial, rapid decline in chemical levels
F. 5. Simulated and experimental data for the clearance ofPDGF from the wound centre over a time course of 10 days. Thechemical concentration at time t is plotted as a percentage of theconcentration at the onset of fibroplasia and contraction (for themodel) or immediately after implantation (for the experimentalsubcutaneous wound chamber assay). The experimental data onPDGF clearance are derived from analyses of the contents of smallporous ‘‘wound chambers’’ subcutaneously implanted into rats bySprugel et al. (1987). For the model simulations, 100×c(0, t) is theappropriate variable, with parameter values as in Fig. 3. Data forthe linear and axisymmetric models are shown alongside theexperimental data because the long cylindrical shape of theimplanted wound chambers is reminiscent of a combination of thetwo geometries. This is illustrated with remarkable clarity by theclearance curves shown above.
F. 4. Simulated and experimental data for wound contraction,plotted as the difference between the current and the fullycontracted wound area divided by the difference between the initialand the fully contracted area, expressed as a percentage. In themodel, this variable is 100×{(1+u(1, t))2−(1+u(1, a))2}/{1−(1+u(1, a))2}, where u(1, a) is the limit of u(1, t) as t:awhich exists and is negative when collagen kinetics are neglected(Section 4). The simulated data are obtained from the numericalsolutions of the axisymmetric model without collagen kinetics, withparameter values as in Fig. 3, except a=0.25. The experimentaldata are extracted from measurements of contracting circularwounds in rat skin by McGrath & Simon (1983). t=0 correspondsto day 3 in their experiments, since this is when the wound areabegins to decrease, indicating the beginning of the proliferativephase of healing (Section 2).
appears to be diffusion-driven, whereas the second,low-level decay may be partly due to production ofthe chemical by the cells which are infiltrating thewound.
4.5.
:
The model predicts that the contracted state isa transient (albeit long) phenomenon, in contrastto real wound healing, in which a permanentlycontracted state is normally observed.
To address this problem, we reconsider the collagenkinetics. At the dermal steady state, the collagenconcentration, r, is set to unity by the kinetic termsin (9). If these terms are neglected, however, then (9)simply reduces to a convection equation for r. Thisapproximation is validated by the relevant parametervalues [notably v and dp—see (9), Appendix andFig. 3 legend] and corroborated by the numericalsimulations [Fig. 3(d)]. Consequently, when n, m andc assume their dermal steady state values, r is notspecified and thus depends on the initial conditionsand on the subsequent healing dynamics. Inparticular, r(x, t) may therefore evolve to a spatially

. ET AL .122
inhomogeneous steady profile. From (10), thesteady-state displacement is given by the equation:
d2udx2+n
ddx 0 r
c2+r21−sru=0, u(0)=0,
dudx
(a)=0. (12)
The steady force balance equation (12) now containsa traction stress term (absent from (11) becausethere the collagen kinetics fix r=1). Thus, animportant result is deduced: a non-zero steady-statedisplacement profile is possible if the collagen kineticsare neglected. In this case, the variables evolve overthe 30-day time course in a similar manner to thesimulations shown in Fig. 3. Over several years,however, the collagen concentration, r, remains veryclose to its 30-day profile and does not return to itsuniform, dermal value. Moreover, the displacement,u, having exhibited contraction over the 30-daytime scale, does not relax back to zero, but remainspermanently contracted, as demonstrated in Fig. 6.Equivalent results are inferred from the axisymmetricformulation (not shown).
Together, these results are consistent with thebiology of wound healing because fibroplasiaand contraction occur when the wound is in atransient, proliferative phase of the healing process.On the much longer time scale of remodelling(Section 2), a number of other processes are involved(Clark, 1988; Mast, 1992) that are deliberatelyexcluded from the model. Thus, the modelcorrectly predicts the permanence of wound contrac-tion on the time scale over which it is intended to beapplied.
5. Summary and Discussion
The framework, functional forms (Section 3)and parameter value estimates (see Appendix)for the model presented in this paper are derivedfrom a detailed examination of experimentaland theoretical reports on fibroplasia and woundcontraction.
The novel inclusion of two cell types, myofibrob-lasts and fibroblasts, enables careful examination ofthe specialized and distinct roles of these cells infibroplasia and wound contraction (Section 2) and inpathological disorders of these processes. Explicitlyincorporating the variable dynamics of a genericchemical which regulates the cellular healing re-sponses is another novel aspect of the model that isevidently justified (Section 2).
These detailed considerations are central in theability of the model to elicit a biologically realisticrange of steady states (Section 4.1) representingunwounded dermis, a contracted wound and a varietyof fibrocontractive disease states, some of whichfeature elevated cell densities only (see Table 1 inSection 4.1), whilst others exhibit considerablyamplified cell, chemical and collagen levels,suggestive of chronic tissue pathologies. These areconsistent with the variety of fibrocontractive diseasesthat are increasingly well characterised clinically,although the underlying pathogenic mechanismsremain poorly understood (Skalli & Gabbiani, 1988;Rudolph & Vande Berg, 1991; Rudolph et al., 1992;Asmussen & Sollner, 1993; Murray, 1993). Thespatiotemporal nature of the pathogenesis, evolution,cessation and regression of these disease statesis currently under investigation (L. Olsen et al.,submitted).
F. 6. Numerical simulations of the linear wound model, using the method of lines and Gear’s method, showing the movement of tattoomarks to highlight tissue displacement, (a) as in Fig. 3(f ), and (b) without kinetics for r(x, t). Parameter values as in Fig. 3. The maximumtime shown, t=300, corresponds to a dimensional time of approximately ten months.

123
The existence and linear stability of these steadystates are given by specific inequalities in terms of themodel parameters (Section 4.1), which together withthe detailed parameter value estimates proposed (seeAppendix), yield a comprehensive predictive methodof determining the occurrence and nature of thesestates.
The results predict that the system evolves to asteady state which is reminiscent of normal dermisin terms of the cellular and chemical levels butwith a contracted steady displacement profile, ifand only if the kinetics for collagen are neglected(Section 4). This is biologically appropriate in thatcollagen kinetics are fundamental to the matrixremodelling phase of healing, operating on arelatively long time scale, rather than to fibroplasiaand contraction which are constitutive features of thetransient, proliferative phase, for which the model isdesigned.
Qualitatively, the numerical simulations corrobo-rate the normally observed processes of fibroplasiaand wound contraction in the healing of real andexperimental wounds (Sections 2 and 4). The linearand axisymmetric models are investigated, in whichthe wound domain is described in terms of one spatialcoordinate (Fig. 2, Section 3). The qualitative modelpredictions are analogous in the two geometries,notably in the dependence on collagen kinetics ofpermanently contracted state and in the form of thedisplacement profiles obtained.
Quantitatively, the numerical results are similar forthe two wound geometries, with relatively enhancedhealing responses (using the same parameter values)towards the wound centre in the axisymmetric casecompared to the linear case, in closer agreementwith experimental data. This improved accuracy isexpected, given the prevalence of circular wounds inexperimental studies from which many parametervalue estimates are inferred (see Appendix). Somenumerical results exhibit excellent correlation withexperimental data, however, as exemplified by the‘‘biphasic’’ clearance of the chemical from the woundand by the reduction in the wound area due tocontraction (details of which are provided in Section4.4). We reiterate that quantitative discrepanciesbetween the experimental and the model resultsmay arise due to significant variations in healingcharacteristics between different species. A probableexplanation is that many parameter values areestimated using data from a variety of in vitro andin vivo investigations using several species, includingrats, pigs and humans. The model can only predict,therefore, ‘‘averaged mammalian’’ fibroplasia andwound contraction.
Another quantity that could be usefully comparedwith experimental data is the ratio of myofibroblaststo total cells in the model, m/(n+m), over thetime-course of contraction. Rudolph (1979) measuredthis figure for excisional wound healing in pigs andrats and McGrath & Hundahl (1982) determined thisratio for a range of spatial locations in pig wounds.But even when the data are plotted so that the timeaxis is expressed as a percentage of the total time fromthe beginning to the end of wound contraction (in aparticular species or in the model), it is evident thatlarge qualitative disparities exist between differentspecies (Rudolph, 1979; McGrath & Hundahl, 1982).Therefore, comparing numerical solutions of themodel with experimental results for this quantity isnot reasonable.
The model results are not sensitive to smallvariations in any of the parameter values or initialconditions, provided points in parameter space arenot varied near bifurcation subspaces or singularphenomena (see Appendix).
An important observation from the model is thatdynamic cell and collagen gradients provide thedriving force for wound contraction (Section 4). Inthis, myofibroblasts are responsible for effectivelyamplifying and transmitting the fibroblast-derivedtraction forces through the tissue, and the collagenmatrix is a critical regulator.
L. O. expresses gratitude to the Engineering and PhysicalSciences Research Council of Great Britain for a ResearchStudentship award to support this work. Part of this workwas carried out while P.K.M. was visiting the School ofMathematics and Statistics, University of Sydney, Aus-tralia, and the Department of Mathematics, WilliamsCollege, Massachusetts, USA. This work was supported inpart by a grant from the London Mathematical Society. Wethank F. Arnold, J. Cook, P. Traqui, G. Cruywagen andJ. D. Murray for helpful discussions.
REFERENCES
A, B., B, D., L, J., R, M., R, K. &W, J. D. (1989). The Molecular Biology of the Cell. NewYork: Garland Publ. Inc.
A, F. & W, D. C. (1992). Angiogenesis in wound healing.Pharmac. Ther. 52, 407–422.
A, P. D. & S, B. (1993) Wound Care. Principles ofWound Healing. Hamburg: Beiersdorf medical Bibliothek.
A, B., F-C, C., D G, L.,C, C. & G, G. (1983). Abnormal behaviour ofcultured fibroblasts from nodule and nonaffected aponeurosis ofDupuytren’s disease. J. Cell. Physiol. 117, 353–361.
B, J. B. L. & H, E. D. (1975). The behaviour of fibroblastsfrom the developing avian cornea. Morphology and movementin situ and in vitro. J. Cell. Biol. 67, 400–418.
B, G. M. (1981). Physical Chemistry for the Life Sciences.New York: McGraw-Hill Inc.

. ET AL .124
B, E. A. & U, J. (1982). Special tissue collagen: Skin. In:Collagen in Health and Disease (M. Jayson & J. Weiss, eds)pp. 474–487. Edinburgh: Churchill Livingstone.
B, E., I, B. & M, C. (1979). Production of atissue-like structure by contraction of collagen lattices by humanfibroblasts of different proliferative potential in vitro. Proc. natn.Acad. Sci. U.S.A. 76, 1274–1278.
B-P, D. F., M, T. W., F, D. M. & R, R.(1984). Platelet-derived growth factor in vivo: Levels, activity andrate of clearance. Blood 64, 458–469.
C, R. A. F. (1988). Overview and general considerations ofwound repair. In: The Molecular and Cellular Biology of WoundRepair (R. A. F. Clark & P. M. Henson, eds) pp. 3–34. NewYork: Plenum.
C, R. A. F. (1989). Wound repair. Curr. Opin. Cell Biol.,1, 1000–1008.
C, R. A. F. (1991). Growth factors and wound repair. J. Cell.Biochem., 46, 1–2.
C, R. A. F. (1993). Regulation of fibroplasia in cutaneouswound repair. Am. J. Med. Sci. 306, 42–48.
D, R. & W, N. A. (1991). The cell proliferation kineticsof the epidermis. In: Physiology, Biochemistry and MolecularBiology of the Skin (L. A. Goldsmith, ed.) pp. 239–265. NewYork: Oxford University Press.
E, H. P. & R, J. B. M. (1990). Cell locomotionforces versus cell contraction forces for collagen latticecontraction: An in vitro model of wound contraction. Tiss. Cell.22, 407–417.
F, Y. C. (1981) Biomechanics. New York: Springer-Verlag.G, G. (1992). The biology of the myofibroblast. Kidney Int.,
41, 530–532.G, G., M, G. & R, G. B. (1973). The fibroblast as
a contractile cell: The myofibroblast. In: Biology of the Fibroblast(J. Pikkarainen & K. Kulonen, eds) pp. 139–154. New York:Academic.
G, G., R, G. B. & M, G. (1971). Presence ofmodified fibroblasts in granulation tissue and their possible rolein wound contraction. Experientia 27, 549–550.
G, G. R. (1992). Chemoattractants and growth factors.In: Wound Healing: Biochemical and Clinical Aspects (I. K.Cohen, R. F. Diegelmann, & W. J. Lindblad, eds) pp. 237–246.Philadelphia: Saunders.
H, J. S., H, S. S. & D, T. F. (1984). Specificcovalent binding of platelet-derived growth factor to humanplasma a2-macroglobulin. Proc. natn. Acad. Sci. U.S.A.81, 342–346.
H, J. S., O, T. J. & H, S. S. (1988). The roleof growth factors in tissue repair I. Platelet-derived growthfactor. In: The Molecular and Cellular Biology of Wound Repair(R. A. F. Clark & P. M. Henson, eds) pp. 243–251. New York:Plenum.
J, J. J. (1992). Collagen degradation. In: Wound Healing:Biochemical and Clinical Aspects (I. K. Cohen, R. F. Diegelmann& W. J. Lindblad, eds) pp. 177–194. Philadelphia: Saunders.
J, R. W. & H, T. K. (1992). Overview of postnatalwound healing. In: Fetal Wound Healing (N. S. Adzick & M. T.Longaker, eds) pp. 25–52. New York: Elsevier.
L, L. D. & L, E. M. (1970). Theory of Elasticity.New York: Pergamon.
L� , J., L, M., N, J., L, C., G, B.& H, C.-H. (1992). Effects of homodimeric isoforms ofplatelet-derived growth factor (PDGF-AA and PDGF-BB) onwound healing in rat. J. Surg. Res. 53, 596–601.
L, M. T., A, N. S. & E, H. P. (1992). Fetalwound contraction. In: Fetal Wound Healing (N. S. Adzick &M. T. Longaker, eds) pp. 149–160. New York: Elsevier.
M, P., H-W, J. & MC, J. (1992).Growth factors and cutaneous wound repair. Prog. Gro. Fact.Res. 4, 24–44.
M, B. A. (1992). The skin. In: Wound Healing: Biochemicaland Clinical Aspects (I. K. Cohen, R. F. Diegelmann & W. J.Lindblad, eds) pp. 344–355. Philadelphia: Saunders.
MG, M. H. & H, S. A. (1982). The spatial andtemporal quantification of myofibroblasts. Plast. Reconstr. Surg.69, 975–983.
MG, M. H. & S, R. H. (1983). Wound geometryand the kinetics of wound contraction. Plast. Reconstr. Surg.72, 66–72.
M, C. J. & P, W. J. (1992). Fibroblast proliferation.In: Wound Healing: Biochemical and Clinical Aspects(I. K. Cohen, R. F. Diegelmann & W. J. Lindblad, eds)pp. 63–76. Philadelphia: Saunders.
M, J. C. (1993). Scars and keloids. Dermatol. Clin.11, 697–708.
M, J. D. (1989). Mathematical Biology. New York:Springer-Verlag.
M, J. D., M, P. K. & T, R. T. (1988).Mechanochemical models for generating biological pattern andform in development. Phys. Rep. 171, 59–84.
M, J. D. & O, G. F. (1984). Cell traction modelsfor generating pattern and form in morphogenesis. J. math. Biol.19, 265–279.
M, J. D., O, G. F. & H, A. K. (1983). Amechanical model for mesenchymal morphogenesis. J. math.Biol. 17, 125–129.
O, G. & R, R. (1968). Human wound repair: I. Epidermalregeneration. J. Cell Biol. 39, 135–151.
O, G. F. (1991). Structure of the skin. In: Physiology,Biochemistry and Molecular Biology of the Skin (L. A.Goldsmith, ed) pp. 3–62. New York: Oxford University Press.
O, G. F., M, J. D. & H, A. K. (1983). Mechanicalaspects of mesenchymal morphogenesis. J. Embryol. explMorphol. 78, 83–125.
P, G. F., B, D. & M, T. A. (1991a). Quantitativeanalysis of inflammatory cell influx, procollagen type I synthesis,and collagen cross-linking in incisional wounds: Influenceof PDGF-BB and TGF-b1 therapy. J. Lab. Clin. Med.,117, 373–382.
P, G. F., V B, J., R, R., T, T.& M, T. A. (1991b). Platelet-derived growth factor-BBand transforming growth factor beta1 selectively modulateglycosaminoglycans, collagen, and myofibroblasts in excisionalwounds. Am. J. Pathol. 138, 629–646.
R, E. W., B-P, D. F. & R, R. (1990).Platelet-derived growth factor. In: Handbook of ExperimentalPharmacology, Vol. 95 part I: Peptide Growth Factors and theirReceptors (M. B. Sporn & A. B. Roberts, eds) pp. 173–262.Heidelberg: Springer-Verlag.
R, A. B., S, M. B., A, R. K., et al. (1986).Transforming growth factor type b: Rapid induction of fibrosisand angiogenesis in vivo and stimulation of collagen formationin vitro. Proc. natn. Acad. Sci. U.S.A. 83, 4167–4171.
R, R., R, R. W. & B-P, D. F. (1986). The biologyof platelet-derived growth factor. Cell 46, 155–169.
R, R (1979). Location of the force of wound contraction.Surg. Gynecol. Obstet. 148, 547–551.
R, R. (1980). Contraction and the control of contraction.World J. Surg. 4, 279–287.
R, R., V B, J. & E, H. P. (1992). Woundcontraction and scar contracture. In: Wound Healing: Biochemicaland Clinical Aspects (I. K. Cohen, R. F. Diegelmann &W. J. Lindblad, eds) pp. 96–114. Philadelphia: Saunders.
R, R. & V B, J. (1991). The myofibroblast inDupuytren’s contracture. Hand Clin. 7, 683–692.
S, N., B, J. G., K, J., Y, M., C, J. W.,C, P. & F, Jr. A. R. (1993). Platelet-derivedgrowth factor indirectly stimulates angiogenesis in vitro. Am. J.Pathol. 142, 1119–1130.
S, H., G, G., S� S., S, E. & M,G. R. (1982). Platelet-derived growth factor is chemotactic forfibroblasts. J. Cell. Biol. 92, 584–588.
S, M., F, D. M. & F, M. W. J. (1992). Controlof scarring in adult wounds by neutralising antibody totransforming growth factor b. Lancet 339, 213–214.

125
S, J. A., M, P., M, J. D. & L, J. (1992).Mathematical models of wound healing in embryonic and adultepidermis. IMA J. Math. Appl. Med. Biol. 9, 177–196.
S, J. A. & M, J. D. (1992). Epidermal woundhealing: The clinical implications of a simple mathematicalmodel. Cell Transpl. 1, 365–371.
S, J. A., S, E. H. & M, J. D. (1993). Chemicalcontrol of eukaryotic cell movement: A new model. J. theor.Biol. 162, 23–40.
S, I. I., K, D. W., K, D. M. & C, R. A. F.(1984). In vivo co-distribution of fibronectin and actin fibers ingranulation tissue: Immunofluorescence and electron microscopestudies of the fibronexus at the myofibroblast surface. J. Cell.Biol. 98, 2091–2106.
S, O. & G, G. (1988). The biology of themyofibroblast. Relationship to wound contraction and fibrocon-tractive diseases. In: The Molecular and Cellular Biology ofWound Repair (R. A. F. Clark & P. M. Henson, eds)pp. 373–402. New York: Plenum.
S, O., S, W., S, T., L, R.,M, D., P, B. & G, G. (1989). Myofibrob-lasts from diverse pathologic settings are heterogeneous in theircontent of actin isoforms and intermediate filament proteins.Lab. Invest. 60, 275–285.
S, J. M. & C, W. J. (1985). Wound contraction.Correlations between the tension generated by granulationtissue, cellular content and rate of contraction. Q. J. expl.Physiol. 70, 539–548.
S, K. H., MP, J. M., C, A. W. & R, R.(1987). Effects of growth factors in vivo. Am. J. Pathol. 129,
601–613.S, D. & H, A. K. (1982). Connective tissue
morphogenesis by fibroblast traction: I: Tissue culture obser-vations. Dev. Biol. 90, 383–398.
T, R. T. & M, J. D. (1992). Continuum model offibroblast-driven wound contraction: Inflammation-mediation.J. theor. Biol. 158, 135–172.
U, J. (1984). Interstitial collagens. In: Biology of theIntegument, Vol. 2: Vertebrates (J. Bereiter-Hahn, A. G.Matoltsky & K. S. Richards, eds) pp. 800–809. Berlin:Springer-Verlag.
V B, J. S., R, R., P, W. L. & D,D. R. (1989). Comparative growth dynamics and actinconcentration between cultured human myofibroblasts fromgranulating wounds and dermal fibroblasts from normal skin.Lab. Invest. 61, 532–538.
W, G. F. & Z, B. R. (1992). Angiogenesis. In: WoundHealing: Biochemical and Clinical Aspects pp. 77–95. Philadel-phia: Saunders.
APPENDIX
In view of the large number of parameters in themodel [Section 3: see eqns (1–5)], accurate parametervalue estimation is a vital part of the modellingprocess. An important step in this process is toreduce the number of dimensional parameters by thefollowing simplifying assumptions:
, The half-maximal concentrations for the self-regulated production and consumption of thechemical are equal, since the same cell surfaceinteraction mechanism occurs in the two cases,and we set Gp=Gc=.G.
, The ratio of the roles of myofibroblasts tofibroblasts in the production of the chemical isequal to that in the consumption of the chemical,so zp=z=.z.
, A similar argument for the roles of the two celltypes in collagen biosynthesis and degradationyields hb=hd=.h.
For the remaining parameters, we first estimatevalues for the typical scalings which are used tonondimensionalize the variables (see below).L: A typical length scale of an excisional dermalwound is 1 cm.T: The progress of a typical wound through theinflammatory and proliferative phases of healing(Section 2) is normally measured in days, so we setT=1 day.n0: Human dermal fibroblasts generally have dimen-sions of the order 100 mm×100 mm×10 mm. Further-more, fibroblasts are sparse in normal adult dermis(Morgan & Pledger, 1992), and stipulating that thereare, on average, ten cells per cubic millimetre, yieldsthe value n0=104 ml−1.c0: PDGF is stored within platelets in the blood-stream at low concentrations—reports suggest15–50 ng ml−1 (Huang et al., 1988). The concen-trations released at the wound by platelets and byactivated inflammatory cells should not, therefore,exceed these levels, and we propose c0=10 ng ml−1.The initial chemical concentration profile iscinit (x)=c0(1+x4)−1.r0: The density of the human body is similar to thatof water, namely 1 g ml−1. By mass, 1 ml of tissuecontains approximately 70% water, 15% fat and 15%other substances, of which roughly three-quarters iscollagen in the case of dermal tissue (Bauer & Uitto,1982). Hence, r010.1g ml−1 is a valid order ofmagnitude estimate.
With these typical scales, the following nondimen-sional variables are introduced:
x=xL
, t =tT
, n=nn0
, m=mn0
, c=cc0
,
r=r
r0, u=
uL
.
Turning to the dimensional model parameter values:Dn : Electron microscope studies of migrating humanfibroblasts suggest that the maximum rate of ran-dom motility is approximately 1 mm min−1 (Bard &Hay, 1975), yielding a diffusion coefficient of1.7×10−10 cm2 s−1.an : In the absence of reported quantitative studies todetermine the chemotactic parameters for dermalfibroblasts, we conducted numerical simulations for a

. ET AL .126
range of values of an . Because fibroplasia fundamen-tally depends on inflammatory stimuli [11], we suggestthat the chemotactic flux of fibroblasts into thewound site should predominate over the randomdiffusive flux and therefore set an=5 ng cm−1 day−1.bn : Concentrations as low as 1 ng ml−1 of PDGFelicit significant fibroblast chemotaxis (Seppa et al.,1982), and experimental data indicate that theconcentration for the half-maximal response is about2 ng ml−1 (Grotendorst, 1992).rn , rnmax, K and dn : The basal growth rate, rn , of apopulation of fibroblasts is difficult to determineexperimentally because the population appears to beconstant, for the following reason: when m=0 andc=0, there is a steady-state cell density, n*, given bythe equation: rnn*(1−n/K)−dnn*=0, implying eithern*=0 or n*=K(1−dn /rn ). Because m=0 and c=0are characteristic of normal dermis, we assume thatn*=n0, the normal dermal fibroblast density. Thus rn
may be determined once K and dn are known.The maximal fibroblast density, K, is the density at
which cell proliferation is totally inhibited. Exper-imental growth curves and various studies usinghigh-density fibroblast populations indicate a value ofthe order of 106 cells per millimetre, so we takeK=106 ml−1. As a check, this implies that eachfibroblast occupies a volume of at least (100 mm)3,which is consistent given that the maximum lineardimension of a fibroblast is about 100 mm).
Cultured dermal fibroblasts have an averagelifespan of approximately 40 population doublings(Azzarone et al., 1983), so denoting the maximaldoubling time by T2 yields dn=(ln 2)/(40×T2).Estimates for T2 are typically around 18 h [1, 32],which implies dn10.02 day−1. Hence, rn=dn (1−n0/K)−110.99 dn=0.0198 and rnmax=(ln 2/T2)−rn10.9.C1/2: The chemical concentrations required to en-hance fibroblast proliferation are somewhat higherthan those for chemotactic responses (Grotendorst,1992; Lepisto et al., 1992). Experimental evidenceindicates that half-maximal enhancement corre-sponds to concentrations of about 10 ng ml−1
(Grotendorst, 1992).k1max, Ck and k2: Quantitative data on the rates ofconversion between fibroblasts and myofibroblastsare not available, so the following intuitive assump-tions are proposed: first, the disappearance ofmyofibroblasts at the end of contraction is predomi-nantly due to reversion to the fibroblast phenotyperather than cell death, so k2qdm ; second, the‘‘half-life’’ of the myofibroblast to fibroblast conver-sion is less than a day (this process primarily involvesintracellular modifications, so the distances involvedare very small and signals are transmitted quickly).
This half-life is (ln 2)/k2, so we take k211 day−1; third,given the intracellular growth factor-driven eventsinvolved, the maximal rate of conversion offibroblasts to myofibroblasts is faster than the reverseprocess, so we take k1max110 day−1; finally, therelatively low chemical concentrations in the woundshould stimulate myofibroblast production, so areasonable order-of-magnitude estimate isCk=1 ng ml−1.er : Growth curves of myofibroblasts cultured fromcontracting wounds indicate that the growth rate islower (50–100%) than for normal dermal fibroblastcultures (Vande Berg et al., 1989; Rudolph & VandeBerg, 1991). The variability in the experimental datais due to the heterogeneity of the cellular compo-sition of wounds (Snowden & Cliff, 1985; Skalli &Gabbiani, 1988). In particular, the myofibroblast/fibroblast density ratio varies with time and spaceduring healing (McGrath & Hundahl, 1982). Assum-ing that a population of ‘‘pure’’ myofibroblastsproliferates at half the rate of fibroblasts, we estimateer=0.5.l: There is currently no data on death rates ofmyofibroblasts, qualitatively or quantitatively. Usingthe following inequalities: dnQdm=ldnQk2 (derivedfrom earlier estimates), however, and the valuesprovided for the bounds, we obtain 1QlQ50 andchoose l=10.Dc : Although direct experimental measurement ofthe diffusivity of PDGF is lacking, its molecularweight has been consistently recorded at around30 000 Da, from which the following theoreticalargument can be used to estimate a value for Dc . Formany large proteins, such as wound growth factors,in aqueous solution at a particular temperatureand pressure, DcAM−1/3
c is a good approximation(Barrow, 1981), where Mc is the molecular weight ofthe chemical. The constant of proportionality isunknown, but if c' represents a second chemical, thenDc1(Mc'/Mc )1/2 Dc'. Data is available for epidermalgrowth factor, whose molecular weight is 6045 anddiffusion coefficient is 9.75×10−7 cm2 s−1. By theargument above, the estimated value of Dc is5.7×10−7 cm2 s−1.p, k'p , k'c and G: The functional forms for theproduction and consumption of the chemical in (3)involving these parameters are derived from the sche-matic reactions:
R+C04k'p
R+pC and R+C04k'c
R
respectively, where R represents a free cell-surfacereceptor molecule, C a chemical molecule, p is an(unknown) positive integer and k'p and k'c are the rate

127
constants. Denoting the concentrations of R by r andC by c, then the law of mass action implies1c/1t=( p−1)k'p rc−k'c rc. Assuming that theconcentration of bound receptors, rbound, is asaturating monotonic increasing function of c, rbound=rtotalc/(G+c), say [see also Sherratt et al., (1993)],where the total receptor concentration is rtotal=rbound+r, then r=Grtotal/(G+c). Finally, rtotal isproportional to cell density, n, giving the expressionsin (3).
The only information on the values of theseparameters is that they are positive. By consideringthe existence and linear stability of steady states(Section 4.1), bounds may be imposed on the value ofa dimensionless parameter, kc , which is related to thedimensional quantity ( p−1)k'p−k'c .z: Myofibroblasts also express cell surface receptorsfor PDGF (Sato et al., 1993) implying that these cellsalso regulate the chemical concentration, so zq0.Without evidence to suggest that myofibroblastsmetabolize PDGF at a different rate of fibroblasts, wetake z=1.dc : When excess PDGF is released from platelets andother stores, it quickly forms a complex with theplasma binding protein a2-macroglobulin with ahalf-life of 4 min (Huang et al., 1984, 1988). Thisbound complex is recognized by a receptor fora2-macroglobulin on the surface of liver cells, and israpidly cleared from plasma with a half-life of only2 min (Bouen-Pope et al., 1984), a figure which yieldsthe estimate dc=500 day−1. It remains unknown,however, whether (a) some of the chemical releasedduring healing is unavailable for binding toa2-macroglobulin, for example if there are insufficientlevels of the binding protein locally at the wound site,and (b) the bound PDGF-a2-macroglobulin complexmay be recognized by fibroblasts and myofibroblastsand the PDGF subsequently internalized andmetabolized. In these scenarios dc could be consider-ably smaller, with significant consequences for themodel solutions.rr , Rr and dr : As stated in Section 4.1., a uniformsteady state (n, m, c, r, u)=(n0, 0, 0, r0, 0) exists,representing unwounded dermis. Therefore, if three ofthe parameters rr : Rr , dr and r0 are known, then sois the fourth, then so is the fourth, by the virtue ofrr (R2
r+r20 )−1−drr0=0, from equation (4) in Section
3. The half-life of collagen turnover in the adulthuman dermis is 2.5 years (Uitto, 1984), which yieldsdr=(ln 2)/(365×2.5×n0) day−118×10−8 day−1. Weestimate Rr using the following intuitive argument.The sigmoidal biosynthesis inhibition function,(R2
r+r2)−1, is half of its maximal (at r=0) value whenr=Rr . We assume that this substantial downregula-
tion of its production occurs at abnormally highcollagen concentrations, so that Rrq1, but not solarge that biosynthesis inhibition is essentiallynegligible. In the numerical simulations, we setRr=3r0. Finally, rr is found by using the estimates forthe other parameters in the above equation.rrmax: Studies using implanted dermal wound‘‘chambers’’ have indicated that PDGF alone doesnot enhance collagen biosynthesis per cell (Lepistoet al., 1992). Transforming growth factor-beta(TGF-b), however, has been shown to markedlyincrease collagen synthesis per cell (Roberts et al.,1986; Pierce et al., 1991a, b). The synergistic effects ofthese two growth factors may accelerate collagenbiosynthesis up to tenfold, so rrmax=10rr is areasonable estimate.Cr : Experimental data suggest that half-maximalenhancement of collagen biosynthesis occurs atTGF-b concentrations of the order of 1 ng ml(Roberts et al., 1986), thus giving an estimate forCr . h: Cultured myofibroblasts are biosyntheticallyactive (Skalli & Gabbiani, 1988), implying thathq0, and observations have suggested that my-ofibroblasts secrete twice as much type I collagen asfibroblasts (Rudolph & Vande Berg, 1991), so thath=2.m1, m2, E' and n': These mechanical parameters arerelated to the bulk and shear viscosities, Young’smodulus and Poisson’s ratio for the adult dermis; forwhich data is not readily available—a ubiquitousproblem in mechanochemical models. Consequently,(dimensionless) parameter variation analyses wereconducted numerically, from which approximateranges of parameter values for which the model yieldsbiologically and mathematically valid solutions werederived.t0: The traction force per unit of collagen concen-tration and fibroblast cell density has been studied indetail both mathematically (Murray & Oster, 1984;Tranquillo & Murray, 1992) and experimentally(Snowden & Cliff, 1985; Stepak & Harris, 1982). Datahas not been given, however, for cell traction forcesin a three-dimensional collagen gel—a system whichis currently attracting considerable research. Atpresent, therefore, numerical experiments are againused to suggest order-of-magnitude estimates.j: Sharp myofibroblast enhancement of tractionis proposed per unit of cell density, motivated bythe strong correlation of myofibroblast densityand the rate of contraction (Rudolph, 1979;McGrath & Hundahl, 1982; Skalli & Gabbiani, 1988),and we set j=10. j and t0 are intimately relatedbecause their product is the coefficient of the

. ET AL .128
myofibroblast-enhanced component of the cell trac-tion term in the force balance equation (5).s: This parameter measures the strength of the linearrestoring forces acting against tissue displacement,but quantitative data on the magnitude of these forcesare not available. Therefore, acceptable parameterestimates were investigated numerically, in conjunc-tion with the other parameters in the force balanceequation, as above.Rt : The following arguments, based upon (a) thebiomechanics of wound contraction and (b) the formof the cell traction stress term in the force balanceequation, are used to estimate this parameter value.In vitro investigations using collagen gels havedemonstrated that the rate of contraction isproportional to the cell density (Bell et al., 1979),which is consistent with the proposed tractionfunction, since tAn. Further, the rate of contractionis inversely proportional to the collagen concentration(Bell et al., 1979). Significantly, tAr−1 in the limit asRt:0 for values of rqRt . Thus the given form agreeswith the experimental data of Bell et al. (1979), if Rt
is sufficiently small. Next, the mathematical conse-quences of the traction term are considered bywriting:
t(r)=r
R2t+r2 , where r=r(x, t).
The momentum flux is given by the cell traction stressgradient:
1t
1x=
R2t−r2
(R2t+r2)2
1r
1x.
As x increases from zero, the wound centre, we expectr to be a monotonically increasing function of x.Hence, 1t/1x and R2
t−r2 have the same sign. In orderto exhibit wound contraction, the traction forces mustact towards the wound centre, so the sign of 1t/1xmust be negative. Thus, an approximate upper boundfor Rt is the minimum value of r for xq0, so RtQrinit .Without further guidance, we choose Rt=0.5 rinit .rinit : The concentration of type I collagen in thewound at the onset of contraction is a small fractionof that of normal dermis, and in the absence ofquantitative evidence from the literature, we set
rinit=0.01r0.Finally, the following nondimensionalization
scheme is used to relate the parameters in the modelequations (6–10) to those in (1–5):
D n=DnTL2 a=
anTL2c0
b =bn
c0s=rnT A =
rnmax
rn
B =C1/2
c0g=
n0
Kk1=k1maxT C =
Ck
c0k2=k2T
d =dnT es=er l =dm
dnD c=
DcTL2
kc=((p−1)k'p−k'c )Tn0
gc=G
c0z =z d c=dcT v=
rrR2rTn0
r30
P=rrmax
rr
Q =Cr
c0f =
Rr
r0d r=drTn0 h=h
m=m1+m2
E'(1+n')T
n=t0R2
tn0
E'(1+n')r0j =jn0 c =
Rt
r0s=
sL2r0
E'(1+n')
rinit=rinit
r0cinit (x)=
cinit (x)c0
We remark that parameter variation sensitivityanalyses were conducted numerically for all of themodel parameters (using the dimensionless quan-tities). For the parameters whose values wereestimated from experimental data or theoreticalcalculations, small variations in these values resultedonly in small quantitative changes in the numericalsolutions of the system. The remaining parameters,whose values were not so readily estimated, werechosen such that in parameter space, the system wasnot near a region of sensitive behaviour in thenumerical simulations, such as bifurcation surfaces.Hence, all of the parameter values used for the resultsin this paper (see Fig. 3 legend) required relativelylarge deviations in order to elicit significantly alterednumerical results.


















