
A cladistic analysis and taxonomic revision of the ...
63
Acta Zool. Fennica 213: 1–63 ISBN 951-9481-58-3 ISSN 0001-7299 Helsinki 11 December 2001 © Finnish Zoological and Botanical Publishing Board 2001 A cladistic analysis and taxonomic revision of the Plesiosauria (Reptilia: Sauropterygia) Frank Robin O’Keefe Department of Anatomy, New York College of Osteopathic Medicine, Old Westbury, New York 11568, U.S.A Received 13 February 2001, accepted 17 September 2001 O’Keefe F. R. 2001: A cladistic analysis and taxonomic revision of the Plesio- sauria (Reptilia: Sauropterygia). — Acta Zool. Fennica 213: 1–63. The Plesiosauria (Reptilia: Sauropterygia) is a group of Mesozoic marine reptiles known from abundant material, with specimens described from all continents. The group originated very near the Triassic–Jurassic boundary and persisted to the end- Cretaceous mass extinction. This study describes the results of a specimen-based cladistic study of the Plesiosauria, based on examination of 34 taxa scored for 166 morphological characters. The Pliosauroidea is found to by polyphyletic due to the inclusion of the Polycotylidae; this second clade is instead a member of the Plesiosauroidea, and thus more closely related to elasmosaurs than to other ‘plio- saurs’. Characters of body proportion such as neck length and head size are very labile, with the ‘pliosauromorph’ body plan evolving three times, while extremely long necks evolved in two clades. Characters from the entire skeleton support these relationships, although characters of the skull roof and palate are especially useful. Lastly, a new genus and species, Hauffiosaurus zanoni, is named based on German material of Toarcian age. Introduction The term ‘plesiosaur’, meaning ‘near-lizard’, is not an informative name from a modern per- spective. However, when Conybeare (1822) coined the term to describe fossils from the English Lias little was known concerning any extinct reptile. The realization that plesiosaurs were a completely extinct group was significant at a time when the occurrence of extinction itself was uncertain (Taylor 1997). These ‘near-rep- tiles’ were named at a time when there was no need, and no context, for a more specific term. The Plesiosauria was a clade of Mesozoic marine reptiles that evolved from stem-group sauropterygians very near the Triassic–Jurassic boundary (reviewed in Rieppel 1997a, Rieppel 2000). The clade diversified during the Jurassic and Cretaceous, only to join the dinosaurs as casualties of the end-Cretaceous mass extinction (Romer 1966). Plesiosaurs were advanced over their ‘nothosaur’-grade forebearers in the evolu-
Transcript of A cladistic analysis and taxonomic revision of the ...

Acta Zool. Fennica 213: 1–63 ISBN 951-9481-58-3 ISSN 0001-7299Helsinki 11 December 2001 © Finnish Zoological and Botanical Publishing Board 2001
A cladistic analysis and taxonomic revision ofthe Plesiosauria (Reptilia: Sauropterygia)
Frank Robin O’Keefe
Department of Anatomy, New York College of Osteopathic Medicine, OldWestbury, New York 11568, U.S.A
Received 13 February 2001, accepted 17 September 2001
O’Keefe F. R. 2001: A cladistic analysis and taxonomic revision of the Plesio-sauria (Reptilia: Sauropterygia). — Acta Zool. Fennica 213: 1–63.
The Plesiosauria (Reptilia: Sauropterygia) is a group of Mesozoic marine reptilesknown from abundant material, with specimens described from all continents. Thegroup originated very near the Triassic–Jurassic boundary and persisted to the end-Cretaceous mass extinction. This study describes the results of a specimen-basedcladistic study of the Plesiosauria, based on examination of 34 taxa scored for 166morphological characters. The Pliosauroidea is found to by polyphyletic due to theinclusion of the Polycotylidae; this second clade is instead a member of thePlesiosauroidea, and thus more closely related to elasmosaurs than to other ‘plio-saurs’. Characters of body proportion such as neck length and head size are verylabile, with the ‘pliosauromorph’ body plan evolving three times, while extremelylong necks evolved in two clades. Characters from the entire skeleton support theserelationships, although characters of the skull roof and palate are especially useful.Lastly, a new genus and species, Hauffiosaurus zanoni, is named based on Germanmaterial of Toarcian age.
Introduction
The term ‘plesiosaur’, meaning ‘near-lizard’, isnot an informative name from a modern per-spective. However, when Conybeare (1822)coined the term to describe fossils from theEnglish Lias little was known concerning anyextinct reptile. The realization that plesiosaurswere a completely extinct group was significantat a time when the occurrence of extinction itselfwas uncertain (Taylor 1997). These ‘near-rep-
tiles’ were named at a time when there was noneed, and no context, for a more specific term.
The Plesiosauria was a clade of Mesozoicmarine reptiles that evolved from stem-groupsauropterygians very near the Triassic–Jurassicboundary (reviewed in Rieppel 1997a, Rieppel2000). The clade diversified during the Jurassicand Cretaceous, only to join the dinosaurs ascasualties of the end-Cretaceous mass extinction(Romer 1966). Plesiosaurs were advanced overtheir ‘nothosaur’-grade forebearers in the evolu-

O’Keefe • ACTA ZOOL. FENNICA No. 2132
tion of wing-shaped fore and hind flippers thatgenerated thrust via lift as well as drag (Godfrey1984, Lingham-Soliar 2000). Plesiosaurs wereunusual among marine reptiles because theyused no axial flexure when generating thrust(Storrs 1993); plesiosaurs are unique amongaquatic tetrapods in the elaboration of both foreand hind limbs for lift-based thrust generation.The evolution of this novel locomotor appara-tus, and the pelagic lifestyle it allowed, led torapid diversification and the evolution of differ-ent morphotypes linked to prey size (Massare1988, O’Keefe 2002).
Plesiosaur taxonomy
The taxonomy of the Infra-order Plesiosauria(de Blainville 1835) suffers from the compara-tive taxonomic chaos of the Victorian era. How-ever, Andrews (1910, 1913), Welles (1943,1952), Tarlo (1960), Persson (1963), and Brown(1981) have all revised plesiosaur taxonomy.All formalized the plesiosauromorph/pliosauro-morph dichotomy in their classifications, fol-lowing the lead of earlier workers such as Owen(1841), Lydekker (1889b), and Seeley (1892).Until the work of Carpenter in 1997, Williston(1925) was the only worker to break with theaccepted, morphotype-based taxonomy. Plesio-saur relationships have been considered recentlyby Brown and Cruickshank (1994). These work-ers also retain the traditional dichotomy. Thecurrent taxonomy is listed below, based onTarlo (1960), Persson (1963), Brown (1981,1993), and Brown and Cruickshank (1994).
Diapsida Osborn, 1903Sauropterygia Owen, 1860
Infra-Order Plesiosauria de Blainville, 1835Superfamily Plesiosauroidea Welles, 1943
Family Plesiosauridae Gray, 1825Family Elasmosauridae Cope, 1870Family Cryptoclididae Williston, 1925Family Cimoliasauridae Delair, 1959
Superfamily Pliosauroidea Welles, 1943Family Pliosauridae Seeley, 1874Family Rhomaleosauridae Kuhn, 1961Family Polycotylidae Williston, 1908
Early taxonomies classified plesiosaurs withrespect to two body proportion morphotypes(classifications reviewed in Welles 1943). Thesemorphotypes were the long-necked, small-headed‘plesiosaurs’ (here termed plesiosauromorphs)and the large-headed, short-necked ‘pliosaurs’(here termed pliosauromorphs). However, as ear-ly as 1907 Williston suggested that the short neckin ‘pliosaurs’ might have evolved at least twice.The validity of the pliosauromorph/plesiosauro-morph dichotomy was challenged more recentlyby Carpenter (1997), and preliminary cladisticanalyses by Bardet (1998) and Druckenmiller(1998) challenged the monophyly of the Super-family Pliosauroidea as traditionally defined. Thisstudy is a specimen-based analysis whose goal isthe elucidation of the phylogeny of the Plesiosau-ria. The terminal taxa in this analysis are genera,as higher-level relationships are the primary goal.The principal finding is the polyphyly of thePliosauroidea as traditionally defined, by inclu-sion of the Polycotylidae. Placement of this fami-ly in the Plesiosauroidea also results in somereorganization of that taxon.
Phylogenetic context
The Sauropterygia is a clade of basal diapsids,more closely related to lepidosaurs than archo-saurs but near this basal dichotomy (see Rieppel& Reisz 1999 for a review). Some recent workhas indicated that Testudines is the sister groupof Sauropterygia, although this work is contro-versial (Rieppel & deBraga 1996, Lee 1997, deBraga & Rieppel 1997, Rieppel & Reisz 1999).The comparative anatomy and phylogeny ofTriassic sauropterygians have been the subjectof intense study over the last decade. Storrs(1991) published a cladogram of stem-groupsauropterygians in the context of his redescrip-tion of Corosaurus, at that time the only Triassicsauropterygian known from the New World.Storrs also reviewed stem-group sauroptery-gians. This phylogeny underwent exhaustive re-vision by Rieppel, who engaged in a program ofredescription, taxonomic revision, and biogeog-raphy of all stem-group sauropterygians (re-viewed in Rieppel 1999, Rieppel 2000, Rieppel

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 3
1994a, 1998, Rieppel & Wild 1996, and espe-cially Rieppel 2000 are good entrance points toRieppel’s literature on the group). Field work byRieppel, Storrs, and others also yielded remainsof a new sauropterygian from Nevada (Sander etal. 1997). This new genus (Augustasaurus) is apistosaurid, and is significant because pistosau-rids are thought to be the sister group to thePlesiosauria (Sues 1987). The only other knownpistosaurid is Pistosaurus from the Triassic ofGermany, described by Meyer (1839) and Ed-inger (1935) from two skulls, the more completeof which is now lost. Rieppel (1994a) rede-scribed the remaining skull, while Sues (1987)described the postcranium of Pistosaurus indetail.
The cladogram topologies obtained by Riep-pel varied as his research program evolved.More definitive versions from later publicationssuch as Rieppel (1998, 1999, 2000) agree on thebroad outline of sauropterygian phylogeny. InRieppel’s topology, placodonts are the firstbranch within the clade Sauropterygia, makingthem the outgroup to all other ‘nothosaur’-gradesauropterygians plus the Plesiosauria (clade Eo-sauropterygia). Three subclades are found with-in this group, one comprised of pachypleuro-saurs such as Dactylosaurus and Neusticosaurus(clade Pachypleurosauroidea). A second clade iscomposed of the ‘nothosaurs’ Simosaurus, Not-hosaurus, and related taxa (clade Nothosauria),and a third contains the Plesiosauria and plesio-saur-like ‘nothosaurs’ (clade Pistosauroidea; no-menclature from Rieppel 1998, Rieppel 1999,2000). Clade Pistosauroidea contains the generaCymatosaurus, Corosaurus, Pistosaurus, andthe closely related Augustasaurus. In the follow-ing discussion, stem-group eosauropterygians(i.e., eosauropterygians minus the Plesiosauria)are refered to using the colloquial terms ‘notho-saur’ or ‘‘nothosaur’-grade taxa’. When the termnothosaur appears without quotes, it refers onlyto the monophyletic clade Nothosauria.
Anatomical background
Some introductory comments concerning theanatomy of plesiosaurs and their sauropterygian
antecedents are necessary. This section discuss-es only areas of variability, especially concern-ing the skull and limb girdles. Specific charac-ters and their states are discussed in Appendix 1.
Cranial anatomy
No comprehensive review of the plesiosaurskull has been attempted since the work ofAndrews (1910, 1913). Brown (1981, 1993),and Brown and Cruickshank (1994) discussedthe skull roof of Cryptoclidus and related gen-era, while the cranial anatomy of rhomaleosau-rids has been treated recently by Cruickshank(1994a, 1994b, 1997). Carpenter (1996) re-viewed the cranial anatomy of the polycotylids.The primitive taxa Plesiosaurus (Storrs 1997)and Thalassiodracon (Storrs & Taylor 1996)have also been redescribed recently, as has theprimitive Eurycleidus (Cruickshank 1994b).
The sauropterygian skull roof is remarkablein several ways. The first and most obvious isthe pattern of temporal fenestration, where thepresence of the upper temporal fenestra and thelack of a lower temporal fenestra led Williston(1925) to classify the sauropterygians within his‘Synaptosauria’. Colbert amended this name to‘Euryapsida’, a term retained by Romer (1956,1966). Like many higher-level taxon names,‘Euryapsida’ has fallen into disuse due to a lackof certainty concerning its monophyly, althoughMerck (1997) has performed a cladistic analysison all ‘euryapsids’ and believes them to bemonophyletic.
The condition of the Permian plesiomorphicdiapsid Araeoscelis, described in detail byVaughn (1955), is an acceptable model fromwhich to derive the sauropterygian skull roof.All sauropterygians retain only the upper tempo-ral fenestra, as does Araeoscelis. The uppertemporal fenestra becomes greatly enlarged inthe Sauropterygia, concurrent with a narrowingof the parietal skull table, creating a large sur-face for attachment of the jaw adductor muscu-lature (Rieppel 1994a). The trends of enlarge-ment of the upper temporal fenestra and narrow-ing of the parietals become extreme in plesio-saurs; in all plesiosaurs, the posterior parietals

O’Keefe • ACTA ZOOL. FENNICA No. 2134
form a sharply keeled sagittal crest, and the pinealforamen moves far forward to a position near theparietal-frontal suture. Both of these charactersare present in Pistosaurus, the most plesiosaur-like of the basal sauropterygians. Also present inPistosaurus and all plesiosaurs is the ‘squamosalarch’, or the meeting at the postero-dorsal marginof the skull of median processes of the squamos-als. This median squamosal suture excludes theparietals from the occiput. The pistosaurid genusCymatosaurus is intermediate between basal‘nothosaurs’ and plesiosaurs in having a verynarrow exposure of the parietal on the occiput(Rieppel & Werneburg 1998).
In Araeoscelis, paired nasal bones contactthe external nares and meet in a long midlinesuture, anterior to the paired frontals that alsomeet broadly on the midline. All three clades ofsauropterygians contain members where the na-sal midline suture is lost due to the formation ofa midline frontal-premaxilla suture. Pistosaurushas this suture, and is further derived in havingvery small nasals that do not contact the externalnares (Edinger 1935). Rieppel (1994a) could notdetermine whether nasals were present in theremaining skull of this taxon. The presence orabsence of nasals in plesiosaurs has always beendebated, with Andrews (1910) identifying themin some pliosaurs. More recently, Storrs hasmaintained that all plesiosaurs lack nasals(Storrs 1991), and his skull roof reconstructionsof the very primitive plesiosaurs Plesiosaurus(Storrs 1997) and Thalassiodracon (Storrs &Taylor 1996) omit nasals. However, nasals arein fact present in Thalassiodracon as well as inall more derived members of the Pliosauroidea(see below). The nasal is lost in all Plesiosauroi-dea, including Plesiosaurus. When present inplesiosaurs, the nasals are always separated byposterior processes of the premaxillae, as is thecase in Cymatosaurus. In many plesiosaurs thepremaxillary processes extend back to contactthe parietals, separating the frontals on the skullsurface, although the frontals may continue tohave a midline suture beneath the premaxillae(Andrews 1911b).
Araeoscelis is plesiomorphic in the retentionof the lacrimal, stretching from orbit to externalnaris. The lacrimal is lost in all sauropterygians(Rieppel 1997b), including all plesiosaurs (con-
tra Andrews 1913, and Taylor & Cruickshank1993). The quadratojugal, also present in Araeo-scelis and most basal sauropterygians, is alsolost in the Pistosauroidea and all plesiosaurs.The position and relations of the jugal is varia-ble in sauropterygians, but Pistosaurus andmany plesiosaurs possess a prominent suturebetween the maxilla and squamosal. This sutureexcludes the jugal from the ventral skull margin;in Simosaurus and other ‘nothosaur’-grade sau-ropterygians the jugal enters the ventral skullmargin (Rieppel 1994a). The jugal is taxonomi-cally important in plesiosaurs, and the configu-ration of the other circum-orbital bones in gen-eral is also diagnostic.
The posterior palate and basicranium arediagnostic in plesiosaurs, but this area is highlyconserved in more basal sauropterygians, and isoften obscured by other skull elements. In all‘nothosaur’-grade sauropterygians, the ptery-goids meet in a median suture that extendscaudally almost to the occipital condyle, form-ing a solid plate of bone beneath the basicrani-um (Storrs 1991, Rieppel 1997b). This ptery-goid median suture closes off both the anteriorand posterior interpterygoid vacuities, the pres-ence of which in Araeoscelis exposes the dermalossification of the parasphenoid as well as theendochondral ossifications of the braincase (i.ebasisphenoid and basioccipital). Araeoscelis isprimitive in this regard, very similar to theCarboniferous diapsid Petrolacosaurus (Reisz1981). In Pistosaurus, the posterior interptery-goid vacuity reappears, again exposing the ven-tral surface of the braincase. The morphology ofthe braincase is unknown in Pistosaurus. Inprimitive plesiosaurs, the palate opens further toreveal the anterior parasphenoid and the brain-case. These structures show a remarkable simi-larity to the condition in Araeoscelis. The reap-pearance of the anterior and posterior interptery-goid vacuities in plesiosaurs is correlated with atrend of reduced ossification in the skeletongenerally, a common feature of many aquatictetrapods (Storrs 1991, Romer 1956). Ossifica-tion of the more ventral structures of the brain-case is also reduced in plesiosaurs, to the pointthat the dorsum sellae and sella turcica are nolonger identifiable in derived pliosauroids.These structures are prominent and well-ossified

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 5
in Nothosaurus (Rieppel 1994b), and are presentbut less well-ossified in the pistosauroid Cyma-tosaurus (Rieppel & Werneburg 1998). Cymato-saurus also possesses a deep, narrow notch inthe posterior border of the clivus, a feature itshares with all plesiosaurs in which the dorsumsellae and clivus are ossified (Fig. 1).
Postcranial anatomy
Romer’s (1956: pp. 298–332) introduction tothe anatomy of the reptilian limb girdles illus-trates the general conditions of the pectoral andpelvic girdles in primitive amniotes. Again,Araeoscelis is a plesiomorphic diapsid that maybe used as a model from which to derive thesauropterygian conditions. In the pelvic girdle ofAraeoscelis, the pubis and ischium form a solidplate without a thyroid fenestra, surmounteddorsally by the ilium (Vaughn 1955). The obtu-rator foramen is primitively present in the pubis.All sauropterygians have a large thyroid fe-nestra. The obturator foramen is present in allsauropterygians except plesiosaurs, the pistosau-roid Cymatosaurus, and the true nothosaur Lari-osaurus. Girdle ossification in plesiosaurs isslow and varies intra-specifically to some degree(Brown 1981), leading Brown to question thetaxonomic utility of characters relating to girdleshape. The plesiosaur pelvic girdle is apomor-phic in the great reduction of the ilium, the lossof contact between ilium and pubis, and theconcomitant absence of the ilium from theacetabulum. In all plesiosaurs, the ilium is anarrow rod of bone running from the ischium to
a reduced sacrum composed of one to threesacral ribs.
The pectoral girdle is more complex andmuch more variable within the Sauropterygia.All sauropterygians have one coracoid ossifica-tion rather than the primitive two present inAraeoscelis; the cleithrum is also absent in allsauropterygians (Rieppel 1997b). Furthermore,there is a broad trend of reduction in the dorsalstructures of the pectoral girdle and elaborationof them ventrally. In placodonts the shouldergirdle is still relatively plesiomorphic. The bladeof the scapula is prominent laterally, and thecoracoids do not meet on the ventral midline.However, the dermal elements of the shouldergirdle (clavicles and interclavicle) are large androbust, and are primarily ventral structures. In‘nothosaur’-grade sauropterygians, the claviclesremain large and, unlike placodonts, have aprominent medial suture and concomitant reduc-tion of the interclavicle. Also in contrast toplacodonts, ‘nothosaur’-grade sauropterygianspossess a ventral median suture of the coracoids.The coracoids are enlarged relative to those ofplacodonts and are distinctively narrowed or‘waisted’ near their centers (Storrs 1991: p. 75).The coracoids are plate-like and not waisted inthe pistosauroid Corosaurus. In Augustasaurusthe coracoids are large ventral plates similar tothose in plesiosaurs (Sander et al. 1997; author’spers. obs.). The coracoid foramen is also lost inCorosaurus and Augustasaurus, a trait thesetaxa share with plesiosaurs.
The pectoral girdle in plesiosaurs is charac-terized by reduction of the scapular blade(Romer 1956) and a great ventral elaboration of
Fig. 1. Eurycleidus arcu-atus braincase, OUMJ.28585. Length of frag-ment 4.5 cm.

O’Keefe • ACTA ZOOL. FENNICA No. 2136
the scapulae. The dorsal blade of the scapula issituated dorsal to the glenoid in ‘nothosaur’-grade sauropterygians, but is anterior to theglenoid in all plesiosaurs (Kebang & Rieppel1998). The ventral processes of the scapulaeangle anteriorly toward the midline and meet ina midline suture in some derived taxa. Thisventral elaboration of the scapulae comes at theexpense of the clavicles and interclavicle, whichare reduced and near the midline in plesiosaurs.The coracoids are also greatly elaborated inplesiosaurs, forming large plate-like extensionsposteriorly with a long midline suture betweenthem. The coracoids may also send processesforward to meet either the clavicles or scapulaeon the midline, dividing the ventral space en-closed by the pectoral girdle into two pectoralfenestrae (for example Welles 1962, Storrs1997). This median pectoral bar is slow to ossify(Brown 1981), as is the comparable medianpelvic bar (present in Plesiosaurus, Storrs 1997,and some elasmosaurs, Welles 1962). The ag-gregate effect of changes in the pelvic andpectoral girdles is the formation of two large,
ventral plates of bone vaguely reminiscent of aturtle plastron. The short space between theanterior and posterior girdles was filled withclosely spaced gastralia. The ventral plates werepresumably the areas of attachment for largelocomotor muscles (Williston 1914).
The limbs of plesiosaurs are highly specializedhydrofoils. The modifications of propodials andepipodials are complex in sauropterygians, espe-cially across the pistosaurid/plesiosaur transitionand within basal plesiosaurs. In general terms, thepropodials tend to lose obvious processes for mus-cle attachment and become short and stout (Storrs1997, Romer 1956). A pronounced bend caudad inthe shaft of the humerus is characteristic of ‘notho-saur’-grade taxa and some very primitive plesio-saurs, but the shaft is straight in all derived plesio-saurs (Storrs 1997). Both the ectepicondylar andentepicondylar foramina are lost in all plesiosaurs;in ‘nothosaur’-grade sauropterygians the ectepi-condylar foramen is often reduced to a groove ornotch, and both features are variably present inbasal sauropterygians. However, the pistosauroidsCorosaurus and Cymatosaurus possess both open-
Table 1. Repositories and abbreviations for sauropterygian material referenced in this report.————————————————————————————————————————————————
Repository Abbrev. Location————————————————————————————————————————————————01 American Museum of Natural History AMNH New York, New York, USA02 The Natural History Museum BMNH London, UK03 Banque de la República de Villa de Leyva BRI Bogota, Columbia04 Bristol Museum and Art Gallery BRSMG Bristol, UK05 Sedgwick Museum CAMSM Cambridge, UK06 Sternberg Museum of Natural History FHSM Fort Hayes, Kansas, USA07 Field Museum of Natural History FMNH Chicago, Illinois, USA08 Institut und Museum für Geologie und Paläontologie GPIT Tübingen, Germany09 Urwelt Museum Hauff Hauff Holzmaden, Germany10 Kansas Museum of Natural History KUVP Lawrence, Kansas, USA11 Leicester City Museum LEICT Leicester, UK12 Manchester Museum MAN UM Manchester, UK13 Geologisch-Paläontologisches Museum der Universität Münster Münster Münster, Germany14 Museum of Comparative Zoology, Harvard University MCZ Cambridge, Massachusetts, USA15 Museum of the Rockies MOR Bozeman, Montana, USA16 National Museum of Wales NMW Cardiff, Wales17 Oxford University Museum of Natural History OXFUM Oxford, UK18 Peterborough Museum and Art Gallery PETMG Peterborough, UK19 Strecker Museum, Baylor University SM Waco, Texas, USA20 Staatliches Museum für Naturkunde, Stuttgart SMNS Stuttgart, Germany21 Southern Methodist University Museum of Paleontology SMUSMP Dallas, Texas, USA22 Texas Technical Institute TTVP Lubbock, Texas, USA23 University of California Museum of Paleontology UCMP Berkeley, California, USA24 Smithsonian Institution USNM Washington, D.C., USA25 University of Wyoming UW Laramie, Wyoming, USA26 Yorkshire Museum YORYM York, UK27 Yale-Peabody Museum YPM New Haven, Connecticut, USA————————————————————————————————————————————————
ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 7
ings, whereas the pistosaurids Pistosaurus andAugustasaurus lack both openings, another condi-tion these taxa share with plesiosaurs. The epipodi-als resemble the propodials in becoming relativelyshort and broad in basal eosauropterygians. Themanus and pes are characterized by hyperphalangyin all plesiosaurs. The phalangeal formulas in thepistosaurids are unknown. The hypophalangy hy-pothesized by Sander et al. (1997) in Augustasau-rus is probably an artifact of preservation (O.Rieppel pers. com.).
Materials and methods
The plesiosaur material discussed here is held in27 institutions in the United States, South Ameri-ca, and Europe (listed in Table 1). Material in 24of these institutions was examined personally;material from the other three institutions (Geolo-gisch-Paläontologisches Museum der UniversitätMünster, Museum of the Rockies, Banque de laRepública de Villa de Leyva) was scored fromthe literature. The three outgroup and 31 ingrouptaxa considered in this report are listed below.Each genus is presented with primary and morerecent references, holotype and specimen number,referred material, and other remarks. Charactersused in the phylogenetic analysis are discussed inthe second part of the Materials and Methodssection, and listed in Appendix 1. The data matrixis listed in Appendix 2.
Description of Hauffiosaurus gen. n. andHauffiosaurus zanoni sp. n.
Hauffiosaurus gen. n.
DIAGNOSIS: as for species.
ETYMOLOGY: The genus name refers to the Urwelt-Muse-um Hauff, the institution in Holzmaden, Germany wherethe type species is housed.TYPE SPECIES: Hauffiosaurus zanoni
Hauffiosaurus zanoni sp. n.
DIAGNOSIS: A plesiomorphic member of cladePliosauridae possessing the following autapo-
morphies: seven premaxillary teeth; ectoptery-goid boss contacts jugal exclusively; propodialsvery long.
TYPE MATERIAL: — Holotype: Hauff museum uncata-logued, a complete skeleton in the Hauff Museum, Holz-maden, Germany. Figs. 2 and 3.ETYMOLOGY: The species name zanoni refers to the late R.T. Zanon, who first realized this specimen represented anew genus (R. T. Zanon unpubl.).REFERRED MATERIAL: none.TYPE LOCALITY: Posidonien-Schiefer, Holzmaden, Germany.AGE: Toarcian.
DESCRIPTION: The specimen here designatedHauffiosaurus zanoni (Hauff museum uncata-logued) is a complete plesiosaur skeleton pre-served in ventral view, found in the Posidonien-Schiefer of Holzmaden, Germany (Toarcian).The skeleton is approximately 2.5 m long, anddisplays an interesting mix of plesiomorphic,derived, and apomorphic features. For a com-plete listing of character states in this taxon, seeAppendix 2.
The skull is preserved in palatal view(Figs. 2 and 3), with the mandibles crushed anddisplaced to one side. The snout and mandibular
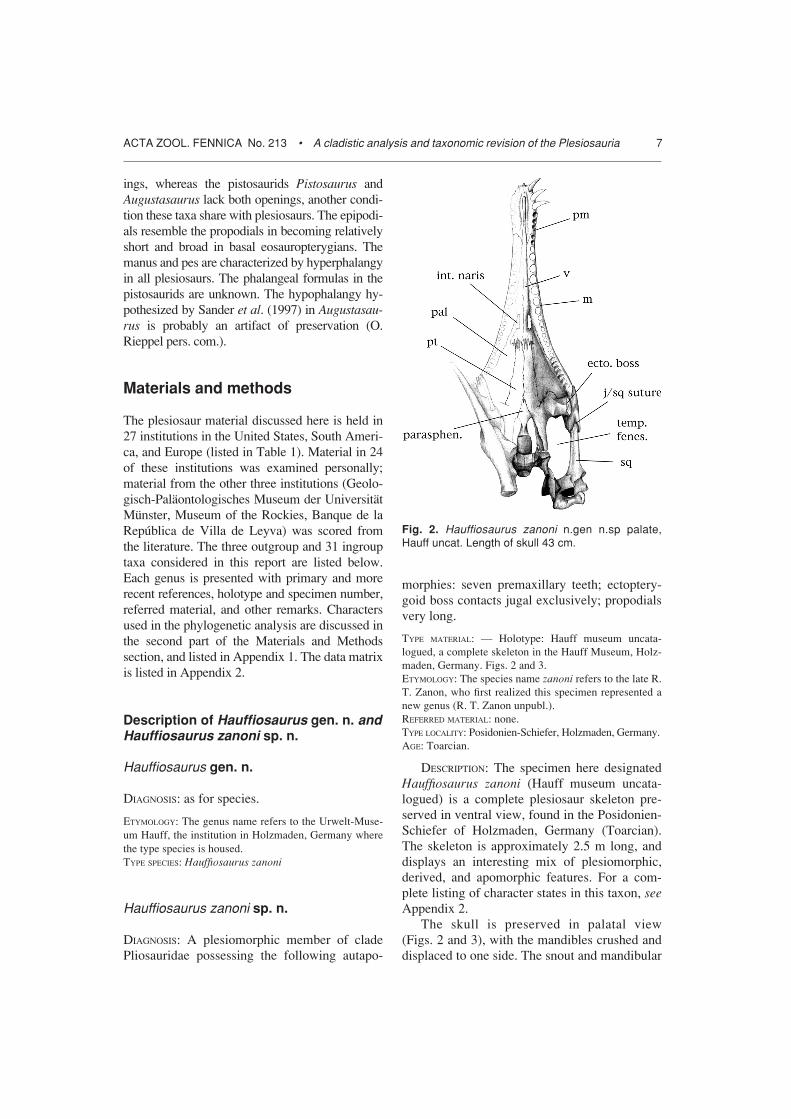
Fig. 2. Hauffiosaurus zanoni n.gen n.sp palate,Hauff uncat. Length of skull 43 cm.

O’Keefe • ACTA ZOOL. FENNICA No. 2138
symphysis are both very long, indicating that thespecimen is not a rhomaleosaur, and is instead amember of the Pliosauridae. On the palate, thevomers extend beyond the posterior margin ofthe internal nares and meet the pterygoids in awide, interdigitating suture very similar to thatin Peloneustes. This suture lacks the medianextension on the midline characteristic of Mac-roplata. The Hauff taxon also possess a promi-nent ectopterygoid boss unlike Macroplata butlike Peloneustes, although the boss articulateslaterally with the jugal only rather than both thejugal and squamosal as in Peloneustes. There isno anterior interpterygoid vacuity. The paras-phenoid is exposed prominently on the palate
surface anterior to the posterior interpterygoidvacuity. The basisphenoid is poorly ossified, asin Peloneustes and all other pliosaurids exceptMacroplata. The pterygoids meet behind theposterior interpterygoid vacuity, and there isevidence of a lateral flange around the vacuitiessimilar to that in Peloneustes and Kronosaurus.However, the sigmoid shape and rolled lateralmargin of the quadrate flange of the pterygoidare absent in the Hauff taxon, as in Macroplatabut unlike all more derived pliosaurids. Thereare seven teeth in each premaxilla, an autapo-morphy (Peloneustes has six teeth in the pre-maxilla). In sum, the palate of the Hauff speci-men shows a mixture of primitive and derived
Fig. 3. Photographs ofthe holotype of Hauffio-saurus zanoni. Top, de-tail of skull and mandi-bles. Compare to Fig. 2.Length of skull 43 cm.Bottom, entire skeletonas preserved. Note that‘body outline’ was carvedduring preparation; nosoft tissue is preservedwith the specimen.

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 9
features, and seems to be intermediate betweenthe plesiomorphic Macroplata and more derivedPeloneustes. The position of the Hauff taxon onthe cladogram reflects this impression.
The most remarkable aspect of the postcrani-um is the extreme length of the propodials. Thehumerus is 32 cm long, while the femur is 35 cmlong. Both propodials are longer than any oftheir associated girdle bones, a unique featureamong plesiosaurs and an autapomorphy of thistaxon. The postcranium in general displays rela-tively small girdles, high aspect-ratio fins, and aneck that is primitive in length and vertebralnumber. Hauffiosaurus zanoni is a stratigraphi-cally early (Toarcian) and morphologically inter-mediate member of the Pliosauridae, and pos-sesses several autapomorphies in the skull andpostcranium.
Included taxa
Outgroup taxa
Three taxa were chosen as outgroups for thisanalysis. They are the nothosaur Simosaurus, thepistosauroid Cymatosaurus, and the family Pis-tosauridae. The family Pistosauridae comprisesthe genera Pistosaurus and Augustasaurus.These two taxa were scored together becausethey are closely related, and because the materialfor the two genera is complementary. The postc-ranium of Augustasaurus is articulated and well-preserved, unlike that of Pistosaurus, which hasled to some doubt in the postcranial reconstruc-tion of the later taxon (Sues 1987). The skull ofAugustasaurus is extant, and a description is inpress a this time (O. Rieppel pers. com.), whilethat of Pistosaurus was described in detail byEdinger (1935; skull now lost).
GENUS: Simosaurus Meyer, 1842
TYPE SPECIES: Simosaurus gaillardoti Meyer, 1842.TYPE MATERIAL: — Holotype: lost. — Neotype: SMNS16700 (Rieppel 1994a).TYPE LOCALITY: Upper Muschelkalk, Lunéville, France.AGE: Ladinian.REFERRED MATERIAL: various, one known species.REMARKS: Taxonomy and referred material reviewed inRieppel 1994a.
GENUS: Cymatosaurus v. Fritsch, 1894
TYPE SPECIES: Cymatosaurus fridericianus v. Fritsch, 1894.TYPE MATERIAL: — Holotype: Institut für Geowissen-schaften, Martin-Luther Universität, Halle/Saale, uncata-logued.TYPE LOCALITY: Lower Muschelkalk, Halle/Saale, Germany.AGE: Ladinian.REFERRED MATERIAL: various, at least four known species.REMARKS: Taxonomy and referred material reviewed inRieppel (1997b, 1998b), and Rieppel and Hagdorn (1997).
FAMILY: Pistosauridae Zittel, 1887
TYPE SPECIES: Pistosaurus longaevus Meyer, 1839.TYPE MATERIAL: — Holotype: Oberfränkisches Erdges-chichtliches Museum, Bayreuth, uncatalogued.TYPE LOCALITY: Upper Muschelkalk, Bayreuth, Germany.AGE: Late Anisian.REFERRED MATERIAL: various, possibly two species.REMARKS: Taxonomy, referred material, and postcraniumreviewed in Sues (1987). Cranium described by Edinger(1935), and Rieppel (1994a).
TYPE SPECIES: Augustasaurus hagdorni Sander, Rieppeland Bucher, 1997.TYPE MATERIAL: — Holotype: FMNH PR 1974.TYPE LOCALITY: Favret Formation, Pershing County, Nevada.AGE: Late Anisian.REFERRED MATERIAL: none.REMARKS: Augustasaurus is one of two nothosaur-gradesauropterygians from North America; the other is Coro-saurus. The skull of Augustasaurus is currently beingdescribed by O. Rieppel, M. Sander, and G. Storrs.
Ingroup taxa
Thirty-one plesiosaur genera were coded forinclusion in the phylogenetic data matrix. Twoof these taxa are unnamed at present; these taxaare referred to by specimen number. Several ofthe genera included here contain more than onespecies. The common genus Plesiosaurus prob-ably contains three valid species, while Mu-raenosaurus contains at least two valid species.Because genus-level and higher relationshipsare the primary goal of this analysis, polytypicgenera are treated as terminal taxa here. Howev-er, species level diagnoses are given in theSystematic Paleontology section where appro-priate. Another complication is that the genusRhomaleosaurus is paraphyletic (see Results).The species of Rhomaleosaurus included here

O’Keefe • ACTA ZOOL. FENNICA No. 21310
are referenced using both genus and speciesnames. The listings of referred material belowmention only material used in this study; com-plete lists of referred material can generally befound in references cited in the remarks for eachtaxon. The sampling of plesiosaur taxa in thisstudy is not exhaustive. Several Rhomaleosau-rus species are omitted, as are several elasmo-saur genera and some other poorly-known taxa.All known clades are well-represented, howev-er, and the omission of some ingroup taxa fromsome clades should not influence the resultsreported here. The genus Rhomaleosaurus andthe Elasmosauridae are both in need of taxo-nomic revision. Lastly, exhaustive synonomiesare not given for each taxon. Some remarks on
the taxonomic history of most genera are givenin the remarks section, along with referenceswhere synonomies can be found for those taxa.
GENUS: Thalassiodracon Storrs & Taylor, 1996
TYPE SPECIES: Thalassiodracon hawkinsi (Owen, 1838).TYPE MATERIAL: — Holotype: BMNH 2018.TYPE LOCALITY: Street, Somerset, England.AGE: Rhaetian-Hettangian boundary.REFERRED MATERIAL: BMNH 14550, 2020, R.1336 (juv.)CAMSM J.46986, OUM J.10337, MM L. 9767.REMARKS: Fig. 4. Storrs and Taylor (1996) created thisgenus name for the taxon ‘Plesiosaurus’ hawkinsi (Owen1838), a stratigraphically early and morphologically prim-itive plesiosaur. These authors list other material referableto this taxon, and describe the skull in some detail. Aredescription of the postcranium is necessary.
GENUS: Eurycleidus Andrews, 1922
TYPE SPECIES: Eurycleidus arcuatus (Owen, 1840).TYPE MATERIAL: — Holotype: BMNH 2030 (see Cruick-shank 1994b).TYPE LOCALITY: Lyme Regis, Dorset, England.AGE: Hettangian- Lower Sinemurian.REFERRED MATERIAL: OUM J.28585, SMNS 16812.REMARKS: Fig. 1. Cruickshank (1994b) redescribes this
Fig. 4. Thalassiodracon hawkinsi skull roof, OUMJ.10337. Abbreviations used in the following figuresare as follows: aipv, anterior interpterygoid vacuity;ang, angular; cor, coranoid; d, dentary; ect, ectop-terygoid; f, frontal; j, jugal; m, maxilla; n, nasal; p,parietal; pal, palatine; pf, prefrontal; pipv, posteriorinterpterygoid vacuity; pm, premaxilla; po, postor-bital; pof, postfrontal; preart, prearticular; pt, ptery-goid; q, quadrate; sof, suborbital fenestra; spl, sple-nial; sq, squamosal; surang, surangular; v, vomer.Length of skull 18 cm.
Fig. 5. Plesiosaurus dolichodeirus skull roof, BMNH39490. Length of skull 23 cm.

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 11
genus and refers other material to it. I refer the Germanspecimen to this taxon based on similarities in the basic-ranium; however this referral is provisional until theGerman material (a complete skeleton) can be describedin more detail. Fig. 1.
GENUS: Plesiosaurus De la Beche & Conybeare,1821
TYPE SPECIES: Plesiosaurus dolichodeirus, Conybeare 1824.TYPE MATERIAL: — Holotype: BMNH 22656.TYPE LOCALITY: Lyme Regis, Dorset, England.AGE: Uppermost Sinemurian.REFERRED MATERIAL: P. dolichodeirus: BMNH 39490,41101, 36183, R.1756, OXFUM J.10304; P. brachyp-terygius: SMNS 51143, Hauff uncatalogued, GPIT 477/1/1; P. guiliemiimperatoris: SMNS 51015.REMARKS: Figs. 5 and 6. Andrews (1896) gives the firstdetailed account of the structure of the palate in this taxon.The genus is reviewed exhaustively in Storrs (1997).Storrs discusses other material also referable to this taxon,and advances the hypothesis that the many nominal speciesin this common and widespread genus are representative ofperhaps three valid species. My interpretation differssomewhat; P. brachypterygius lacks the lunate ulna char-acteristic of P. dolichodeirus, and may be a valid species
as well. I agree with Storrs that P. guilemiimperatoris is avalid species; that author also states that material fromFrance may represent another valid species. The Frenchmaterial was not examined in the course of this study.
GENUS: Rhomaleosaurus Seeley, 1874 (Seeley1874b)
TYPE SPECIES: Rhomaleosaurus cramptoni Seeley, 1874.
Rhomaleosaurus victor (Fraas, 1910)
TYPE MATERIAL: — Holotype: SMNS 12478.TYPE LOCALITY: Posidonien-Schiefer, Holzmaden, Germany.AGE: Toarcian.REFERRED MATERIAL: none.REMARKS: Fig. 7. The taxonomic history of this species iscomplex; this species was originally described as ‘Thau-matosaurus victor’ (Meyer 1841) by Fraas (1910). Thegenus Rhomaleosaurus is in need of revision (Cruick-shank 1994a; see below). Lydekker (1889b) attempted tomake Rhomaleosaurus a junior synonym of Thaumato-saurus, based on his opinion that Seeley’s type of thegenus, Rhomaleosaurus cramptoni (Seeley 1874b), doesnot differ from the type material of ‘Thaumatosaurus’.Tarlo (1960) rejected ‘Thaumatosaurus’ because the typematerial is not diagnostic (reviewed in Taylor 1992a).
Fig. 6. Plesiosaurus dolichodeirus palate, BMNH41101. Length of skull 20 cm.
Fig. 7. Rhomaleosaurus victor palate, SMNS 12478.Length of mandible 40 cm.

O’Keefe • ACTA ZOOL. FENNICA No. 21312
Tarlo (1960) also believed the complete skeleton named‘Thaumatosaurus’ victor by Fraas (1910) to be congener-ic with Rhomaleosaurus, and recommended dropping‘Thaumatosaurus’ altogether. I follow his suggestion.Lydekker (1889a) also referred an isolated jaw symphy-sis from India to Thaumatosaurus; the symphysis hascharacters diagnostic of the Rhomaleosauridae but is notdiagnostic at the genus level.
Rhomaleosaurus megacephalus Stuchbury, 1846
TYPE MATERIAL: — Holotype: lost. — Neotype: LEICTG221.1851 (Cruickshank 1994a).TYPE LOCALITY: Barrow upon Soar, Leicestershire, England.AGE: Earliest Hettangian.REMARKS: Fig. 8. Cruickshank (1994a) designates anddescribes the neotype, which is a complete skeletonsuperficially similar to the destroyed holotype describedand figured by Stuchbury (1846) under the name ‘Plesio-saurus’ megacephalus. Cruickshank (1994a) and Taylor(1992a) also discuss the taxonomic confusion surround-ing the name Rhomaleosaurus.
Rhomaleosaurus zetlandicus Phillips, 1854 (inAnon. 1854)
TYPE MATERIAL: — Holotype: YORYM G503TYPE LOCALITY: Alum Shale, Whitby, Yorkshire, England.
AGE: Toarcian.REMARKS: Taylor (1992a) discusses the taxonomy of thisspecimen and of Rhomaleosaurus in general. Taylor(1992b) describes the cranial anatomy of this taxon.
GENUS: Simolestes Andrews, 1909
TYPE SPECIES: Simolestes vorax Andrews 1909.TYPE MATERIAL: — Holotype: BMNH R.3319.TYPE LOCALITY: Oxford Clay, Peterborough, England.AGE: Callovian.REFERRED MATERIAL: none.REMARKS: Andrews (1913) and Tarlo (1960) describe thistaxon thoroughly.
GENUS: Leptocleidus Andrews, 1922
TYPE SPECIES: Leptocleidus capensis (Andrews, 1911a).TYPE MATERIAL: — Holotype: BMNH R.4828.TYPE LOCALITY: Berwick, Sussex, England.AGE: Barremian.REFERRED MATERIAL: SAM-K5822 (South African Museum).REMARKS: Cruickshank (1997) reviews the type materialof this genus and refers the South African specimen tothe genus. The type material was originally described byAndrews (1911a) under the genus name ‘Plesiosaurus’.
GENUS: Macroplata Swinton, 1930
TYPE SPECIES: Macroplata longirostris (Blake, 1876).TYPE MATERIAL: — Holotype: MCZ 1033.TYPE LOCALITY: Alum Shale, Whitby, Yorkshire, England.AGE: Toarcian.REFERRED MATERIAL: MAN UM 8004.REMARKS: Fig. 9. Macroplata longirostris was named anddescribed from a complete skeleton by Blake (1876) as‘Plesiosaurus’ longirostris. Swinton (1930) erected Mac-roplata for the specimen when he realized that it was notreferable to Plesiosaurus. However, as discussed byWhite (1940), Blake and later workers were aware thatthe specimen was a composite. Blake (1876) argued thatthe holotype be restricted to the skull only. White (1940)felt that the associated vertebral column might also beincluded in the holotype. The skull was prepared poorlyin Victorian times, and the serious ‘pyrite disease’ notedby White (1940) has continued to degrade the alreadybattered skull. However, several diagnostic characters arestill visible, such as the very long snout and the plesio-morphic, rhomaleosaur-like pattern of the posterior ba-sicranium. Both of these characters are shared by a newskull with associated complete skeleton in the Manches-ter Museum (MAN UM 8004), which was found at thesame locality and in the same formation as the holotype.The Manchester specimen also possesses a groove infront of the external naris, and a perforation in the dorsalsurface of the basisphenoid through which the clivus isvisible. Both of these characters are also present in theMacroplata holotype, and I therefore refer MAN UM8004 to this taxon.
Fig. 8. Rhomaleosaurus megacephalus palate, LEICTG221.1851. Length of mandible 42 cm.

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 13
GENUS: unnamed (‘Macroplata tenuiceps’).
TYPE SPECIES: n/a.TYPE MATERIAL: — Holotype: BMNH R.5488.TYPE LOCALITY: Harbury, Warwickshire, England.AGE: unknown.REMARKS: This specimen is a complete skeleton in thecollection of the Natural History Museum, London. Thespecimen is identified as ‘Macroplata tenuiceps’ on thelabels accompanying the material. The skeleton is cer-tainly not referable to this genus based on charactersnoted by Cruickshank (1994a), Swinton (1930), andWhite (1940). The specimen is certainly a rhomaleosau-rid, although the skull material is not diagnostic at thespecies level. The taxonomic status of this specimenawaits revision of the Rhomaleosauridae.
GENUS: Hauffiosaurus gen. n.
TYPE SPECIES: Hauffiosaurus zanoni sp. n.HOLOTYPE : Hauff Museum, uncatalogued; see above.TYPE LOCALITY: Posidonien-Schiefer, Holzmaden, Germany.AGE: Toarcian.REFERRED MATERIAL: none.REMARKS: Figs. 2 and 3. This specimen is a complete
skeleton of an unnamed taxon in the collections of theHauff Museum. The taxon is named here, and a prelimi-nary description is given above with figures of the skulland skeleton.
GENUS: Peloneustes Lydekker, 1889 (Lydekker1889b)
TYPE SPECIES: Peloneustes philarchus (Seeley, 1869).TYPE MATERIAL: — Holotype: CAMSM J.46913TYPE LOCALITY: Oxford Clay, Peterborough, England.AGE: Callovian.REFERRED MATERIAL: BMNH R.8574, R.3803, R.3897,R.3318.REMARKS: Figs. 10, 11 and 12. Lydekker names anddescribes this taxon (1889b), and comments on itsrelationships to Thaumatosaurus. Tarlo (1960) discuss-es this taxon, and Andrews (1913) describes it in detail.Linder (1913) describes Oxford Clay material of thistaxon at the SMNS, and compares it to Pliosaurus. Thistaxon is extremely well-represented by material in theLeeds collection in the Natural History Museum
Fig. 9. Macroplata longirostris palate, MAN UM8004.
Fig. 10. Peloneustes philarchus skull roof, BMNH R.8574. Length of skull 69.5 cm.

O’Keefe • ACTA ZOOL. FENNICA No. 21314
(BMNH), including several complete skeletons withwell-preserved skulls.
GENUS: Liopleurodon Sauvage, 1873
TYPE SPECIES: Liopleurodon ferox Sauvage, 1873.TYPE MATERIAL: — Holotype: BMNH R.3536.TYPE LOCALITY: Oxford Clay, Peterborough, England.AGE: Callovian.REFERRED MATERIAL: BMNH R.2680, GPIT 1754/2.REMARKS: Tarlo (1960) offered the latest review of UpperJurassic pliosaurs, and reviews the complex taxonomichistory of the genus name. Andrews (1913) describes thistaxon in detail under the name ‘Pliosaurus’ ferox. Thegenus Stretosaurus was a made a junior synonym of thistaxon by Halstead (1989); this poorly-defined taxon isbased on enormous pliosaurid postcranial elements fromthe Kimmeridge Clay. Further comparison of this materi-al with that of Pliosaurus is probably necessary.
GENUS: Pliosaurus Owen, 1841
TYPE SPECIES: Pliosaurus brachydeirus Owen, 1841.
TYPE MATERIAL: — Holotype: OXFUM J.9245 A,B.TYPE LOCALITY: Oxford Clay, Peterborough, England.AGE: Callovian.REFERRED MATERIAL: BMNH R.3891, BRSMG Cc332,‘Westbury Pliosaur 2’ (BRSMG uncatalogued.).REMARKS: Taylor and Cruickshank (1993) discuss thetaxonomic history of this genus. Tarlo (1960) also reviewsthis taxon. Referred specimens listed above are Kim-meridgean, from the Kimmeridge Clay in Wiltshire, UK.
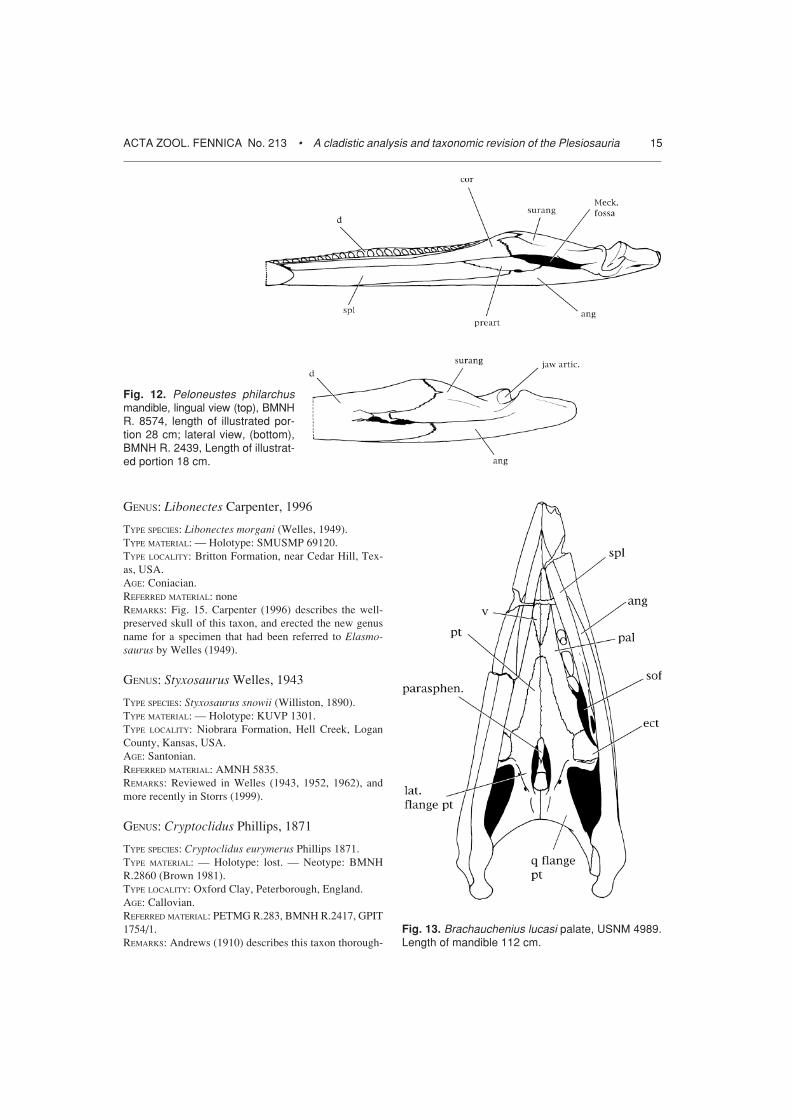
GENUS: Brachauchenius Williston, 1903
TYPE SPECIES: Brachauchenius lucasi Williston, 1903.TYPE MATERIAL: — Holotype: USNM 4989.TYPE LOCALITY: Greenhorn Limestone, Ottawa County,Kansas, USA.AGE: Turonian.REFERRED MATERIAL: USNM 2361, FHSM VP321.REMARKS: Fig. 13. This taxon was described in detail byWilliston (1907). Carpenter (1996) referred the FortHayes skull to the genus.
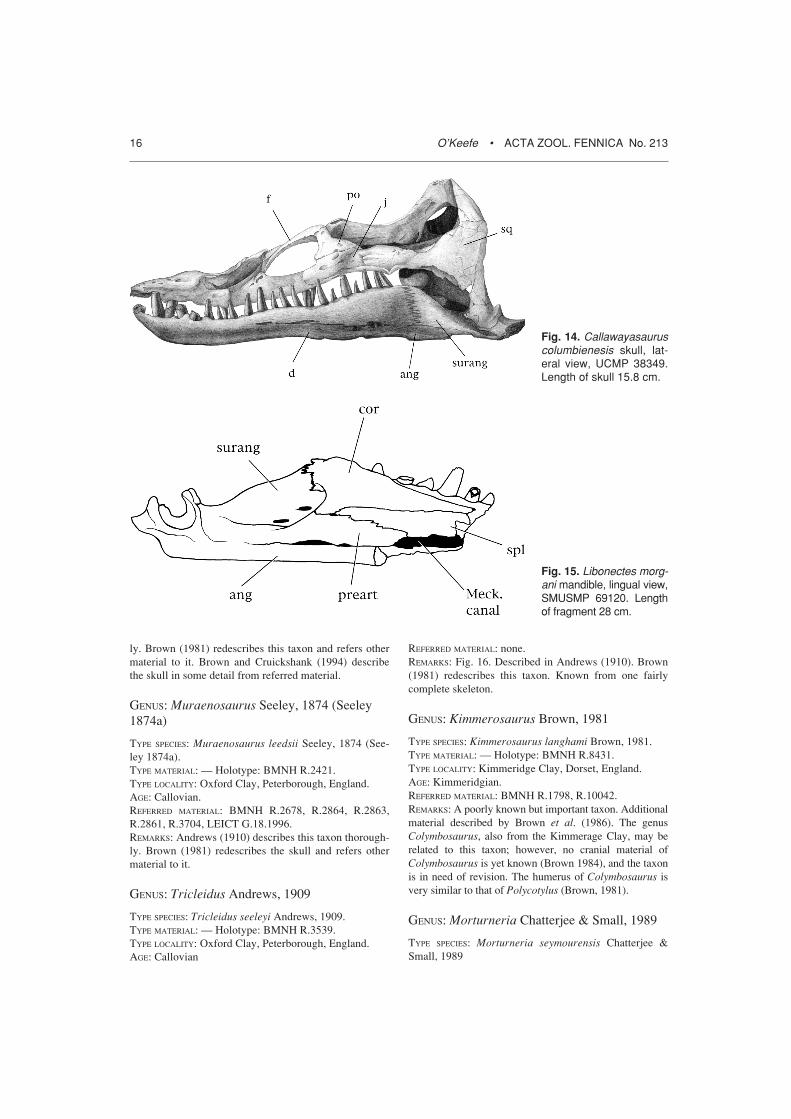
GENUS: Microcleidus Owen, 1865
TYPE SPECIES: Microcleidus homalospondylus Owen, 1865.TYPE MATERIAL: — Holotype: YORYM G.502.TYPE LOCALITY: Alum Shale, Whitby, England.AGE: Toarcian.REFERRED MATERIAL: MM L. 7077.REMARKS: This genus is in need of redescription, havinglast been treated by Watson (1911).
GENUS: Brancasaurus Wegner, 1914
TYPE SPECIES: Brancasaurus brancai Wegner 1914.TYPE MATERIAL: — Holotype: Münster; Wegner does notgive the specimen number.TYPE LOCALITY: Münster, Westphalia, Germany.AGE: Valangian.REFERRED MATERIAL: none.REMARKS: Wegner’s (1914) thorough description is theonly publication on this taxon.
GENUS: Callawayasaurus Carpenter, 1999
TYPE SPECIES: Callawayasaurus columbienesis (Welles,1962).TYPE MATERIAL: — Holotype: UCMP 38349.TYPE LOCALITY: Columbia, South America.AGE: Aptian.REFERRED MATERIAL: UCMP 125328, partial skull. Samelocality.REMARKS: Fig. 14. This genus was erected by Carpenter(1999) for the holotype of ‘Alzadasaurus’ columbienesis(Welles 1962), an essentially complete elasmosaur skele-ton from the Aptian of Columbia. The skull shows anumber of primitive features. For holotype and otherinformation on Alzadasaurus proper see Welles (1962).
Fig. 11. Peloneustes philarchus palate, SMNS 10113.Length of skull 63 cm.
ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 15
GENUS: Libonectes Carpenter, 1996
TYPE SPECIES: Libonectes morgani (Welles, 1949).TYPE MATERIAL: — Holotype: SMUSMP 69120.TYPE LOCALITY: Britton Formation, near Cedar Hill, Tex-as, USA.AGE: Coniacian.REFERRED MATERIAL: noneREMARKS: Fig. 15. Carpenter (1996) describes the well-preserved skull of this taxon, and erected the new genusname for a specimen that had been referred to Elasmo-saurus by Welles (1949).
GENUS: Styxosaurus Welles, 1943
TYPE SPECIES: Styxosaurus snowii (Williston, 1890).TYPE MATERIAL: — Holotype: KUVP 1301.TYPE LOCALITY: Niobrara Formation, Hell Creek, LoganCounty, Kansas, USA.AGE: Santonian.REFERRED MATERIAL: AMNH 5835.REMARKS: Reviewed in Welles (1943, 1952, 1962), andmore recently in Storrs (1999).
GENUS: Cryptoclidus Phillips, 1871
TYPE SPECIES: Cryptoclidus eurymerus Phillips 1871.TYPE MATERIAL: — Holotype: lost. — Neotype: BMNHR.2860 (Brown 1981).TYPE LOCALITY: Oxford Clay, Peterborough, England.AGE: Callovian.REFERRED MATERIAL: PETMG R.283, BMNH R.2417, GPIT1754/1.REMARKS: Andrews (1910) describes this taxon thorough-
Fig. 12. Peloneustes philarchusmandible, lingual view (top), BMNHR. 8574, length of illustrated por-tion 28 cm; lateral view, (bottom),BMNH R. 2439, Length of illustrat-ed portion 18 cm.
Fig. 13. Brachauchenius lucasi palate, USNM 4989.Length of mandible 112 cm.
O’Keefe • ACTA ZOOL. FENNICA No. 21316
ly. Brown (1981) redescribes this taxon and refers othermaterial to it. Brown and Cruickshank (1994) describethe skull in some detail from referred material.
GENUS: Muraenosaurus Seeley, 1874 (Seeley1874a)
TYPE SPECIES: Muraenosaurus leedsii Seeley, 1874 (See-ley 1874a).TYPE MATERIAL: — Holotype: BMNH R.2421.TYPE LOCALITY: Oxford Clay, Peterborough, England.AGE: Callovian.REFERRED MATERIAL: BMNH R.2678, R.2864, R.2863,R.2861, R.3704, LEICT G.18.1996.REMARKS: Andrews (1910) describes this taxon thorough-ly. Brown (1981) redescribes the skull and refers othermaterial to it.
GENUS: Tricleidus Andrews, 1909
TYPE SPECIES: Tricleidus seeleyi Andrews, 1909.TYPE MATERIAL: — Holotype: BMNH R.3539.TYPE LOCALITY: Oxford Clay, Peterborough, England.AGE: Callovian
Fig. 14. Callawayasauruscolumbienesis skull, lat-eral view, UCMP 38349.Length of skull 15.8 cm.
Fig. 15. Libonectes morg-ani mandible, lingual view,SMUSMP 69120. Lengthof fragment 28 cm.
REFERRED MATERIAL: none.REMARKS: Fig. 16. Described in Andrews (1910). Brown(1981) redescribes this taxon. Known from one fairlycomplete skeleton.
GENUS: Kimmerosaurus Brown, 1981
TYPE SPECIES: Kimmerosaurus langhami Brown, 1981.TYPE MATERIAL: — Holotype: BMNH R.8431.TYPE LOCALITY: Kimmeridge Clay, Dorset, England.AGE: Kimmeridgian.REFERRED MATERIAL: BMNH R.1798, R.10042.REMARKS: A poorly known but important taxon. Additionalmaterial described by Brown et al. (1986). The genusColymbosaurus, also from the Kimmerage Clay, may berelated to this taxon; however, no cranial material ofColymbosaurus is yet known (Brown 1984), and the taxonis in need of revision. The humerus of Colymbosaurus isvery similar to that of Polycotylus (Brown, 1981).
GENUS: Morturneria Chatterjee & Small, 1989
TYPE SPECIES: Morturneria seymourensis Chatterjee &Small, 1989
ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 17
TYPE MATERIAL: — Holotype: TT VP9219.TYPE LOCALITY: Lopez de Bertodana Formation, SeymourIsland, Antarctica.AGE: Maastrichtian.REFERRED MATERIAL: none.REMARKS: This taxon is the best-known of a poorly-known groupof aberrant Cretaceous cryptoclidids. This group also includes thetaxon Aristonectes parvidens Cabrera 1941, from South Ameri-ca; like Morturneria this taxon is Maastrichtian in age.
GENUS: currently unnamed
TYPE SPECIES: n/a.TYPE MATERIAL: — Holotype: MOR 751.TYPE LOCALITY: Thermopolis Shale, Montana, USA.AGE: Upper Albian.REFERRED MATERIAL: none.REMARKS: This specimen is an unnamed, primitive poly-cotylid described in a thesis by Druckenmiller (1998).Druckenmiller has a publication naming this taxon inpress (Druckenmiller pers. com.).
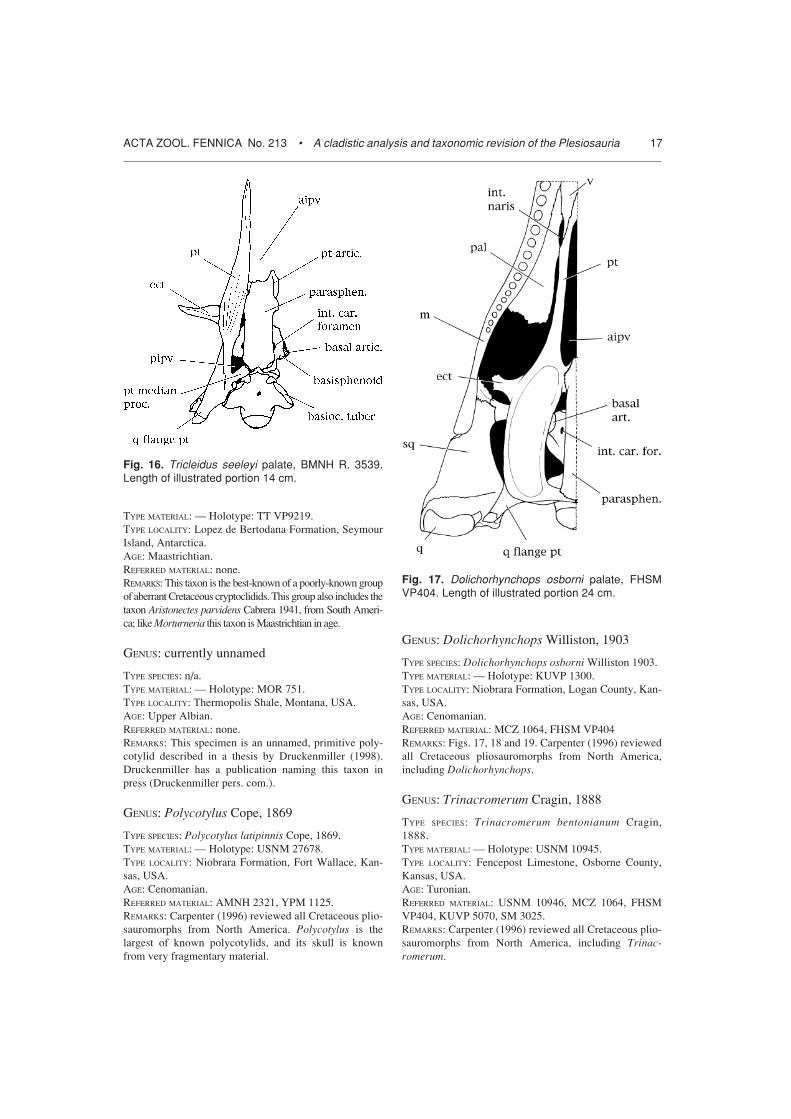
GENUS: Polycotylus Cope, 1869
TYPE SPECIES: Polycotylus latipinnis Cope, 1869.TYPE MATERIAL: — Holotype: USNM 27678.TYPE LOCALITY: Niobrara Formation, Fort Wallace, Kan-sas, USA.AGE: Cenomanian.REFERRED MATERIAL: AMNH 2321, YPM 1125.REMARKS: Carpenter (1996) reviewed all Cretaceous plio-sauromorphs from North America. Polycotylus is thelargest of known polycotylids, and its skull is knownfrom very fragmentary material.
GENUS: Dolichorhynchops Williston, 1903
TYPE SPECIES: Dolichorhynchops osborni Williston 1903.TYPE MATERIAL: — Holotype: KUVP 1300.TYPE LOCALITY: Niobrara Formation, Logan County, Kan-sas, USA.AGE: Cenomanian.REFERRED MATERIAL: MCZ 1064, FHSM VP404REMARKS: Figs. 17, 18 and 19. Carpenter (1996) reviewedall Cretaceous pliosauromorphs from North America,including Dolichorhynchops.
GENUS: Trinacromerum Cragin, 1888
TYPE SPECIES: Trinacromerum bentonianum Cragin,1888.TYPE MATERIAL: — Holotype: USNM 10945.TYPE LOCALITY: Fencepost Limestone, Osborne County,Kansas, USA.AGE: Turonian.REFERRED MATERIAL: USNM 10946, MCZ 1064, FHSMVP404, KUVP 5070, SM 3025.REMARKS: Carpenter (1996) reviewed all Cretaceous plio-sauromorphs from North America, including Trinac-romerum.
Fig. 16. Tricleidus seeleyi palate, BMNH R. 3539.Length of illustrated portion 14 cm.
Fig. 17. Dolichorhynchops osborni palate, FHSMVP404. Length of illustrated portion 24 cm.

O’Keefe • ACTA ZOOL. FENNICA No. 21318
GENUS: Attenborosaurus Bakker, 1993
TYPE SPECIES: Attenborosaurus conybeari (Sollas, 1881).TYPE MATERIAL: — Holotype: lost; BMNH R.1339 (cast).TYPE LOCALITY: Charmouth, Dorset, England.AGE: Sinemurian (?).REFERRED MATERIAL: BMNH 40140, BMNH R.1360, BMNH39514.REMARKS: Described by Owen (1865) as ‘Plesiosaurusrostratus’, material referable to this taxon was renamedby Bakker (1993) to replace ‘Plesiosaurus’ conybeari(Sollas 1881), after Persson (1963) remarked that the
specimen was clearly not a member of the genus Plesio-saurus. Persson also noted that the skull was very similarto that of Rhomaleosaurus, but the postcranium wasplesiosauromorph; see O’Keefe (2002) for interpretation.
GENUS: Kronosaurus Longman, 1924
TYPE SPECIES: Kronosaurus queenslandicus Longman, 1924.TYPE MATERIAL: — Holotype: Queensland Museum;Longman (1924, 1930) does not list specimen number.TYPE LOCALITY: Army Downs, north of Richmond, Queens-land, Australia.REFERRED MATERIAL: MCZ 1284, 1285.REMARKS: Kronosaurus was originally described on thebasis of a fragment of mandibular symphysis and propo-dial fragments. The more complete Harvard material wascollected in 1931–1932 and described by White (1935).The Harvard skull has been on loan to C. McHenry forsome time, who is working on a redescription of thistaxon based on new material (C. McHenry pers. com.).An additional complete skeleton was assigned to thisgenus by Hampe (1992). Hampe also furnishes 22 char-acters and a cladogram of seven ‘pliosaur’ species.
Characters and coding
The 34 taxa listed above were scored for 166morphological characters. Of these characters,107 concerned the skull and 59 were postcrani-al. About half of the characters appear in theliterature in some form. Appendix 1 gives anumber and name for each character, the refer-ence (if any) for that character, a description ofstates, numerical codings, and any relevant re-marks. The references cited in the table are themost informative and generally the most recent
Fig. 18. Dolichorhynchopsosborni skull roof in ob-lique lateral view, MCZ1064. Length of illustrat-ed portion 39 cm.
Fig. 19. Dolichorhynchops osborni mandible, FHSMVP404; lateral (top), dorsal (middle), lingual (bot-tom). Length of illustrated portion 19 cm.

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 19
source for a given character, and the listing isnot exhaustive. The figures where states of agiven character are illustrated are listed in theremarks section for each character. Charactersare scored with the primitive state as zero forconvenience, and polarity was determined bycomparison with the outgroup. Most charactersare unordered; those that are ordered are notedin the remarks section. Characters are only or-dered if a second state is logically dependent onthe presence of the first state, such as charactersconcerning successive neck lengths or thenumber of maxillary teeth. Running all charac-ters as unordered results in identical results asrunning some characters as ordered. The matrixhas a relatively high proportion of charactersthat are inapplicable to some taxa, due largely to
the profound reorganizations of the pectoralgirdle and palate between the outgroup and theingroup. Characters which code only for theoutgroup were included to help establish thetopology within the outgroup, which can effectcharacter reconstruction at the basal node of theingroup and hence the ingroup topology. Char-acters which code only for the ingroup willaffect only the topology of the ingroup. Thepresence of both types of characters will notgive rise to spurious effects as long as the basalingroup node is strongly supported by othercharacters, and it is in this case (bootstrap value86%, decay index 6; see Fig. 20).
The data matrix (Appendix 2) was analyzedusing PAUP 3.1.1 (Swofford & Begle 1993).The non-plesiosaur taxa Simosaurus, Cymato-
Fig. 20. Phylogenetic relationships of the Plesiosauria. Topology shown is the strict consensus of 12 MPTs,tree lengths 432. CI = 0.47, RCI = 0.34, RI = 0.72. Numbers just below and to the left of a given node arebootstrap support values; those to the right are decay indices. Indices in parantheses are the support for agiven node after morphometric characters have been removed and analysis rerun. Dots represent nodeswith less than 50% bootstrap support and a decay index of one. Named groups are diagnosed in the text.

O’Keefe • ACTA ZOOL. FENNICA No. 21320
saurus, and Pistosauridae were specified as theoutgroup, and the outgroup was constrained tobe paraphyletic with respect to the ingroup toreflect the topology of Rieppel (2000). Theheuristic search strategy was used, using tree-bisection-reconnection (TBR) branch swapping.The reference taxon was random; iterationsshowed that using any taxon as the referencetaxon yielded the same set of trees. Initial analy-sis was run with all characters included andweighted equally. A second analysis was per-formed with the ‘morphometric’ characters re-moved, based on the finding that the pliosauro-morph body type may have evolved convergent-ly (O’Keefe 2002).
Results
Parsimony analysis of the data matrix yieldedtwelve most- parsimonious trees (MPTs), eachwith a tree length of 432. The strict consensustree computed from the twelve MPTs is shownin Fig. 20. The MPTs had a Consistency Index(CI) of 0.47, and a Rescaled Consistency Index(RCI) of 0.34 (excluding uninformative charac-ters). The Retention Index (RI) was 0.72. Iinvestigated the robustness of the strict consen-sus tree topology by bootstrapping the characterlist (1000 replicates), and by calculating decayindices (Bremer 1994) for each ingroup node.Bootstrap percentages and decay indices arestated beneath each node in Fig. 20. The tree iswell-supported with the exception of some cryp-tocleidoid relationships and the ingroup rela-tionships of the rhomaleosaurids.
The topology of clade Plesiosauria replicatesmany of the findings advanced by earlier work-ers. The basic dichotomy between the Plesiosau-roidea and the Pliosauroidea is a well-supportedfinding. Within the Pliosauroidea two largemonophyletic groups exist, the Rhomaleosauri-dae and the Pliosauridae, as suggested by Car-penter (1997). The base of the clade Pliosauroi-dea contains the plesiomorphic taxon Thalassio-dracon, as well as the plesiomorphic and transi-tional Eurycleidus. The basal position of Eury-cleidus was predicted by Cruickshank (1994b).The Polycotylidae is not a member of the Plio-sauroidea, establishing that the pliosauromorph
body type is a polyphyletic grade. To investigatethe stability of this result, parsimony analysiswas performed with the constraint of pliosau-roid/polycotylid monophyly in effect. This anal-ysis resulted in two MPTs with tree lengths of447, fifteen steps longer than the MPTs from theoriginal analysis. This large increase in treelength indicates that polyphyly of the traditionalPliosauroidea is a well-supported finding, andthat monophyly of the traditional Pliosauroideais unlikely.
Within the Plesiosauroidea, the monophyly ofthe Elasmosauridae is well-supported. TheCryptocleidoidea, however, is the one group inwhich the cladogram departs radically from thetraditional taxonomy. The placement of Mu-raenosaurus in this clade suggests that Muraeno-saurus is not an elasmosaur (contra Andrews1910). The elasmosaur-like long neck and smallhead evolved independently in Muraenosaurus.Also within the Cryptocleidoidea is a novel cladecomposed of Tricleidus, the Polycotylidae, andthe short-necked cryptocleidoids Kimmerosaurusand Morturneria. This clade documents the deri-vation of the pliosauromorph polycotylids from aTricleidus-like cryptocleidoid. Taxonomy ofclade Plesiosauria is revised below in the system-atic paleontology section of the discussion. Thetaxonomic revision is followed by a discussion ofcharacter transitions at various nodes.
To investigate the effect of morphometriccharacters on the tree topology, a PAUP analy-sis was run with the eight morphometric char-acters excluded (characters 1–7, 112). Thisanalysis resulted in 6 MPTs, each with a treelength of 387. The CI of these trees was 0.49,and the RCI was 0.36. The RI was 0.73. Thetopology of the strict consensus tree computedfrom this set of MPTs was almost identical tothe topology in the first analysis, differing onlyin the position of Microcleidus. In the firstanalysis Microcleidus formed a polytomy withtwo large clades, the elasmosaurs and crypto-cleidoids. The second, non-morphometric anal-ysis resolved this polytomy so that Microcleid-us became the outgroup to a clade in which theelasmosaurs and cryptocleidoids were sistertaxa. Microcleidus is a plesiomorphic and prob-lematic taxon, and it is treated as a stem taxonin this report.

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 21
The one area where the removal of themorphometric characters had a significant effectwas the tree length, and the RCI derived from it.The RCI increased from 0.34 to 0.36 in thesecond analysis. The MPTs derived from thesecond analysis were 47 steps shorter than thosederived from the first; this is an average ofalmost six steps for each of the eight morpho-metric characters. The average number of stepsper character for the rest of the data is about 2.5.The length of the neck and skull, and othercharacters of body proportion, are thereforemore labile than the remaining characters. Theobservation that the number of steps per mor-phometric character is about three times theaverage for the other characters can be ex-plained by the fact that the pliosauromorph bodytype evolved independently three times. Decayindices calculated on this tree showed minorimprovement for several nodes, and the reviseddecay indices appear in Fig. 20 in parenthesesnext to the indices from the primary analysis.
Discussion
The first section of this discussion is a revisionof the higher taxonomy of the Plesiosauria,based on the cladogram topology in Fig. 20. Thetaxonomic revision is followed by a short dis-cussion of characters important in the diagnosisof various clades and a short review of morpho-logical evolution in the Plesiosauria.
Systematic paleontology
The following is a revised taxonomy of thePlesiosauria. All diagnostic characters are de-scribed in Appendix 1, and scoring can be foundin the character matrix in Appendix 2. Allsynapomorphies listed in the diagnoses are unam-biguous. Character reconstructions that differ be-tween ACCTRAN and DELTRAN optimizationsare not listed. Only genera included in the presentanalysis are listed; note that the taxonomic sam-pling in this analysis is not exhaustive at thegenus level, especially within the Elasmosauridaeand Rhomaleosauridae. The definitions givenhere are node-based (see Sereno 1998, and refer-
ences therein for discussion). Taxa defined on thebasis of more than two taxa refer to clades whosebasal node is well-supported but whose internalrelationships are obscured by polytomy.
Sauropterygia Owen, 1860
DIAGNOSIS: diagnosed in Rieppel 1998.DEFINITION: A monophyletic taxon including thePlacodontia and the Eosauropterygia.
Pistosauria Rieppel, 1998
DIAGNOSIS: diagnosed in Rieppel 1998.DEFINITION: A monophyletic taxon including Cy-matosaurus, Pistosaurus, and the Plesiosauria.
Plesiosauria de Blainville, 1835
REVISED DIAGNOSIS: Maxilla/squamosal contactpresent; quadrate embayed anteriorly; parasphe-noid is exposed anterior to posterior interptery-goid vacuities on palate surface; cervical zygo-pophyses narrower than centrum width; dorsalneural arches shorter than centrum height; zy-gosphene/zygantrum articulation absent; longi-tudinal pectoral bar present; lack of articulationbetween ilium and pubis; expanded distal propo-dials with dorsal trochanter/tuberosity; ulna dis-tinctly lunate; shifted fifth metapodial present;hyperphalangy present.REVISED DEFINITION: A taxon including the Plesi-osauroidea, the Pliosauroidea, their most recentcommon ancestor, and all descendants.
Plesiosauroidea Welles, 1943
REVISED DEFINITION: A monophyletic taxon in-cluding Plesiosaurus and the Euplesiosauria,their most recent common ancestor, and alldescendants.REVISED DIAGNOSIS: Prefrontal contacts margin ofexternal naris; nasal absent; cervical neural spinesnot angled backward; dorsal neural spines com-pressed and blade-like.
Euplesiosauria (new taxon)
DIAGNOSIS: Relative skull length short; anteriorborder of pineal foramen formed by frontal;long lateral and short medial posterior processesof squamosal; jugal confined to posterior orbit

O’Keefe • ACTA ZOOL. FENNICA No. 21322
margin; supraoccipital deep with sigmoid ven-tral sutures; ectopterygoid contacts postorbitalbar out of palate plane; distinct change in angleof cervical zygopophyses; anterior processes ofscapula meet in median symphysis; longitudinalpectoral bar present, formed by coracoid andscapula; humerus not angled.DEFINITION: A taxon including the Elasmosauri-dae, the Cryptocleidoidea, and Microcleidus,their most recent common ancestor, and alldescendants.
FAMILY: Elasmosauridae Cope, 1870
REVISED DIAGNOSIS: Anterior quadrate embay-ment absent (reversal); premaxilla excludedfrom border of internal naris; vomer extendsposterior to internal nares; number of cervicalrib heads reduced to one; coracoids long withdeep median embayment; ventro-medial marginof pubis concave; ulna not lunate (reversal);epipodials wider than long.REVISED DEFINITION: A taxon including Branca-saurus, Styxosaurus, their most recent commonancestor, and all descendants.
Cryptocleidoidea Williston, 1925
REVISED DIAGNOSIS: Fin aspect ratio low; anteriorinterpterygoid vacuity wide with rounded ends;anterior parasphenoid short and blunt; coronoidexposed on lateral mandible surface; grooveformed by prearticular/socket in angular; atlascentrum exposed on lateral surface betweenintercentrum and neural arch; anterior neuralflange on cervical neural spines.REVISED DEFINITION: A taxon including the Cryp-toclididae, the Tricleidia, their most recent com-mon ancestor, and all descendants.
FAMILY: Cryptocleididae Williston, 1925
REVISED DIAGNOSIS: Humerus longer than femur,occipital condyle short with no groove, cervicalrib heads elongate.REVISED DEFINITION: A taxon including Muraeno-saurus, Cryptoclidus, their most recent commonancestor, and all descendants.
Tricleidia (new taxon)
DIAGNOSIS: Pterygoid medial process for articu-
lation with parasphenoid present; parasphenoidcontacts basioccipital on midline; basioccipitaltubers reduced and confluent with basisphenoidarticulation; cervical neural spines not com-pressed (reversal); clavicles meet in medialsymphysis behind notch.DEFINITION: A taxon including the Polycotylidae,the Cimoliasauridae, Tricleidus, their most re-cent common ancestor, and all descendants.
FAMILY: Cimoliasauridae Delair, 1959
REVISED DIAGNOSIS: Rostrum long, unconstricted,and hoop-like; paraoccipital process articulateswith squamosal only; teeth very small and nee-dle-like; number of premaxillary teeth seven orgreater; number of maxillary teeth much greaterthan thirty.REVISED DEFINITION: A taxon including Morturn-eria, Kimmerosaurus, their most recent commonancestor, and all descendants.
FAMILY: Polycotylidae Williston, 1908
REVISED DIAGNOSIS: Neck length short; ischiumlonger than pubis; maxillary/squamosal suturepresent and formed by posterior expansion ofmaxilla; pterygoids with distinct medial proc-esses which meet behind posterior interptery-goid vacuities; pterygoids dished lateral to pos-terior interpterygoid vacuities; mandibular sym-physis scoop-like or long; splenial included inmandibular symphysis; number of cervical ver-tebrae reduced; longitudinal pectoral bar presentand formed by clavicle and coracoid; posteriorperforations in coracoid; supernumerary ossifi-cations in propodial and epipodial rows; inter-locking phalanges anterior to fifth digit.REVISED DEFINITION: A taxon including Polycoty-lus, Dolichorhynchops, their most recent com-mon ancestor, and all descendants.
Pliosauroidea Welles, 1943
REVISED DIAGNOSIS: Snout constricted at premax-illa/maxilla suture; occipital condyle short withno groove; posterior bulb formed by squamosalspresent; pterygoids meet anterior to posteriorinterpterygoid vacuity; premaxilla excludedfrom internal naris margin; mandibular symphy-sis scoop-like or long; splenial included in man-

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 23
dibular symphysis; ventral keel present on cervi-cal vertebrae; posterior articulation for succeed-ing neural spine in cervical neural spines absent.REVISED DEFINITION: A taxon including Thalassi-odracon, an unnamed clade comprised of Eury-cleidus, Attenborosaurus, the Rhomaleosauri-dae, and the Pliosauridae, their most recent com-mon ancestor, and all descendants.
FAMILY: Pliosauridae Seeley, 1874 (Seeley, 1874)
REVISED DIAGNOSIS: Preorbital skull length longerthan postorbital skull length; rostrum elongate;no contact between premaxilla and externalnaris; distinct postero-lateral process of frontalabsent (reversal); anterior interpterygoid vacuityabsent (reversal); vomer reaches past internalnares and meets pterygoid in wide interdigitat-ing suture; mandibular symphysis long; numberof maxillary teeth between twenty and thirty;humerus not angled.REVISED DEFINITION: A taxon including Macro-plata, Brachauchenius, their most recent com-mon ancestor, and all descendants.
FAMILY: Rhomaleosauridae (Kuhn, 1961)
REVISED DIAGNOSIS: Grooves in front of externalnares present; paraoccipital process robust;squared lappet of pterygoid underlying ptery-goid quadrate ramus present; lateral palatal fen-estration present; bowed maxilla present; pre-maxillary/dentary fangs present; cervical centralengths less than heights; cervical zygopophysesas wide as centra (reversal).REVISED DEFINITION: A taxon including Rhomale-osaurus victor, Leptocleidus, their most recentcommon ancestor, and all descendants.
Comments on the classification
The above classification is based on the moreconservative first parsimony analysis, in whichall characters were included. The morphometriccharacters have high levels of homoplasy; suchhigh levels of convergence can arise, however,in rapidly-evolving characters, and can holdphylogenetic information, especially withinsmaller subclades (Chippindale & Wiens 1994).It is therefore conservative to base taxonomic
conclusions on the MPTs from all data. Nodesthat were not replicated in the non-morphomet-ric tree were not used in the classification.Taxonomic conclusions are also not based onnodes which have weak decay indices and boot-strap support. A good example is the Rhomaleo-sauridae. The genus Rhomaleosaurus is para-phyletic according to the cladogram althoughthe decay index and bootstrap values are low(decay index one, bootstrap support < 50%) forall rhomaleosaurid ingroup relationships. A for-mal revision of Rhomaleosaurus is best left untila more detailed cladistic analysis of the Rhoma-leosauridae is made.
The revised classification splits the Plesiosau-ria into four major subclades, two within thePlesiosauroidea and two within the Pliosauroidea.Within the clade Plesiosauroidea, the Elasmosau-ridae remains as traditionally defined, with Bran-casaurus as the most basal elasmosaur. See Car-penter (1999) for a more complete list of elasmo-saur taxa and a revised taxonomy of this clade.The Cryptocleidoidea is revised here to includethe Polycotylidae and Muraenosaurus; further-more, a new clade is recognized (the Tricleidia)comprising the Cimoliasauridae, the polycotylids,and Tricleidus. Tricleidus is a generalized form,falling out in a trichotomy with the other mem-bers of the Tricleidia. Tricleidus possesses sever-al synapomorphies linking it with the short-necked plesiosauroids, including the presence ofposterior medial processes of the pterygoids, re-duced basioccipital tubers, a median contact be-tween the basioccipital and parasphenoid, and thepossession of a third distinct articulation on thepropodials for a supernumery ossification in theepipodial row. All short-necked plesiosauroidsshare these traits.
A last, problematic taxon within the Plesio-sauroidea is Microcleidus. This taxon possessesseveral plesiomorphic characters such as reten-tion of a lunate ulna and retention of an angledhumerus; however, the skull is elasmosaur-likein the absence of an anterior interpterygoidvacuity and in the location of the jaw articula-tion (below the tooth row). The coracoids areknown from fragmentary material, but appearnot to have the posterior extensions and embay-ment synapomorphic of true elasmosaurs. Theneck is long and the cervical rib heads elongate,

O’Keefe • ACTA ZOOL. FENNICA No. 21324
although these characters are homoplastic. Be-cause of this homoplasy the topology given bythe morphometric-omitted cladogram is proba-bly more accurate in this case, with Microcleid-us forming the sister taxon to a clade consistingof the Elasmosauridae and Cryptocleidoidea.The outgroup to this clade is the very plesiomor-phic Plesiosaurus. A thorough redescription ofthis taxon based on the type material as well asanother complete skeleton at Manchester shouldshed more light on this genus.
The clade Pliosauroidea includes two long-recognized clades of pliosaurs, the Rhomaleo-sauridae and the Pliosauridae, as well as theplesiomorphic stem taxa Thalassiodracon, Eury-cleidus, and Attenborosaurus. Of the two majorpliosauroid clades, the Pliosauridae possessesmore derived characters. The rhomaleosauridsare conservative, similar to Thalassiodracon(and hence plesiomorphic) in the palate, skullroof, and postcranium. This conservatism ispartially responsible for the lack of support forintrafamilial rhomaleosaurid relationships. Chang-es in relative neck and skull length and a greatincrease in body size are the only known trendsin this clade.
In contrast to the Rhomaleosauridae, thePliosauridae is morphologically derived, andgood intermediates exist that document the deri-vation of the pliosaurid conditions of the skullroof, palate and braincase. The monophyly ofthe Pliosauridae is a correspondingly well-sup-ported finding. The plesiomorphic taxa Eury-cleidus and Attenborosaurus form a polytomywith the Pliosauridae and Rhomaleosauridae.The taxonomic status of these genera are leftopen, and the Pliosauridae is limited to Macro-plata and more derived taxa.
Major patterns in plesiosaur cranialevolution
This overview describes the morphology of thefour major plesiosaur subclades in variable re-gions of the skull. These areas are the skull roof,the palate, and the braincase. Discussion of theanatomy characteristic of each subclade is in-tended as an aid to future classification.
The skull roof
An important feature of the skull roof is theretention of the nasals in Thalassiodracon andthe rest of the Pliosauroidea. Nasals werethought to be absent in all plesiosaurs by Storrs(1991); however, the loss of nasals diagnosesonly the Plesiosauroidea, not the Plesiosauria asa whole. The nasals in all pliosauroid taxa aresmall but present, and usually form the posteriormargin of the external nares. The pliosauroidcondition is very similar to that of Cymatosau-rus. In some very derived members of the Plio-sauridae, the nasal forms the anterior rather thanthe posterior margin of the external naris. Thenasals in all pliosaurs lack a midline suture,being separated on the midline by processes ofthe premaxillae.
The pattern of circumorbital bones is diag-nostic within the Plesiosauria. The lacrimal isabsent in all plesiosaurs due to its loss in morebasal sauropterygians (Rieppel 1997b). The pre-frontal and postfrontal do not exclude the frontalfrom the orbital margin in early plesiosaurs, andthis condition is retained in all plesiosauroids,including the polycotylids. However, the frontalis excluded from the orbital margin in all morederived rhomaleosaurids and pliosaurids. Thejugal is also diagnostic; in the most primitiveplesiosauroids and pliosauroids the jugal reach-es anteriorly to about the midpoint of the ventralorbital margin, which is plesiomorphic (Rieppel1997b). In more derived plesiosauroids the jugalis restricted to the posterior orbital margin. Thiscondition contrasts with that in more derivedpliosauroids, where the jugal reaches far anteriorto the orbit and forms its anterior border, meet-ing the prefrontal and excluding the maxillacompletely from the orbital margin. The pres-ence of a prominent anterior process of the jugaland its contact with the prefrontal probablyaccount for its misidentification as a lacrimal byearlier workers (Andrews 1913, Taylor &Cruickshank 1993).
The cryptocleidoids are extreme in their spe-cialization of the cheek region (Brown &Cruickshank 1994, Brown 1993). In Cryptoclid-us and Tricleidus the contact between maxillaand squamosal is lost, and the jugal forms a

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 25
narrow bar connecting these two elements be-neath the orbit. This area of the cheek is notknown in Muraenosaurus or the Cimoliasauri-dae. This condition is reversed in the polycoty-lids, where the maxilla again forms a suture withthe squamosal. The morphology of this suturediffers from the corresponding suture in truepliosaurs. In true pliosaurs the maxilla/squamos-al suture is small and not significant, and thejugal is large and has a prominent lateral suturewith the squamosal. In the polycotylids the jugalis small and restricted to the posterior orbitalmargin, and does not form a prominent suturewith the squamosal. The major suture in thisregion is that between the maxilla and squamos-al, which is formed by an expanded posteriorprocess of the maxilla. A possible explanationfor this pattern is secondary strengthening ofthis area of the skull made necessary by theevolution of a long snout in taxa with a highlyreduced cheek region, such as Tricleidus.
A feature that evolves repeatedly in long-snouted taxa is a contact between the dorsalprocesses of the premaxillae and the parietals. Amedian premaxilla/parietal suture evolves fourtimes, once each in the three derivations of thepliosauromorph body type and once in relativelylong-snouted elasmosaurs. This suture probablyserves to strengthen the interorbital region of theskull in response to greater forces produced at thetip of the long snout. A long snout is alsogenerally correlated with larger tooth size and amore robust skull in general; larger teeth proba-bly indicate larger average prey size and a con-comitant need to reinforce the skull. The robustmaxilla/squamosal suture in the polycotylids isalso interpretable as a means of strengthening theskull in response to increased snout length.
The palate
The primitive plesiosaur palate is derived com-pared to the condition in more basal sauroptery-gians. The plesiomorphic condition for the ple-siosaur palate is shown by Thalassiodracon andPlesiosaurus; both taxa possess anterior andposterior interpterygoid vacuities exposing theventral surface of the braincase. The pterygoids
do not meet behind the posterior interpterygoidvacuities in either taxon. In Plesiosaurus, thepterygoids also fail to meet between the posteri-or and anterior vacuities, exposing the cultri-form process of the parasphenoid (Storrs 1997).This open palate is retained (with some modifi-cation) in all members of the Cryptocleidoidea.In the Elasmosauridae and Microcleidus, how-ever, the anterior interpterygoid vacuity is ab-sent. The pterygoids also meet in a prominentmidline suture behind the posterior interptery-goid vacuity in all elasmosaurs, but not in Mi-crocleidus. Elasmosaurs are therefore character-ized by a secondarily closed palate, and thiscondition is foreshadowed to some extent inMicrocleidus.
The anterior interpterygoid vacuity is small inThalassiodracon, and is separated from the poste-rior vacuities by a long midline suture of thepterygoids. This pattern is retained withoutchange in the rhomaleosaurids. The rhomaleosau-rids are advanced over Thalassiodracon, howev-er, in possessing a contact of the pterygoidsbehind the posterior interpterygoid vacuity; thiscontact is partial in the plesiomorphic speciesRhomaleosaurus megacephalus and complete inall later rhomaleosaurids. The anterior palate ofrhomaleosaurs is like that of Eurycleidus, andunlike that of derived pliosaurids, in that thevomers do not extend far posterior to the internalnares. The Thalassiodracon material is not suf-ficiently well preserved to establish the presenceor absence of sub-orbital fenestrae (Storrs &Taylor 1997). This feature occurs in some rhoma-leosaurids and some pliosaurids. The presence orabsence of sub-orbital fenestrae can be difficult toestablish because the palatine is very thin and israrely preserved intact.
In contrast to that of rhomaleosaurids, thepalate in pliosaurids is highly derived comparedto the plesiomorphic condition in Thalassiodra-con. The anterior interpterygoid vacuity isclosed in the intermediate form Macroplata. Thevomers extend posterior to the internal nares andmeet the pterygoids in a wide, interdigitatingsuture in Macroplata, as well as in Hauffiosau-rus and all more derived pliosaurids. The quad-rate flange of the pterygoid is wide, sigmoid inshape, and possesses a ridged lateral margin in

O’Keefe • ACTA ZOOL. FENNICA No. 21326
Kronosaurus, Peloneustes, Liopleurodon, Plio-saurus, and Brachauchenius. The pliosauridsMacroplata and Hauffiosaurus are plesiomor-phic for this character; however, Attenborosau-rus is derived, possessing the ridged lateralmargin and sigmoid shape of the quadrate ptery-goid flange. Attenborosaurus is intermediate foranother synapomorphy of derived pliosaurids,namely the elaboration of lateral flanges of thepterygoid on either side of the posterior interpt-erygoid vacuities. These flanges are present inKronosaurus and all more derived pliosaurids,although they fuse over the posterior end of theinterpterygoid vacuity in Liopleurodon andmore derived forms. The elaboration of theseflanges gives the posterior palate a distinct two-tiered morphology in lateral view, with thelateral flanges, ventral braincase structures, andinterpterygoid vacuity on a more ventral leveland the quadrate pterygoid flanges reaching dor-sally and caudally back toward the quadrates ona more dorsal level. The polycotylids show notrace of this condition; the palate in this group isvery similar to the cryptocleidoid pattern, asobserved by Carpenter (1997).
The mandible
The plesiosaurian mandible has been a source ofconfusion for many years. The presence of thecoronoid and prearticular bones has been debat-ed; Andrews illustrates both bones as absent inMuraenosaurus (1910) and the prearticular asabsent in Peloneustes (1913). Cruickshank(1994b) states that absence of the coronoid istypical of plesiosauroids, and that the prearticu-lar is usually absent as well. Storrs and Taylor(1996) illustrate a prearticular but no coronoidin Thalassiodracon, while Taylor (1992b) illus-trates both bones as present in Rhomaleosaurus.I believe the prearticular is present in all taxawith adequately preserved material. The coro-noid is present in all taxa but Kimmerosaurus.The morphology of the lower jaw in plesiosau-roids is illustrated here by Libonectes (Fig. 15)and Dolichorhynchops (Fig. 19), and that ofpliosauroids by Peloneustes (Fig. 12). The lin-gual surface of the lower jaw is poorly ossifiedin plesiosaurs and the Meckelian canal is often
open for some of its length. The bones coveringthe Meckelian canal (splenial, coronoid, andprearticular) are very thin and often damaged orlost in fossils, which may account for the confu-sion concerning the two smaller bones.
The lower jaw symphysis was taken as animportant taxonomic character by Tarlo (1960)and later workers. The simple, unexpanded sym-physis present in most plesiosauroids is in factderived over the primitive condition exemplifiedby Thalassiodracon (Storrs & Taylor 1996) orthe outgroup taxon Cymatosaurus (Rieppel1997b). In these taxa the symphysis is rein-forced and scoop-like (Rieppel 1997b, character51), and the presence of a scoop-like symphysisin Rhomaleosaurus and related taxa is plesio-morphic. The snout and lower jaw symphysisbecomes very elongate in the most primitivemembers of the Pliosauridae and remains astereotyped feature of that family. The length ofthe symphysis does vary within this group,remaining very long in Peloneustes and revert-ing to scoop-like in Liopleurodon, Pliosaurus,and related taxa. A long lower jaw symphysis isalso present in all members of the Polycotylidae,where a long snout evolved convergently.
The braincase
The morphology of the braincase is known tovarying degrees in different plesiosaur subc-lades. In the elasmosaurids and rhomaleosauridsvery little is known, because a broad suture ofthe pterygoids between the posterior and anteri-or interpterygoid vacuities obscures most of theanterior braincase in ventral view (although seeCarpenter, 1997, for comments on the moredorsal elements of the elasmosaur braincase).More is known concerning the cryptocleidoidsand pliosaurids, in which the posterior interpt-erygoid vacuities are larger and open fartheranteriorly. In addition, the brain case is not astightly integrated into the skull roof in the latertaxon, so that the elements are more often pre-served in a visible manner.
The braincases of Plesiosaurus and Thalas-siodracon are similar, and are very primitivewhen observed in ventral view. The occipitalcondyle is hemispherical in Plesiosaurus and set

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 27
off from the body of the basioccipital by agroove, while in Thalassiodracon the condyle isless rounded with no groove (Storrs and Taylor1996, Storrs 1997). These condyle morpholo-gies are characteristic of later plesiosauroids andpliosauroids, respectively. The occipital condylemay have a notochordal pit; the pit is moreobvious in material from younger animals, andthere is intraspecific variation in its presence.The body of the basioccipital is a short, robustblock of bone, with paired articulations on itsdorsal surface for the exoccipitals (see Brown1981; Fig. 1). The basioccipital produces twoprominent basioccipital tubers projecting ante-ro-ventrally that articulate with the pterygoids.The exoccipitals are columnar bones rising dor-sally on either side of the foramen magnum, andgiving rise laterally to slender paraoccipitalprocesses that trend laterally to articulate withthe squamosals. The occiput is therefore open,and the suture between exoccipital and opisthot-ic is seldom visible (Storrs and Taylor 1996).The medial surface of each exocciptal is piercedby the jugular foramen and by one or twoforamina for the passage of the hypoglossalnerve (Storrs and Taylor 1996, Carpenter 1997).
In Plesiosaurus and Thalassiodracon, thebasioccipital tubers also articulate on their ante-rior margins with lateral processes of the paras-phenoid. These lateral parasphenoid processesare termed cristae ventrolaterales following Re-isz (1981: p. 24). The possession of the cristaeventrolaterales is a plesiomorphic condition foundin Petrolacosaurus (Reisz 1981) and Araeoscelis(Vaughn 1955). In these plesiomorphic taxa, thecristae ventrolaterales are two crests of bonesurrounding a deep basisphenoid fossa on themidline, with the posterior margin of the basi-sphenoid visible in the floor of this fossa. Thecondition in basal plesiosaurs is similar, exceptthe basisphenoid fossa has been filled with a discof basisphenoid confluent with the ventral planeof the cristae ventrolaterales. In Petrolacosaurusand Araeoscelis, the body of the parasphenoidruns forward into a long, narrow cultriform proc-ess. Plesiosaurus has a similar cultriform proc-ess. The condition in Thalassiodracon is un-known due to that taxon’s possession of a medi-an pterygoid suture between the posterior andanterior interpterygoid vacuities. Ventral to the
parasphenoid in the region of origin of the cultri-form process is the dorsum sellae and the bodyof the basisphenoid (the clivus) just posterior toit, a shelf of bone which ossifies just posterior tothe sella turcica (Romer 1956). In both Plesio-saurus and Thalassiodracon the basisphenoid iswell ossified, and the basal articulations arisinglaterally from it are prominent. Like Araeoscelis,Thalassiodracon possesses foramina in the ven-tral surface of the basisphenoid for the passageof the paired internal carotid arteries. The pres-ence or absence of these foramina was impossi-ble to establish in Plesiosaurus. These foraminaare not ossified in Eurycleidus, and the basisphe-noid facet of the basal articulation was impossi-ble to identify as a distinct feature (Cruickshank1994b). These observations probably arise fromthe fact that the Eurycleidus braincase material isfrom a juvenile, and so is not well-ossified.However, lack of ossification of the basisphe-noid is also characteristic of later pliosaurids, sothe condition in Eurycleidus may foreshadowthis to some degree. All plesiosaurs share a lackof ossification of the lateral walls of the brain-case. The area of origination of the pila antoticafrom the dorsal surface of the basisphenoid istherefore impossible to identify.
Two major changes to the plesiomorphicpattern of the plesiosaur braincase can be identi-fied in more derived groups. In the cryptoclei-doids the palate remains open and the skull roofis gracile, while the braincase is well-ossified.The cristae ventrolaterales disappear and theparasphenoid loses the characteristic triangularshape present in Araeoscelis and Plesiosaurus.The parasphenoid becomes a massive, block-like medial structure extending caudally almostto the basioccipital in more primitive cryptoclei-doids, and forming a novel median suture withthe basioccipital in more derived taxa (seeFig. 21). The cultriform process is also lost; theanterior margin of the parasphenoid becomesbroad and blunt and develops novel articulationswith the pterygoids on its antero-lateral edges.In contrast to the parasphenoid, the basisphe-noid in this group is conservative. Facets for thebasal articulation are well-defined, and the inter-nal carotid foramina pierce the body of thebasisphenoid ventro-laterally and run into thefloor of the sella turcica.

O’Keefe • ACTA ZOOL. FENNICA No. 21328
Fig. 21. Schematic transformation series of the basicranium and posterior palate, Plesiosauroidea. Ho-mologous bones are indicated in shades of gray. The plesiomorphic condition for this clade is demon-strated by Plesiosaurus. The palate is broadly open, and the pterygoids lack a median suture betweenboth the anterior and posterior interpterygoid vacuities, and beneath the basiocciptal. The parasphenoid isexposed medially along its entire length, the cultriform process is wide and prominent, and is confluentwith a ventral eminence or ridge running posterior to the exposed basisphenoid on the midline. Thebasisphenoid is visible in ventral view in two areas: anteriorly the processes comprising the basalarticulation are visible in the anterior of the posterior interpterygoid vacuities, while posteriorly an island ofbasisphenoid is exposed between parasphenoid and basioccipital. In elasmosaurs, here represented byLibonectes, this pattern is modified by the formation of a median pterygoid suture over the basioccipitaland basisphenoid. The anterior interpterygoid vacuity is also closed. In Tricleidus, however, the anteriorinterpterygoid vacuity stays open and the cultriform process losses its pointed anterior end, insteaddeveloping prominent later sutures with the pterygoids. The ventral eminence of the parasphenoid reach-es almost to the basiocciptal, and is broad along its entire length. Each pterygoid develops a novelprocess reaching medially to the ventral eminence of the parasphenoid. The basal articulation is well-developed, and the ectopterygoid reaches dorsally to the ventral surface of the postorbital bar. Thesituation in Dolichorhynchops is very similar, except that the medial processes of the pterygoids meet in asuture ventral to the basiocciptal. Additionally, the ventral eminence of the parasphenoid reaches far tothe posterior, overriding the basiocciptal and the median suture of the pterygoids.
In derived pliosaurids, the skull roof andpterygoids are massive and well-ossified, where-as the braincase is poorly ossified. In pliosauridssuch as Peloneustes, the anterior margin of theparasphenoid articulates tightly with the ptery-goids at the anterior margin of the posterior
interpterygoid vacuities. From this articulationthe parasphenoid runs caudally, expanding intothe plesiomorphic triangular shape. However,the parasphenoid ends after a short distance,articulating with the basisphenoid on the mid-line. The cristae ventrolaterales are again absent.

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 29
The basisphenoid is usually preserved as a ru-gose and unremarkable barrel of bone betweenthe parasphenoid and the basioccipital. In lifethe basisphenoid was completely encased incartilage, and the dorsum sellae and sella turcicausually failed to ossify. One extremely well-preserved skull of Peloneustes (BMNH 3803)does preserve a poorly ossified basisphenoid.Enough morphology is visible on this specimento establish that the basal articulation is presentin the plesiomorphic location, and the pterygoidfacet of the basal articulation is usually identifia-ble in skulls of Peloneustes. Foramina for theinternal carotids were not preserved in this spec-imen. In conclusion, the braincase in pliosauridsis a poorly ossified structure showing few mor-phological details in most specimens; well-pre-served specimens seem to demonstrate that thecartilaginous elements of the braincase retainedthe plesiomorphic pattern seen in Thalassiodra-con. Trends in basicranium evolution are illus-trated in two summary figures (Figs. 21 and 22).
Trends in postcranial evolution
The most remarkable aspect of plesiosaur post-cranial evolution is the convergent attainment ofbody morphotypes. The ‘pliosauromorph’ bodytype, characterized by a relatively large head,short neck, relatively long coracoids and ischia,and low aspect-ratio paddles, evolves in threeplesiosaur clades (Rhomaleosauridae, Pliosauri-dae, Polycotylidae; for analysis and discussionsee O’Keefe 2002). This striking convergence isresponsible for the polyphyly of the Pliosauroideaas traditionally defined, because characters ofbody overall body proportion were the basis oftraditional taxonomy (e.g. Persson 1963). Thisanalysis strongly rejects the traditional hypothesisthat all pliosauromorph taxa form a monophyleticgroup. In addition, the long neck thought to becharacteristic of elasmosaurs exclusively alsoevolved in the cryptoclidid Muraenosaurus (con-tra Andrews 1910, Persson 1963).
Several specific trends in the limb girdlesand limbs also deserve mention. Stratigraphical-ly early and plesiomorphic plesiosaurs, includ-ing Thalassiodracon and Plesiosaurus, retain adistinct bend or angle in the shaft of the humerus
Fig. 22. Schematic transformation series of the ba-sicranium and posterior palate, Pliosauroidea. Ho-mologous bones are indicated in shades of gray.The plesiomorphic condition for this clade (andprobably for plesiosaurs in general) is demonstrat-ed here by Thalassiodracon. The pattern in thistaxon is very similar to that described for Plesio-saurus in Fig. 21, except that the pterygoids meetin a median suture anterior to the posterior interpt-erygoid vacuities, and the parasphenoid is suturedto the pterygoids on the midline. The cultriformprocess is not visible in ventral view. The anteriorinterpterygoid vacuity is present but not illustratedhere; it is visible in Fig. 4. The pattern in Rhomale-osaurus is very similar to Thalassiodracon, theonly difference being a median suture of the ptery-goids over the basiocciptal and basisphenoid (par-tial in R. megacephalus, complete in most otherrhomaleosaurs). By contrast the palate is very de-rived in true pliosaurs, here represented by Pelone-ustes. The parasphenoid is not ossified around thebasisphenoid as in other plesiosaurs, although giv-en that the parasphenoid is dermal bone it isdoubtful that it persisted in cartilage. The basisphe-noid is very poorly ossified, and the basal articula-tion was cartilaginous. The pterygoids form ventralflanges lateral to the posterior interpterygoid vacui-ties that curve medially and meet beneath the basi-occiptal. The quadrate flanges of the pterygoidsare prominent, sigmoid processes which overridethe lateral flanges of the pterygoids.

O’Keefe • ACTA ZOOL. FENNICA No. 21330
(Storrs 1997). All ‘nothosaur’-grade sauroptery-gians also have this bend (Rieppel 1997b), whilethe shaft of the humerus is straight in all morederived plesiosaurs (Storrs 1997). Correlatedwith this transition is the loss of the ‘lunate ulna’in more derived plesiosaurs; plesiomorphic ple-siosaurs have epipodials that, while short, arevery similar to those in ‘nothosaur’-grade sau-ropterygians, and recognizable as radius, ulna,tibia, and fibula (Storrs 1997). In more derivedplesiosaurs these bones become unidentifiableon the basis of morphology, being simple ossi-fied disks integrated into the other ossificationsof the flipper (Storrs 1993, Caldwell 1997a,1997b). The functional significance of thesetransitions has yet to be investigated. A lastcharacter of note is the presence of a mediansymphysis of the scapulae. All plesiosauroidswere thought to have this symphysis (Persson1963); however, the placing of the Polycotylidaewithin the Plesiosauroidea demonstrates thatthis character is reversed in the polycotylids.This placement may also explain the similaritiesin the humeri of Polycotylus and Colymbosau-rus, although revision of the later taxon is neces-sary. A character of unknown signifigance is thescalloped margin of the vertebral centra in manycryptocleidoids; this may be a synapomorphy ofthis clade but further research is necessary. Ifpresent, however, the character would form an-other link between the Polycotylidae and theCimoliasauridae. The Cimoliasauridae certainlydeserve further research effort, both on knownmaterial and through field work.
Conclusions
1. The Plesiosauria is a monophyletic clade ofsauropterygians most closely related to thePistosauridae.
2. The Plesiosauria is composed of variousstem taxa and four major derived clades.These clades are the Rhomaleosauridae andPliosauridae within the Pliosauroidea, andthe Cryptocleidoidea and Elasmosauridaewithin the Plesiosauroidea.
4. The Rhomaleosauridae is a highly conserva-tive, plesiomorphic clade, most similar to thebasal pliosauroid Thalassiodracon.
3. The Polycotylidae is not a member of thePliosauroidea, and is instead a derived groupof cryptocleidoids.
4. Morphometric characters relating to bodyproportions are very homoplastic in the Ple-siosauria, and the pliosauromorph body typeevolved three times.
5. Nasals are present in all members of thePliosauroidea.
6. The plesiosaur braincase is very primitive,comparable to that of primitive diapsids suchas Araeoscelis.
Acknowledgements
This project was part of a doctoral dissertationsubmitted to the University of Chicago. Thedissertation was advised by J. Hopson and O.Rieppel, whose constant patience and assistanceof all kinds were critical to the project’s comple-tion. J. Hopson allowed me access to R. T.Zanon’s unpublished research, which was veryhelpful in getting the project started. C. Abraczin-skas taught me to draw what I see and made thisproject possible. A. Dudek, D. and S. O’Keefe,and M. Silverman provided help and support of aphysical, emotional, and/or metaphysical nature.
The following people assisted in visits tocollections at various institutions, and I thankthem all for their hospitality and help of allkinds: M. Benton, R. Vaughan, G. Dyke, T.Sharpe, J. Nudds, A. Cruickshank, J. Martin, P.Manning, M. Evans, A. Dawn, P. Powell, D.Norman, M. Dorling, S. Chapman, A. Milner, S.Donovan, M. Taylor, K. Padian, P. Holroyd, S.Parris, D. Parris, D. Miao, L. Martin, S. Chatter-jee, M. Brett-Surman, M. Turner, G. Gaffney, C.Holton, D. Schaff, R. Schoch, R. Hauff, M.Maisch, R. Wild, R. Zakrzewski, D. Lintz, L.Jacobs, J. Head, D. Winkler.
This manuscript benefitted from thoroughand very helpful reviews by O. Rieppel, J.Hopson, M. Sander, and G. Storrs. Their help isgratefully acknowledged. This research wassponsored in part by grants from the Samuel P.Welles Fund, the University of Chicago Wom-en’s Board, the University of Chicago HindsFund, and a Nierman Award. An Area 271Publication.

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 31
References
Andrews, C. W. 1895: On the structure of the skull ofPeloneustes philarchus. — Annals and Magazine ofNatural History, London 16(6): 242–256.
Andrews, C. W. 1896: On the structure of the plesiosauri-an skull. — Quarterly Journal of the GeologicalSociety, London 52: 246–253.
Andrews, C. W. 1909: On some new Plesiosauria fromthe Oxford Clay of Peterborough. — Annals andMagazine of Natural History, London 4(8): 418–429.
Andrews, C. W. 1910: A catalogue of the marine reptilesof the Oxford Clay, Part I. — British Museum(Natural History), London, England.
Andrews, C. W. 1911a: Description of a new plesiosaur(Plesiosaurus capensis, sp. nov.) from the UitenhageBeds of Cape Colony. — Annals of the South AfricanMuseum 1: 309–322.
Andrews, C. W. 1911b: On the structure of the roof of theskull and of the mandible of Peloneustes, with someremarks on the plesiosaurian mandible generally. —Geological Magazine of London 8: 160–164.
Andrews, C. W. 1913: A descriptive catalogue of themarine reptiles of the Oxford Clay, Part II. — BritishMuseum (Natural History), London, England.
Andrews, C. W. 1922: Description of a new plesiosaurfrom the Weald Clay of Berwick (Sussex). — Quar-terly Journal of the Geological Society of London 78:285–295.
Anonymous. 1854: Report of the council of the YorkshirePhilosophical Society 1853: 7–8.
Bakker, R. 1993: Plesiosaur extinction cycles — events thatmark the beginning, middle, and end of the Cretaceous.— In: Caldwell, W. G. E. & Kauffman, E. G. (eds.),Evolution of the Western Interior Basin: 641–664.Geological Survey of Canada, Special Paper 39.
Bardet, N. 1998: A preliminary cladistic analysis of thePlesiosauria. — Journal of Vertebrate Paleontology18(3) supp.: 26A.
Benton, M. J. & Spencer, P. S. 1995: Fossil reptiles ofGreat Britain. — Chapman and Hall, London.
Blake, J. F. 1876: Plesiosaurus longirostrus. — Tate, R.& Blake, J. F. (eds.), The Yorkshire Lias: 250–252.Van Voorst, London.
Brown, D. S. 1981: The English Upper Jurassic Plesio-sauroidea (Reptilia) and a review of the phylogenyand classification of the Plesiosauria. — Bulletin ofthe British Museum of Natural History (geol.) 35(4):253–347.
Brown, D. S. 1984: Discovery of a specimen of theplesiosaur Colymbosaurus trochanterius (Owen) onthe island of Portland. — Proc. Dorset Nat. Hist.Archaeol. Soc. 105: 170.
Brown, D. S. 1993: A taxonomic reappraisal of thefamilies Elasmosauridae and Cryptoclididae (Reptil-ia: Plesiosauridae). — Revue de Paleobiologie Spe-cial Volume (7): 9–16.
Brown, D. S. & Cruickshank, A. R. I. 1994: The skull of
the Callovian plesiosaur Cryptoclidus eurymerus andthe sauropterygian cheek. — Palaeontology 37 (4):941–953.
Brown, D. S., Milner, A. C. & Taylor, M. A. 1986: Newmaterial of the plesiosaur Kimmerosaurus langhamiBrown from the Kimmeridge Clay of Dorset. —Bulletin of the British Museum of Natural History(geol.) 40(5): 225–234.
Cabrera, A. 1941: Un plesiosaurio neuvo del Cretáceo delChubut. — Revisita del Museo de la Plata (NeuvoSerio) 2(8): 113–130.
Caldwell, M. W. 1997a: Limb osteology and ossificationpatterns in Cryptoclidus (Reptilia: Plesiosauroidea)with a review of Sauropterygian limbs. — Journal ofVertebrate Paleontology 17(2): 295–307.
Caldwell, M. W. 1997b: Modified perichondral ossifica-tion and the evolution of paddle-like limbs in ichthy-osaurs and plesiosaurs. — Journal of VertebratePaleontology 17(3): 534–547.
Carpenter, K. 1996: A review of short-necked plesiosaursfrom the Cretaceous of the western interior, NorthAmerica. — Neues Jahrbuch Geol. Palaönt. Abh.201(2): 259–287.
Carpenter, K. 1997: Comparative cranial anatomy of twoNorth American Cretaceous plesiosaurs. — In: Call-away, J. M. & Nicholls, E. L. (eds.), Ancient marinereptiles: 191–216. Academic Press, San Diego, Cali-fornia.
Carpenter, K. 1999: Revision of North American elasmo-saurs from the Cretaceous of the western interior. —Paludicola 2(2): 148–173.
Carroll, R. T. 1988: Vertebrate paleontology and evolu-tion. — W. H. Freeman and Company, New York.
Carroll, R. T. & Gaskill, P. 1985: The nothosaur Pachyp-leurosaurus and the origin of plesiosaurs. — Philo-sophical Transactions of the Royal Society of LondonB 309: 343–393.
Chatterjee, S. & Small, B. J. 1989: New plesiosaurs from theUpper Cretaceous of Antarctica. — In: Crame, J. A.(ed.), Origins and evolution of the Antarctic biota. —Geological Society Special Publication 47: 197–215.
Chippindale, P. T. & Wiens, J. J. 1994: Weighting,partitioning, and combining characters in phylogeneticanalysis. — Systematic Biology 43(2): 278–287.
Conybeare, W. D. 1822: Additional notices on thefossil genera Ichthyosaurus and Plesiosaurus. —Transactions of the Geological Society of London2(1):103–123.
Conybeare, W. D. 1824: On the discovery of an almostperfect skeleton of the Plesiosaurus. — Transactionsof the Geological Society of London 1: 382–389.
Cope, E. D. 1869: Synopsis of the extinct Batrachia,Reptilia and Aves of North America. — Transac-tions of the American Philosophical Society (newseries) 14: 1–252.
Cragin, F. 1888: Preliminary description of a new or littleknown saurian from the Benton of Kansas. — Ameri-can Geology 2: 404–407.

O’Keefe • ACTA ZOOL. FENNICA No. 21332
Cruickshank, A. R. I. 1994a: Cranial anatomy of theLower Jurassic pliosaur Rhomaleosaurus megaceph-alus (Stutchbury) (Reptilia: Plesiosauria.). — Philo-sophical Transactions of the Royal Society of LondonB 343: 247–260.
Cruickshank, A. R. I. 1994b: A juvenile plesiosaur (Ple-siosauria: Reptilia) from the Lower Lias (Hettangian:Lower Jurassic) of Lyme Regis, England: a pliosau-roid-plesiosauroid intermediate? — Zoological Jour-nal of the Linnean Society 112: 151–178.
Cruickshank, A. R. I. 1997: A Lower Cretaceous pliosau-rid from South Africa. — Annals of the South AfricanMuseum 105(2): 207–226.
de Blainville, H. D. 1835: Description de quelques espèc-es de reptiles de la Californie, précédée de l’analysed’un système general d’Erpetologie et d’Amphibio-logie. — Nouvelles Annales du Muséum (National)d’History Naturaelle, Paris 4:233–296.
de Braga, M. & Rieppel, O. 1997: Reptile phylogeny andthe interrelationships of turtles. — Zoological Jour-nal of the Linnean Society 120: 281–354.
De la Beche, H. T. & Conybeare, W. D. 1821: Notice ofthe discovery of a new fossil animal, forming a linkbetween the Ichthyosaurus and the crocodile, togeth-er with general remarks on the osteology of Ichthyo-saurus. — Transactions of the Royal Society ofLondon 5: 559–594.
Delair, J. B. 1959: The Mesozoic reptiles of Dorset. PartII. — Proceedings of the Dorset Natural History andArchaeological Society 80: 52–90.
Druckenmiller, P. S. 1999: Osteology and relationshipsof a new plesiosaur from the Thermopolis Shale ofMontana. — Journal of Vertebrate Paleontology19(3) suppl.: 42A.
Edinger, T. 1935: Pistosaurus. — Neues Jahrbuch fürMineralogie, Geologie, und Paläontologie 74:321–359.
Fraas, E. 1910: Plesiosaurier aus dem oberen Lias vonHolzmaden. — Palaeontographica 57: 105–140.
Gray, J. E. 1825: A synopsis of the genera of reptiles andAmphibia, with a description of some new species.— Annals of Philosophy 26: 193–217.
Halstead, L. B. 1989: Plesiosaur locomotion. — Quarter-ly Journal of the Geological Society of London 146:37–40.
Hampe, O. 1992: Ein großwüchsiger Pliosauride (Reptil-ia: Plesiosauria) aus der Unterkreide (oberes Aptium)von Kolumbien. — Courier Forsch.-Inst. Sencken-berg 145: 1–32.
Kuhn, O. 1961: Die Familien der rezenten und fossilenAmphibien und Reptilien. — Verlagshaus Meisen-bach, Bamberg, Germany.
Lee, M. S. Y. 1997 Reptile relationships turn turtle. —Nature 389: 245–246.
Linder, H. 1913: Beiträge zur Kenntnis der Plesiosaurier-Gattungen Peloneustes und Pliosaurus, nebst An-hang: Über die beiden ersten Halswirbel der Plesio-saurier. — Geologie und Paläontologie Abhandlu-
gen, n. ser. 15(5): 339–409.Lingham-Soliar, T. 2000: Plesiosaur locomotion: is the
four-wing problem real or merely an atheoreticalexercise? — N. Jb. Geol. Paläont. Abh. 217(1): 45–87.
Longman, H. A. 1924: Some Queensland fossil vertebrates.— Memoirs of the Queensland Museum 8: 26–28.
Longman, H. A. 1930: Kronosaurus queenslandicus. Agiant Cretaceous pliosaur. — Memoirs of the Queens-land Museum 10: 1–7.
Lydekker, R. 1889a: On the generic position of the so-called Plesiosaurus indicus. — Geological Survey ofIndia 22(1): 49–51.
Lydekker, R. 1889b: On the remains and affinities of fivegenera of Mesozoic reptiles. — Quarterly Journal ofthe Geological Society of London 45: 41–59.
Massare, J. A. 1988: Swimming capabilities of Mesozoicmarine reptiles: implications for method of preda-tion. — Paleobiology 14(2): 187–205.
Merck, J. W. 1997: A phylogenetic analysis of theeuryapsid reptiles. — Journal of Vertebrate Paleon-tology 17(3) suppl.: 65A.
Meyer, H. v. 1839: Mitteilung an Professor Bronn gerich-tet. — Neues Jahrbuch für Mineralogie, Geognosie,Geologie und Petrefakten-Kunde 1839: 699–701.
Meyer, H. v. 1841: Thaumatosaurus oolithicus der fos-sile WunderSaurus aus dem Oolith. — Neues Jahr-buch für Mineralogie, Geologie, und Paläontologie1841: 176–184.
Meyer, H. v. 1842: Simosaurus, die Stumpfschnauze, einSaurier aus dem Muschelkalke von Luneville. —Neues Jahrbuch für Mineralogie, Geognosie, Geolo-gie und Petrefakten-Kunde 1842: 184–197.
Newman, B. & Tarlo, L. B. H. 1967: A giant marine reptilefrom Bedfordshire. — Animals, London 10(2): 61–63.
O’Keefe, F. R. 2000: Phylogeny and convergence in thePlesiosauria (Reptilia: Sauropterygia). — Ph.D. the-sis, University of Chicago, Chicago IL.
O’Keefe, F. R. 2002: The evolution and functional mor-phology of plesiosaur and pliosaur morphotypes inthe Plesiosauria (Reptilia: Sauropterygia). — Paleo-biology 28(1). [In press].
Osborn, H. F. 1903: The reptilian subclasses Diapsidaand Synapsida and the early history of the Diaptosau-ria. — Memoirs of the American Museum of NaturalHistory 1: 449–507.
Owen, R. 1838: A description of Viscount Cole’s speci-men of Plesiosaurus macrocephalus (Conybeare). —Proceedings of the Geological Society of London 2:663–666.
Owen, R. 1840: Report on British Fossil Reptiles, Part I.— Reports of the British Association for the Ad-vancement of Science, London: 42–126.
Owen, R. 1841: Report on British fossil reptiles, Part II.— Reports of the British Association for the Ad-vancement of Science, London: 60–65.
Owen, R. 1860: On the orders of fossil and RecentReptilia, and their distribution through time. — Re-port of the British Association for the Advancement

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 33
of Science 29: 153–166.Owen, R. 1865: A monograph on the fossil Reptilia of the
Liassic formations, Part 3. — Palaeontological Soci-ety of London, London.
Persson, P. O. 1963: A revision of the classification of thePlesiosauria with a synopsis of the stratigraphical andgeological distribution of the group. — Lunds Univer-sites Årsskrift N. F. Ard. 2 Bd. 59 Num. 1: 1–57.
Phillips, J. 1871: Geology of Oxford and the valley of theThames. — Oxford University Press, Oxford, England.
Reisz, R. R. 1981: A diapsid reptile from the Pennsylva-nian of Kansas. — Special Publication of the Muse-um of Natural History, University of Kansas 7: 1–74.
Rieppel, O. 1994a: Osteology of Simosaurus gaillardotiand the relationships of stem-group Sauropterygia.— Fieldiana Geology, New Series 28:1–85.
Rieppel, O. 1994b: The braincases of Simosaurus andNothosaurus: monophyly of the Nothosauridae (Rep-tilia: Sauropterygia). — Journal of Vertebrate Pale-ontology 14(1): 9–23.
Rieppel, O. 1997a: Introduction to Sauropterygia. — In:Callaway, J. M. & Nicholls, E. L. (eds.), Ancientmarine reptiles: 107–119. Academic Press, San Di-ego, California.
Rieppel, O. 1997b: Revision of the sauroptergian reptilegenus Cymatosaurus v. Fritsch, 1894, and the rela-tionships of Germanosaurus Nopcsa 1928, from theMiddle Triassic of Europe. — Fieldiana (Geology)N. S. 36: 1–38.
Rieppel, O. 1998: Corosaurus alcovensis Case and thephylogenetic interrelationships of Triassic stem-groupSauropterygia. — Zoological Journal of the LinneanSociety 124: 1–41.
Rieppel, O. 1999: Phylogeny and paleobiogeography ofTriassic Sauropterygia: problems solved and unre-solved. — Palaeogeography, Palaeoclimatology, Pal-aeoecology 153: 1–15.
Rieppel, O. 2000: Sauropterygia I. Placodontia, Pachyp-leurosauria, Nothosauroidea, Pistosauroidea. — In:Kuhn, O. & Wellnhofer, P. (eds.), Encylopedia ofpaleoherpetology, Part 12A: 1–134. Verlag Dr. Frie-drich Pfeil, München, Germany.
Rieppel, O. & deBraga, M. 1996: Turtles as diapsidreptiles. — Nature 384: 453–455.
Rieppel, O. & Hagdorn, H. 1997: Paleobiogeography ofMiddle Triassic Sauropterygia in central and westernEurope. — In: Callaway, J. M. & Nicholls, E. L.(eds.), Ancient marine reptiles: 121–144. AcademicPress, San Diego, California.
Rieppel, O. C. & Reisz, R. R. 1999: The origin and earlyevolution of turtles. — Annual Review of Ecologyand Systematics 30: 1–22.
Rieppel, O. & Werneburg 1998: A new species of thesauropterygian Cymatosaurus from the lowerMuschelkalk of Thuringia, Germany. — Palaeontol-ogy 41(4): 575–589.
Rieppel, O. & Wild, R. 1996: A revision of the genusNothosaurus (Reptilia: Sauropterygia) from the Ger-
man Triassic, with comments on the status of Con-chiosaurus clavatus. — Fieldiana (Geology) N. S.34: 1–82.
Romer, A. S. 1956: Osteology of the Reptiles. — Univer-sity of Chicago Press, Chicago, Illinois.
Romer, A. S. 1966: Vertebrate paleontology, 3rd ed. —University of Chicago Press, Chicago, Illinois.
Sander, P. M., Rieppel, O. C. & Bucher, H. 1997: A newpistosaurid (Reptilia: Sauropterygra) from the Mid-dle Triassic of Nevada and its implications for theorigins of plesiosaurs. — Journal of VertebratePaleontology 17(3): 526–533.
Sauvage, H. E. 1873: Notes sur les reptiles fossiles.— Bulletin of the Geological Society of France1(3): 365–380.
Seeley, H. 1874a: On Muraenosaurus leedsi, a plesiosau-rian from the Oxford Clay. — Quarterly Journal ofthe Geological Society, London 30: 197–208.
Seeley, H. 1874b: Note on some generic modifications ofthe plesiosaurian pectoral arch. — Quarterly Journalof the Geological Society of London 30: 436–449.
Seeley, H. 1892: The nature of the shoulder girdle andthe clavicular arch in Sauropterygia. — Proceedingsof the Royal Society of London 51: 119–151.
Sereno, P. C. 1998: A rationale for phylogenetic defini-tions, with application to the higher-level taxonomyof Dinosauria. — Neues Jahrbuch Geol. Palaönt.Abh. 210(1):41–83.
Sollas, W. J. 1881: On a new species of Plesiosaurus (P.conybeari) from the Lower Lias of Charmouth; withobservations on P. megacephalus, Stutchbury, and P.brachycephalus, Owen. — Quarterly Journal of theGeological Society of London 37: 440–480.
Storrs, G. W. 1991: Anatomy and relationships of Coro-saurus alcovensis (Diapsida: Sauropterygia) and theTriassic Alcova Limestone of Wyoming. — Bulletinof the Peabody Museum of Natural History 44: 1–151.
Storrs, G. W. 1993: Function and phylogeny in saurop-terygian (Diapsida) evolution. — American Journalof Science 293 A: 63–90.
Storrs, G. W. 1997: Morphological and taxonomic clarifi-cation of the genus Plesiosaurus. — In: Callaway, J.M. & Nicholls, E. L. (eds.), Ancient marine reptiles:145–190. Academic Press, San Diego, California.
Storrs, G. W. 1999: An examination of Plesiosauria(Diapsida: Sauropterygia) from the Niobrara Chalk(Upper Cretaceous) of central North America. —University of Kansas Paleontological Contributions11: 1–15.
Storrs, G. W. & Taylor, M. A. 1996: Cranial anatomy of anew plesiosaur genus from the lowermost Lias, (Rhae-tian/Hettangian) of Street, Somerset, England. —Journal of Vertebrate Paleontology 16(3): 403–420.
Stutchbury, S. 1846: Description of a new species ofPlesiosaurus, in the museum of the Bristol Institu-tion. — Quarterly Journal of the Geological Societyof London 2: 411–417.
Sues, H. D. 1987: Postcranial skeleton of Pistosaurus and

O’Keefe • ACTA ZOOL. FENNICA No. 21334
interrelationships of the Sauropterygia. — Zoologi-cal Journal of the Linnean Society 90: 109–131.
Swinton, W. E. 1930: Preliminary account of a newgenus and species of plesiosaur. — Annals andMagazine of Natural History 6(10): 206–209.
Swofford, D. L. & Begle, D. P. 1993: PAUP: Phyloge-netic analysis using parsimony, version 3.1.1. —Laboratory of Molecular Systematics, SmithsonianInstitution, Washington, D.C.
Tarlo, L. B. 1960: A review of the Upper Jurassicpliosaurs. — Bulletin of the British Museum (NaturalHistory) Geology 4(5): 147–189.
Taylor, M. A. 1992a: Taxonomy and taphonomy ofRhomaleosaurus zetlandicus (Plesiosauria, Reptilia)from the Toarcian (Lower Jurassic) of the YorkshireCoast. — Proceedings of the Yorkshire GeologicalSociety 49: 49–55.
Taylor, M. A. 1992b: Functional anatomy of the head ofthe large aquatic predator Rhomaleosaurus zetlandi-cus (Plesiosauria, Reptilia) from the Toarcian (LowerJurassic) of the Yorkshire Coast. — PhilosophicalTransactions of the Royal Society of London B: 335:247–280.
Taylor, M. A. 1997: Before the dinosaur: the historicalsignificance of the fossil marine reptiles. — In:Callaway, J. M. & Nicholls, E. L. (eds.), Ancientmarine reptiles: xix–xlvi. Academic Press, San Di-ego, California.
Taylor, M. A. & Cruickshank, A. R. I. 1993: Cranialanatomy and functional morphology of Pliosaurusbrachyspondylus (Reptilia: Plesiosauria) from theUpper Jurassic of Westbury, Wiltshire. — Philo-sophical Transactions of the Royal Society of LondonB 341: 399–418.
Vaughn, P. P. 1955: The Permian reptile Araeoscelisrestudied. — Bulletin of the Museum of ComparativeZoology 113(5): 305–467.
von Fritsch, K. 1894: Beitrag zur Kenntnis der Saurierdes Halle’schen unteren Muschelkalkes. — Abhand-lungen der Naturforschenden Gesellschaft zu Halle20: 273–302.
Watson, D. W. S. 1911: Upper Lias Reptilia, Part 3.Microcleidus macropterus (Seeley) and the limbs ofMicrocleidus homalospondylus (Owen). — Memoirsof the Manchester Literary and Philosophical Society55(17): 1–9.
Wegner, T. 1914: Brancasaurus brancai n. g. n. sp., einElasmosauride aus Wealden Westfalens. — In: Bran-
ca-Festschrift: 235–302. Verlag von Gebrüder Born-traeger, Leipzig, Germany.
Welles, S. P. 1943: Elasmosaurid plesiosaurs with de-scription of new material from California and Colo-rado. — Memoirs of the University of California 13:125–254.
Welles, S. P. 1949: A new elasmosaur from the EagleFord Shale of Texas. — Fondren Science Series,Southern Methodist University 1: 1–28.
Welles, S. P. 1952: A review of the North AmericanCretaceous elasmosaurs. — University of CaliforniaPublications in the Geological Sciences 29: 47–144.
Welles, S. P. 1962: A new species of elasmosaur from theAptian of Columbia and a review of the Cretaceousplesiosaurs. — University of California Publicationsin the Geological Sciences 44(1): 1–96.
White, T. E. 1935: On the skull of Kronosaurus queens-landicus Longman. — Occasional Papers of theBoston Society of Natural History 8: 219–228.
White, T. E. 1940: Holotype of Plesiosaurus longirostrusBlake and the classification of the plesiosaurs. —Journal of Paleontology 14(5): 451–467.
Williston, S. W. 1890: A new plesiosaur from the Nio-brara Cretaceous of Kansas. — Transaction of theKansas Academy of Sciences, 12: 174–178.
Williston, S. W. 1903: North American plesiosaurs, Part1. — Field Columbian Museum Publication (Geolo-gy) 73(2): 1–77.
Williston, S. W. 1906: North American plesiosaurs:Elasmosaurus, Cimoliasaurus, and Polycotylus.— American Journal of Science, Fourth Series21(123): 221–236.
Williston, S. W. 1907: The skull of Brachauchenius, withobservations of the relationships of the plesiosaurs.— Proceedings of the National Museum 32(1540):477–493.
Williston, S. W. 1908: North American plesiosaurs. Tri-nacromerum. — Journal of Geology, Chicago 16:715–736.
Williston, S. W. 1914: Water reptiles of the past andpresent. — University of Chicago Press, Chicago,Illinois.
Williston, S. W. 1925: The osteology of the reptiles. —Cambridge University Press, Cambridge, Massachu-setts.
Zittel, K. A. 1887: Handbuch der Paläontologie. I.Paläozoologie. III. Amphibien, Reptilien, Vögel. —Oldenbourg, München und Leipzig.

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 35
Ap
pen
dix
1. C
hara
cter
s, s
tate
defi
nitio
ns, r
efer
ence
s, a
nd c
omm
ents
for
the
clad
istic
cha
ract
ers
used
in th
is p
aper
. For
dis
cuss
ion
see
text
.—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—C
har.
type
Cha
ract
erS
tate
s/C
odin
gC
itatio
nR
emar
ks—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—1
Mor
phom
etric
Rel
ativ
e sk
ull l
engt
hP
rimiti
ve/ ‘
noth
osau
rian’
(0)
,B
row
n 19
81, c
har.
2S
core
d fr
om m
orph
omet
ric d
ata
in O
’Kee
fe 2
000:
larg
e (1
), s
mal
l (2)
chap
ter
3. T
hala
ssio
drac
on is
prim
itive
for
this
char
acte
r, a
s ar
e m
ost r
hom
aleo
saur
ids.
2M
orph
omet
ricR
elat
ive
neck
leng
thP
rimiti
ve (
0), l
ong
(1),
sho
rt (
2)S
torr
s 19
91, c
har.
2S
core
d fr
om m
orph
omet
ric d
ata
in O
’Kee
fe 2
000:
chap
ter
3.
3M
orph
omet
ricR
elat
ive
leng
th o
fS
ubeq
ual (
0), i
schi
um lo
nger
(1)
,B
row
n 19
81, c
har.
29
Sco
red
from
mor
phom
etric
dat
a in
O’K
eefe
200
0:is
chiu
m/p
ubis
pubi
s lo
nger
(2)
chap
ter
3.
4M
orph
omet
ricR
elat
ive
leng
th o
fS
ubeq
ual (
0), c
orac
oid
long
er, (
1)W
elle
s 19
62S
core
d fr
om m
orph
omet
ric d
ata
in O
’Kee
fe 2
000:
scap
ula/
cora
coid
chap
ter
3; d
eriv
ed e
lasm
osau
rs a
re th
e on
lypl
esio
saur
s in
whi
ch th
is r
atio
is s
ubeq
ual.
5M
orph
omet
ricR
elat
ive
leng
th o
fS
ubeq
ual (
0), h
umer
us lo
nger
(1)
,B
row
n 19
81, c
har.
32
Sco
red
from
mor
phom
etric
dat
a in
O’K
eefe
200
0:hu
mer
us/fe
mur
fem
ur lo
nger
(2)
chap
ter
3.
6M
orph
omet
ricP
reor
bita
l and
Sub
equa
l (0)
, lon
ger
preo
rbita
l (1)
,R
iepp
el 1
994a
, 199
7b,
Thi
s ra
tio is
con
trol
led
by th
e le
ngth
of t
he s
nout
post
orbi
tal s
kull
leng
thsh
orte
r (2
)ch
ars.
9, 1
2re
lativ
e to
the
tem
pora
l fen
estr
a an
d su
spen
soriu
m.
7M
orph
omet
ricF
in a
spec
t rat
ioH
igh
(0),
low
(1)
Sco
red
from
mor
phom
etric
dat
a in
O’K
eefe
200
0:ch
apte
r 3;
the
two
stat
es a
re b
ased
on
a br
eak
inth
e di
strib
utio
n of
asp
ect r
atio
dat
a.
8S
kull
shap
eE
long
ate
rost
rum
Abs
ent (
0), p
rem
axill
a on
ly (
1),
Par
tially
mod
ified
from
Sta
te o
ne is
defi
ned
as a
long
sno
ut w
ith li
ttle
or n
ove
ry lo
ng w
ith m
axill
a in
clud
ed (
2),
Tar
lo 1
960
incr
ease
in s
nout
leng
th p
oste
rior
to p
rem
axill
a/el
onga
te a
nd h
oop-
like/
unco
nstr
icte
d (3
)m
axill
a su
ture
. Sta
te tw
o in
clud
es le
ngth
enin
g of
max
illa;
sta
te th
ree
desc
ribes
the
larg
e, u
ncon
stric
ted
snou
ts o
f Kim
mer
osau
rus
and
rela
ted
taxa
. Thi
sch
arac
ter
is o
rder
ed, u
sing
the
char
acte
r st
ate
tree
(0(3
)(1,
2)).
Fig
s. 7
, 2 a
nd 1
1.
9S
kull
shap
eC
onst
ricte
d sn
out
Unc
onst
ricte
d (0
), c
onst
ricte
d at
Rie
ppel
199
4a, 1
997b
,C
onst
rictio
n at
the
prem
axill
a/m
axill
a su
ture
is c
omm
onm
axill
a/pr
emax
illa
sutu
re(1
), s
econ
dS
torr
s 19
91, c
hars
. 3, 1
3in
long
-sno
uted
sau
ropt
eryg
ians
. Sta
te tw
o de
scrib
esco
nstr
ictio
n in
max
illa
(2)
the
cond
ition
of L
iopl
euro
don ,
whi
ch h
as a
sec
ond,
mor
e po
ster
ior
cons
tric
tion
(And
rew
s 19
13).
Thi
sch
arac
ter
is o
rder
ed b
ecau
se th
e se
cond
con
stric
tion
neve
r oc
curs
in th
e ab
senc
e of
the
first
. Fig
. 11.
Con
tinue
d

O’Keefe • ACTA ZOOL. FENNICA No. 21336A
pp
end
ix 1
. Con
tinue
d.—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—C
har.
type
Cha
ract
erS
tate
s/C
odin
gC
itatio
nR
emar
ks—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—10
Sku
ll sh
ape
Tem
pora
l em
argi
natio
nP
rese
nt (
0), a
bsen
t(1)
Sto
rrs
1991
, cha
r. 6
Ara
eosc
elis
has
a s
trai
ght s
kul l
mar
gin
belo
w th
ete
mpo
ral f
enes
tra;
mos
t sau
ropt
eryg
ians
hav
e an
emar
gina
tion
here
. The
con
ditio
n in
ple
sios
aurs
isdi
fficu
lt to
det
erm
ine,
as
the
varia
tion
is c
ontin
uos;
my
crite
rion
for
the
pres
ence
of e
mar
gina
tion
is a
nex
cava
ted
vent
ro-la
tera
l em
baym
ent o
f the
squa
mos
al. B
y th
is c
riter
ion
mos
t ple
sios
aurs
hav
ete
mpo
ral e
mar
gina
tion.
Fig
. 14.
11S
kull
roof
Dor
so-m
edia
l pro
cess
Con
tact
s fr
onta
l (0)
, con
tact
s pa
rieta
lA
ndre
ws
1911
bT
he p
rem
axi l l
ae s
end
dors
o-m
edia
l pro
cess
esof
pre
max
i l la
at p
inea
l for
amen
(2),
con
tact
s an
terio
rpo
ster
iorly
acr
oss
the
skul
l roo
f in
mos
tex
tens
ion
of th
e pa
rieta
l (1)
plio
saur
omor
ph ta
xa a
nd s
ome
elas
mos
aurs
, and
thes
e pr
oces
ses
may
mee
t the
par
ieta
ls in
var
ious
way
s. W
hile
thes
e pr
oces
ses
sepa
rate
the
fron
tals
on
the
dors
al m
idlin
e, th
e fr
onta
ls m
ay r
etai
n a
mid
line
sutu
re b
enea
th th
e pr
emax
illae
(A
ndre
ws
1911
b).
Fig
s. 5
and
10.
12S
kull
roof
Pre
max
illa/
exte
rnal
Pre
sent
(0)
, abs
ent (
1)C
onta
ct o
f the
pre
max
illa
with
the
exte
rnal
nar
is is
naris
con
tact
prim
itive
, occ
urrin
g in
Ara
eosc
elis
. Thi
s co
ntac
t is
lost
in s
ome
plio
saur
s. F
ig. 1
0.
13S
kull
roof
Dor
sal m
edia
n fo
ram
enA
bsen
t (0)
, pre
sent
(1)
Cru
icks
hank
199
7A
sha
llow
dep
ress
ion
occu
rs in
the
dors
o-m
edia
lpr
oces
ses
of th
e pr
emax
illae
in s
ome
rhom
aleo
saur
ids.
14S
kull
roof
Fro
ntal
s pa
ired/
fuse
dP
aire
d (0
), fu
sed
(1)
Rie
ppel
199
4a, 1
997b
,F
usio
n of
the
fron
tals
on
the
skul
l roo
f can
onl
y be
in a
dult
char
s. 1
1, 1
4sc
ored
for
taxa
in w
hich
the
fron
tal m
idlin
e su
ture
isvi
sibl
e; th
is c
hara
cter
is th
eref
ore
inap
plic
able
for
taxa
in w
hich
the
dors
o-m
edia
l pro
cess
es o
f the
prem
axill
ae r
each
the
parie
tals
.
15S
kull
roof
Par
ieta
ls p
aire
d or
Pai
red
(0),
fuse
d po
ster
iorly
(1)
, fus
ed (
2)R
iepp
el 1
997b
, cha
r. 1
7F
usio
n of
the
parie
tals
on
the
dors
al m
idlin
e is
an
fuse
d in
adu
ltim
port
ant c
hara
cter
in b
asal
‘not
hosa
urs’
; how
ever
this
cha
ract
er is
diffi
cult
to s
core
in ta
xa w
here
the
parie
tal s
kull
tabl
e is
a n
arro
w s
agitt
al c
rest
, whi
ch is
vuln
erab
le to
bre
akag
e. I
have
ther
efor
e sc
ored
this
char
acte
r as
inap
plic
able
for
ples
iosa
urs.
16S
kll
fF
tl
ithith
tW
itht
(0)
Ri
l199
419
97b
Adi
t it
tl
tl
f th
ft
l

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 3716
Sku
ll ro
ofF
ront
al w
ith o
r w
ithou
tW
ithou
t pro
cess
es (
0),
Rie
ppel
199
4a, 1
997b
,A
dis
tinct
pos
tero
-late
ral p
roce
ss o
f the
fron
tal o
ccur
sdi
stin
ct p
oste
ro-la
tera
lw
ith p
roce
sses
(1)
char
s. 1
2, 1
5; S
torr
s 19
91in
the
pist
osau
roid
s C
ymat
osau
rus
and
Pis
tosa
urus
. It
proc
ess
char
. 30
also
occ
urs
in v
ery
prim
itive
ple
sios
aurs
suc
h as
Tha
lass
iodr
acon
and
Ple
sios
auru
s . F
ig. 4
.
17S
kull
roof
Pos
torb
ital b
arB
oth
po a
nd p
of h
ave
orbi
tal c
onta
ct (
0),
In s
ome
elas
mos
aurs
, the
pos
tfron
tal i
s co
nfine
d to
fron
tal/p
o su
ture
exc
lude
s po
f fro
mth
e in
side
of t
he te
mpo
ral f
enes
tra
and
excl
uded
from
orbi
t mar
gin
(1)
the
orbi
t by
a fr
onta
l-pos
torb
ital s
utur
e.
18S
kull
roof
Fro
ntal
ent
ers
mar
gin
Doe
s no
t (0)
,R
iepp
el 1
994a
, 199
7b,
of te
mpo
ral f
enes
tra
does
nar
row
ly (
1)ch
ars.
13,
16
19S
kull
roof
Fro
ntal
con
tact
sD
oes
cont
act (
0),
The
fron
tal c
onta
cts
the
exte
rnal
nar
is in
som
eex
tern
al n
aris
does
not
con
tact
(1)
plio
saur
s. F
ig. 1
0.
20S
kull
roof
Pin
eal f
oram
en lo
catio
nM
iddl
e of
par
ieta
l (0)
, ant
erio
r (1
)R
iepp
el 1
994a
, 199
7b,
The
mov
emen
t of t
he p
inea
l for
amen
to th
e an
terio
r of
char
s. 1
5, 1
8, a
lso
the
parie
tal s
kul l
tabl
e is
a c
hara
cter
sha
red
by th
eS
ues
1987
pist
osau
rids
and
ples
iosa
urs.
Som
e w
orke
rs (
Wel
les
1962
, Car
pent
er 1
996)
con
tend
that
the
pine
alfo
ram
en is
lost
in s
ome
taxa
; It i
s pr
esen
t in
all t
axa
Iha
ve e
xam
ined
. Fig
s. 4
and
5.
21S
kull
roof
Pin
eal f
oram
enN
ot b
orde
red
by fr
onta
l (0)
,A
ndre
ws
1911
bIn
Pel
oneu
stes
(an
d po
ssib
ly o
ther
plio
saur
s kn
own
bord
ered
ant
erio
rlybo
rder
ed b
y fr
onta
l (1)
from
infe
rior
mat
eria
l), th
e fr
onta
ls fo
rm p
art o
f the
by fr
onta
ls o
n do
rsal
ante
rior
mar
gin
of th
e pi
neal
fora
men
, but
onl
ysk
ull s
urfa
cebe
neat
h th
e pr
emax
illae
. Tax
a w
ere
scor
ed a
s de
rived
for
this
cha
ract
er o
nly
if th
e fr
onta
l for
med
par
t of t
hem
argi
n on
the
dors
al s
kull
roof
, as
is th
e ca
se in
som
eel
asm
osau
rs a
nd c
rypt
oclid
ids,
but
not
in p
liosa
urs.
22S
kull
roof
Pre
fron
tal p
rese
ntP
rese
nt (
0), a
bsen
t (1)
or a
bsen
t
23S
kull
roof
Acc
esso
ry fe
nest
raA
bsen
t (0)
, pre
sent
(1)
Car
pent
er 1
996
Car
pent
er d
escr
ibed
this
feat
ure
in th
e po
lyco
tylid
sab
ove
orbi
tsD
olic
hory
ncho
ps a
nd T
rinac
rom
erum
; I w
as u
nabl
e to
confi
rm it
s pr
esen
ce in
eith
er ta
xon.
The
orb
ital r
egio
nin
pol
ycot
ylid
s is
del
icat
e an
d lia
ble
to b
reak
age.
24S
kull
roof
Fro
ntal
pro
cess
Abs
ent (
0), p
rese
nt (
1)C
arpe
nter
199
6C
arpe
nter
des
crib
ed th
is fe
atur
e in
the
poly
coty
lids
proj
ects
into
orb
itD
olic
hory
ncho
ps a
nd T
rinac
rom
erum
; It i
s pr
esen
t in
both
taxa
alth
ough
pre
serv
atio
n of
the
feat
ure
is p
oor
due
to th
e de
licat
e na
ture
of t
he b
ones
in th
e or
bit r
egio
n.
Con
tinue
d
O’Keefe • ACTA ZOOL. FENNICA No. 21338A
pp
end
ix 1
. Con
tinue
d.—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—C
har.
type
Cha
ract
erS
tate
s/C
odin
gC
itatio
nR
emar
ks—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—25
Sku
ll ro
ofP
arie
tal s
kul l
tabl
eR
elat
ivel
y br
oad
(0),
con
stric
ted
(1),
Rie
ppel
199
4a, 1
997b
,P
osse
ssio
n of
a s
harp
ly k
eele
d sa
gitta
l cre
st is
asa
gitta
l cre
st (
2)ch
ars.
16,
19;
feat
ure
shar
ed b
y al
l ple
sios
aurs
and
Pis
tosa
urus
. The
Bro
wn
1981
, cha
r. 9
sagi
ttal c
rest
is th
e cu
lmin
atio
n of
a tr
end
of s
kul l
tabl
ena
rrow
ing
in th
e pi
stos
auro
ids.
Fig
s. 1
4 an
d 18
.
26S
kull
roof
Squ
amos
al p
rodu
ces
No
med
ial p
roce
ss (
0), m
edia
lT
he s
utur
e of
the
quad
rate
with
the
squa
mos
al is
long
, thi
n pr
oces
spr
oces
s an
d so
cket
-l ike
squ
amos
al (
1)pr
omin
ent a
nd r
obus
t in
all p
lesi
osau
rs. I
n so
me
cove
ring
quad
rate
plio
saur
s th
e sq
uam
osal
has
des
cend
ing
late
ral a
ndla
tera
l lym
edia
l pro
cess
es o
n ei
ther
sid
e of
the
quad
rate
inoc
cipi
tal v
iew
, for
min
g a
sock
et th
at a
ccep
ts th
equ
adra
te. I
n so
me
ples
iosa
uroi
ds th
e la
tera
l pro
cess
is m
uch
long
er th
an th
e m
edia
l pro
cess
, the
late
ral
proc
ess
reac
hes
alm
ost t
o th
e ja
w a
rtic
ulat
ion,
and
the
sutu
re is
less
soc
ket-
like.
The
pol
ycot
ylid
s po
sses
sth
e se
cond
type
of s
utur
e.
27S
kull
roof
Squ
amos
al d
orsa
lN
o su
ture
(0)
, mee
t in
arch
at m
idlin
e (1
)S
ues
1987
,A
n oc
cipi
tal a
rch
form
ed b
y a
med
ian
squa
mos
alpr
oces
ses
Rie
ppel
199
7b, c
har.
36
sutu
re is
cha
ract
eris
tic o
f all
ples
iosa
urs
and
Pis
tosa
urus
. Cym
atos
auru
s is
inte
rmed
iate
in h
avin
g a
very
nar
row
sep
arat
ion
of th
e sq
uam
osal
s in
occ
ipita
lvi
ew (
see
Rie
ppel
199
8b).
Fig
s. 5
and
10.
28S
kull
roof
Squ
amos
al/p
osto
rbita
lC
onta
ct (
0), n
o co
ntac
t (1)
The
squ
amos
al o
ften
cont
acts
the
post
orbi
tal w
ithin
cont
act
the
tem
pora
l fen
estr
a on
the
rear
of t
he p
osto
rbita
lba
r; h
owev
er, t
his
char
acte
r co
ncer
ns o
nly
the
mar
gin
of th
e te
mpo
ral f
enes
tra
in la
tera
l vie
w. C
onta
ct o
f the
two
bone
s ex
clud
es th
e ju
gal f
rom
the
mar
gin
of th
ete
mpo
ral f
enes
tra.
Fig
. 14.
29S
kull
roof
Juga
l/squ
amos
alN
o co
ntac
t (0)
, con
tact
(1)
Con
tact
of t
he ju
gal a
nd s
quam
osal
is c
hara
cter
istic
of
cont
act
all p
lesi
osau
rs a
nd C
ymat
osau
rus.
Fig
. 14.
30S
kull
roof
Juga
l ext
ends
Ant
erio
r m
argi
n (0
), m
iddl
e of
orb
it (1
),R
iepp
el 1
994a
, 199
7b,
Fig
s. 4
, 5 a
nd 1
0.an
terio
rly a
long
ven
tral
rest
ricte
d to
pos
terio
r m
argi
n (2
)ch
ars.
20,
23;
Sto
rrs
1991
,or
bita
l mar
gin
char
. 24;
Cru
icks
hank
1994
b, c
har.
2
31S
kull
roof
Juga
l con
tact
sC
onta
cts
orbi
t (0)
, exc
lude
dM
odifi
ed fr
om S
torr
s 19
91,
orbi
t mar
gin
by p
o/m
con
tact
(1)
char
. 24
32S
kull
roof
Juga
l/pre
fron
tals
utur
eA
bsen
t(0)
pres
ent(
1)M
any
plio
saur
sha
vea
sutu
rebe
twee
nth
epr
efro
ntal

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 39
32S
kull
roof
Juga
l/pre
fron
tal s
utur
eA
bsen
t (0)
, pre
sent
(1)
Man
y pl
iosa
urs
have
a s
utur
e be
twee
n th
e pr
efro
ntal
ante
rior
to o
rbit
and
juga
l ant
erio
r to
the
orbi
t, ex
clud
ing
the
max
illa
from
the
orbi
t mar
gin.
The
juga
l ext
ends
forw
ard
and
med
ially
aro
und
the
orbi
t to
mee
t the
pre
fron
tal.
The
pres
ence
of t
he ju
gal i
n th
is p
ositi
on h
as b
een
inte
rpre
ted
as a
lacr
imal
by
And
rew
s (1
913)
, and
Tay
lor
and
Cru
icks
hank
(19
93);
how
ever
the
lacr
imal
is lo
st in
bas
al s
auro
pter
ygia
ns. F
ig. 1
0.
33S
kull
roof
Juga
l for
ms
narr
ow b
arD
oes
not (
0), d
oes
(1)
Cru
icks
hank
199
4b,
As
reco
nstr
ucte
d by
Bro
wn
and
Cru
icks
hank
(19
94),
betw
een
orbi
t and
char
. 2an
d B
row
n (1
981)
, the
che
ek r
egio
n in
cry
ptoc
l idid
s is
tem
pora
l em
argi
natio
nre
duce
d, s
o th
at th
e ju
gal i
s th
e so
le e
lem
ent b
etw
een
the
tem
pora
l em
argi
natio
n an
d th
e or
bit.
Thi
s ar
ea is
very
frag
i le a
nd g
ener
ally
not
pre
serv
ed in
the
cryp
tocl
idid
s. T
he a
pom
orph
ic c
ondi
tion
of th
e ch
eek
in p
olyc
otyl
ids
is v
ery
sim
ilar
to th
is c
ondi
tion.
Fig
. 18.
34S
kull
roof
Qua
drat
ojug
alP
rese
nt (
0), a
bsen
t (1)
Rie
ppel
199
4a,
1997
b, c
hars
. 24,
29;
Sto
rrs
1991
, cha
r. 7
;S
ues
1987
35S
kull
roof
Nas
alN
ot r
educ
ed (
0), r
educ
ed (
1), a
bsen
t (2)
Rie
ppel
199
7b, c
har.
6;
Sto
rrs
(199
1) b
elie
ved
the
nasa
l los
t in
all p
lesi
osau
rs;
Sto
rrs
1991
, cha
r. 3
4;ho
wev
er it
is s
mal
l but
pre
sent
in P
elon
eust
es a
ndS
ues
1987
othe
r pl
iosa
urs,
as
wel
l as
Tha
lass
iodr
acon
. Fig
s. 4
and
10.
36S
kull
roof
Loca
tion
of n
asal
Beh
ind
exte
rnal
nar
is (
0), i
n fr
ont o
fT
his
char
acte
r is
app
licab
le o
nly
to ta
xa in
whi
ch th
ere
lativ
e to
ext
erna
lex
tern
al n
aris
(1)
nasa
ls a
re p
rese
nt. T
he d
eriv
ed s
tate
occ
urs
inna
risst
ratig
raph
ical
ly la
te p
liosa
urs
such
as
Bra
chau
chen
ius.
37S
kull
roof
Dis
tinct
gro
oves
Abs
ent (
0), p
rese
nt (
1)D
istin
ct g
roov
es in
the
max
illae
ant
erio
r to
and
lead
ing
ante
rior
to e
xter
nal
into
the
exte
rnal
nar
is a
re p
rese
nt in
som
ena
risrh
omal
eosa
urid
s an
d M
acro
plat
a .
38S
kull
roof
Nas
al e
nter
s m
argi
nD
o en
ter
(0),
do
not e
nter
(1)
Rie
ppel
199
7b, c
har.
7;
The
nas
als
are
redu
ced
and
do n
ot e
nter
the
exte
rnal
of e
xter
nal n
aris
Sto
rrs
1991
, cha
r. 3
6;na
ris in
Pis
tosa
urus
, and
ther
e is
som
e qu
estio
nS
ues
1987
whe
ther
the
nasa
ls a
re p
rese
nt in
this
taxo
n at
all
(Rie
ppel
199
4a).
Whe
n pr
esen
t, th
e na
sals
do
ente
rth
e m
argi
n of
the
exte
rnal
nar
is in
ple
sios
aurs
. Fig
s.4
and
10.
Con
tinue
d

O’Keefe • ACTA ZOOL. FENNICA No. 21340A
pp
end
ix 1
. Con
tinue
d.—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—C
har.
type
Cha
ract
erS
tate
s/C
odin
gC
itatio
nR
emar
ks—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—39
Sku
ll ro
ofP
refr
onta
l con
tact
sN
o co
ntac
t (0)
, con
tact
(1)
The
pre
fron
tal r
uns
from
the
ante
rior
mar
gin
of th
em
argi
n of
ext
erna
lor
bit t
o th
e po
ster
ior
mar
gin
of th
e ex
tern
al n
aris
inna
risP
lesi
osau
rus ,
ela
smos
aurid
s, c
rypt
ocl id
ids,
and
the
poly
coty
l ids.
Fig
s. 5
and
18.
40S
kull
roof
Pre
fron
tal a
ndF
ront
al e
xclu
ded
(0),
fron
tal e
nter
sR
iepp
el 1
997b
, cha
r. 1
0F
ig 1
0.po
stfr
onta
l exc
lude
mar
gin
(1)
fron
tal f
rom
dor
sal o
rbit
mar
gin
41S
kull
roof
Max
i l la/
squ
amos
alN
o co
ntac
t (0)
, con
tact
(1)
, exp
ande
dW
il lis
ton
1903
, p. 1
9T
his
char
acte
r de
scrib
es th
e co
ntac
t, or
lack
of
cont
act
post
erio
r fla
nge
(2)
cont
act,
betw
een
the
post
erio
r en
d of
the
max
i l la
and
the
squa
mos
al. S
tate
two
desc
ribes
the
cond
ition
inpo
lyco
tylid
s, in
whi
ch th
e m
axill
a fo
rms
a st
out,
expa
nded
con
tact
with
the
squa
mos
al.
42O
ccip
utE
xocc
ipita
l par
ticip
ates
Do
not p
artic
ipat
e (0
), d
o pa
rtic
ipat
e (1
)B
row
n 19
81, c
har.
7;
in fo
rmat
ion
of o
ccip
ital
Cru
icks
hank
199
4bco
ndyl
e
43O
ccip
utO
ccip
ital c
ondy
leH
emis
pher
ical
with
gro
ove
(0),
sho
rt w
ithB
row
n 19
81, c
har.
12
Bro
wn
(198
1) s
tres
ses
the
pres
ence
of a
gro
ove
mor
phol
ogy
no g
roov
e (1
)be
twee
n th
e oc
cipi
tal c
ondy
le a
nd th
e bo
dy o
f the
basi
occi
pita
l in
his
defin
ition
of t
his
char
acte
r. T
hegr
oove
is v
aria
bly
pres
ent a
nd p
roba
bly
varie
son
toge
netic
ally
; I h
ave
foun
d th
e sh
ape
of th
e co
ndyl
eto
be
mor
e re
liabl
e in
defi
ning
sta
tes.
Fig
. 11.
44O
ccip
utP
arao
ccip
ital
No
proc
ess/
occ
iput
pla
te-li
ke (
0),
Rie
ppel
199
7b, c
har.
31
A p
late
-like
occ
iput
is p
rese
nt in
mos
t bas
alpr
oces
s/ fo
rmat
ion
ofro
d-sh
aped
pro
cess
(1)
saur
opte
rygi
ans;
how
ever
, an
open
occ
iput
with
apo
stte
mpo
ral f
ossa
rod-
shap
ed p
arao
ccip
ital p
roce
ss o
ccur
s in
the
mar
gin
pist
osau
roid
s an
d al
l ple
sios
aurs
.
45O
ccip
utD
istin
ct s
quam
osal
With
out n
otch
(0),
with
not
ch (
1)R
iepp
el 1
997b
, cha
r. 3
2no
tch
for
artic
ulat
ion
ofpa
raoc
cipi
tal p
roce
ss
46O
ccip
utP
arao
ccip
ital p
roce
ssG
raci
le (
0), r
obus
t (1)
Bro
wn
1981
, cha
r. 1
1M
odifi
ed fr
om B
row
n (1
981)
. My
‘gra
cile
’ is
a co
mbi
natio
nm
orph
olog
yof
Bro
wn’
s ‘lo
ng’ a
nd ‘t
hin’
sta
tes,
whi
le m
y ‘ro
bust
’is
a c
ombi
natio
n of
his
‘sho
rt’ a
nd ‘t
hick
’ sta
tes.
The
para
occi
pita
l pro
cess
is r
obus
t in
true
plio
saur
s.

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 41
para
occi
pita
l pro
cess
is r
obus
t in
true
plio
saur
s.
47O
ccip
utP
arao
ccip
ital p
roce
ssS
quam
osal
exc
lusi
vely
(0)
, qua
drat
eLi
bone
ctes
has
sta
te tw
o, w
hile
the
artic
ulat
ion
of th
ear
ticul
atio
nex
clus
ivel
y (1
), b
oth
quad
rate
and
para
occi
pita
l pro
cess
shi
fts to
the
quad
rate
in s
ome
squa
mos
al (
2)cr
ypto
clid
ids
and
all p
olyc
otyl
ids.
48O
ccip
utV
entr
al e
xten
t of
Doe
s no
t ext
end
vent
ral t
o oc
cipi
tal
And
rew
s 19
10T
ricle
idus
and
the
poly
coty
l ids
have
long
par
aocc
ipita
lpa
raoc
cipi
tal p
roce
ssco
ndyl
e (0
), e
xten
ds p
ast c
ondy
le (
1)pr
oces
ses
whi
ch e
xten
d ve
ntra
l of t
he o
ccip
ital
cond
yle.
49O
ccip
utN
atur
e of
par
aocc
ipita
lN
o co
ntac
t (0)
, con
tact
at l
ater
alT
aylo
r an
d C
ruic
ksha
nk (
1993
) an
d T
aylo
r (1
992a
)pr
oces
s/ q
uadr
ate
artic
ulat
iono
nly
(1),
long
con
tact
alo
ngil l
ustr
ate
the
occi
put o
f Plio
saur
us a
ndpt
eryg
oid
flang
ebo
dies
of p
roce
sses
(2)
Rho
mal
eosa
urus
res
pect
ivel
y, d
epic
ting
a lo
ngco
ntac
tco
ntac
t (st
ate
two)
bet
wee
n th
e pa
raoc
cipi
tal p
roce
ssan
d th
e qu
adra
te fl
ange
of t
he p
tery
goid
in b
oth
taxa
. Idi
d no
t obs
erve
this
con
ditio
n; th
ese
taxa
pos
sess
stat
e on
e, a
s i l l
ustr
ated
by
Cru
icks
hank
(19
94a)
.
50O
ccip
utQ
uadr
ate
flang
e of
Qua
drat
e on
ly (
0), q
uadr
ate
and
Mod
ified
from
Bro
wn
The
squ
amos
al s
hare
s in
the
quad
rate
art
icul
atio
n of
pter
ygoi
d/ q
uadr
ate
squa
mos
al (
1)19
81, c
har.
10;
the
quad
rate
flan
ge o
f the
pte
rygo
id in
som
ear
ticul
atio
nC
ruic
ksha
nk 1
994b
,el
asm
osau
rs a
nd s
ome
plio
saur
s.ch
ar. 3
51O
ccip
utQ
uadr
ate
prod
uces
Pro
cess
abs
ent (
0), p
roce
ss p
rese
nt (
1)di
stin
ct p
roce
ss fo
rar
ticul
atio
n w
ithpt
eryg
oid
flang
e
52B
rain
case
Dor
sal w
ing
ofB
road
/ col
umna
r (0
), r
educ
ed (
1)R
iepp
el 1
997b
, cha
r. 3
9ep
ipte
rygo
id
53B
rain
case
Epi
pter
ygoi
d do
rsal
Con
tact
(0)
, no
cont
act (
1)Z
anon
unp
ublis
hed
proc
ess
cont
acts
parie
tal
54O
ccip
utQ
uadr
ate
emba
yed/
Mas
sive
qua
drat
e (0
), d
ishe
dM
ost p
lesi
osau
rs h
ave
a di
shed
or
emba
yed
ante
rior
dish
ed-s
hape
dan
terio
rly (
1)m
argi
n of
the
quad
rate
abo
ve th
e ja
w a
rtic
ulat
ion.
Thi
san
terio
rlyem
baym
ent i
s lo
st in
som
e el
asm
osau
rs.
55S
kull
roof
Pos
terio
r bu
lb fo
rmed
Abs
ent (
0), p
rese
nt (
1)T
he a
pex
of th
e sq
uam
osal
arc
h is
ela
bora
ted
into
aby
squ
amos
als
post
erio
rly-d
irect
ed b
ulb
in m
ost p
liosa
urs.
Fig
. 10.
Con
tinue
d

O’Keefe • ACTA ZOOL. FENNICA No. 21342A
pp
end
ix 1
. Con
tinue
d.—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—C
har.
type
Cha
ract
erS
tate
s/C
odin
gC
itatio
nR
emar
ks—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—56
Bra
inca
seS
upra
occi
pita
lR
ound
(0)
, med
ian
proc
ess
(1)
Zan
on u
npub
lishe
dT
he s
upra
occi
pita
l has
a m
edia
n pr
oces
s pr
ojec
ting
mor
phol
ogy
vent
ral ly
into
the
fora
men
mag
num
in m
ost
ples
iosa
urs.
Som
e ta
xa la
ck th
is fe
atur
e, a
lthou
gh it
isal
so d
elic
ate
and
l iabl
e to
bre
akag
e. T
hesu
prao
ccip
ital i
n ge
nera
l is
not b
ound
tigh
tly to
the
rest
of t
he s
kul l
and
is o
ften
lost
, at l
east
in L
ower
Jura
ssic
taxa
. Fig
. 5.
57P
alat
eS
hape
of t
he q
uadr
ate
Cur
ved
with
rai
sed
late
ral m
argi
n (0
),T
he q
uadr
ate
flang
e of
the
pter
ygoi
d is
a la
rge,
pter
ygoi
d fla
nge
stra
ight
and
nar
row
ing
(1),
sig
moi
d w
ithsi
gmoi
d st
ruct
ure
in m
any
plio
saur
s. T
his
cond
ition
isro
l led
late
ral m
argi
n (2
)de
rived
com
pare
d to
Tha
lass
iodr
acon
.A
ttenb
oros
auru
s di
spla
ys a
fairl
y in
term
edia
teco
nditi
on. F
ig. 1
1.
58P
alat
eS
quar
ed la
ppet
of
No
squa
red
lapp
et (
0), s
quar
ed la
ppet
(1)
Thi
s ch
arac
ter
desc
ribes
the
elab
orat
ion
of a
sm
all
pter
ygoi
d un
derli
esfla
nge
or la
ppet
from
the
body
of t
he p
tery
goid
, whi
chqu
adra
te p
tery
goid
unde
rlies
the
quad
rate
flan
ge o
f the
pte
rygo
id a
t its
flang
eor
igin
atio
n fr
om th
e pa
lata
l por
tion
of th
at b
one.
59B
rain
case
Sup
raoc
cipi
tal d
epth
/S
hallo
w (
0), d
eep
ante
ro-p
oste
riorly
/T
he s
upra
occi
pita
l in
Ple
sios
auru
s an
d ot
her
prim
itive
sigm
oid
sutu
resi
gmoi
d su
ture
with
exo
ccip
ital a
ndta
xa is
sha
llow
ant
ero-
post
erio
rly; t
he s
utur
e w
ith th
epr
ootic
(1)
exoc
cipi
tal i
s sh
ort a
nd th
e la
tera
l con
tact
with
the
proo
tic is
sm
all.
In o
ther
ple
sios
aurs
the
supr
aocc
ipita
lte
nds
to b
ecom
e de
eper
, and
the
sutu
re w
ith th
eex
occi
pita
l and
pro
otic
bec
omes
long
er, m
ore
late
ral,
and
sigm
oid
in s
hape
. The
spa
ce fo
r th
e ve
rtic
alse
mic
ircul
ar c
anal
is u
sual
ly la
rger
and
dee
per
in th
ese
cond
type
of s
upra
occi
pita
l. F
ig. 6
.
60P
alat
eA
nter
ior
inte
rpte
rygo
idA
bsen
t (0)
, slit
-like
(1)
, bro
ad w
ith r
ound
The
pre
senc
e of
an
ante
rior
inte
rpte
rygo
id v
acui
ty is
ava
cuity
ends
(2)
reve
rsal
with
res
pect
to m
ore
basa
l sau
ropt
eryg
ians
,w
hich
hav
e a
clos
ed p
alat
e. F
igs.
4, 5
, etc
.
61P
alat
eP
oste
rior
inte
rpte
rygo
idA
bsen
t (0)
, pre
sent
(1)
Sto
rrs
1991
, cha
r. 1
1;T
he p
rese
nce
of a
n po
ster
ior
inte
rpte
rygo
id v
acui
ty is
vacu
ityS
ues
1987
a re
vers
al w
ith r
espe
ct to
mor
e ba
sal s
auro
pter
ygia
ns,
whi
ch h
ave
a cl
osed
pal
ate.
Pre
sent
in a
ll pl
esio
saur
san
d P
isto
saur
us. F
igs.
6, 7
, 8, e
tc.

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 4362
Pal
ate
Pte
rygo
ids
mee
tP
tery
goid
s do
not
mee
t (0)
, pte
rygo
ids
Will
isto
n 19
08 il
lust
rate
sIn
Pis
tosa
urus
and
som
e Lo
wer
Jur
assi
c pl
esio
saur
spo
ster
ior
to p
oste
rior
mee
t (1)
, mee
t but
are
cov
ered
by
stat
e tw
oth
e pt
eryg
oids
do
not m
eet o
n th
e m
idlin
e of
the
pala
tein
terp
tery
goid
vac
uity
post
erio
r pa
rasp
heno
id p
roce
ss (
2)su
rfac
e be
hind
the
post
erio
r in
terp
tery
goid
vac
uity
.M
ost l
ater
ple
sios
aurs
hav
e di
stin
ct p
tery
goid
proc
esse
s w
hich
mee
t on
the
mid
l ine
behi
nd th
eva
cuity
. Thi
s ch
arac
ter
is o
rder
ed b
ecau
se s
tate
two
isde
pend
ent o
n th
e po
sses
sion
of s
tate
one
. Fig
s. 6
,11
and
17.
63P
alat
eP
tery
goid
s m
eet
Do
not m
eet b
etw
een
vacu
ities
(0)
, do
In p
lesi
osau
rs p
osse
ssin
g bo
th a
nter
ior
and
post
erio
rbe
twee
n an
terio
r an
dm
eet b
etw
een
vacu
ities
(1)
inte
rpte
rygo
id v
acui
ties,
the
pter
ygoi
ds m
ay o
r m
aypo
ster
ior
inte
rpte
rygo
idno
t mee
t bet
wee
n th
e tw
o va
cuiti
es. I
n ta
xa in
whi
chva
cuiti
esth
e pt
eryg
oids
do
not m
eet,
the
two
vacu
ities
are
sepa
rate
d by
the
cultr
iform
pro
cess
of t
hepa
rasp
heno
id. F
igs.
4, 6
, 7, e
tc.
64P
alat
eB
asio
ccip
ital e
xpos
edE
xpos
ed (
0), n
ot e
xpos
ed (
1)T
his
char
acte
r is
con
ditio
nal o
n th
e pr
esen
ce o
f apo
ster
ior
to p
oste
rior
deriv
ed s
tate
for
char
acte
r 62
. The
med
ial p
roce
sses
pter
ygoi
d su
ture
of th
e pt
eryg
oid
form
ing
the
med
ial s
ymph
ysis
beh
ind
the
post
erio
r in
terp
tery
goid
vac
uity
eith
er c
ompl
etel
yco
ver
the
body
of t
he b
asio
ccip
ital,
or fa
il to
cov
er it
.F
igs.
7 a
nd 8
.
65P
alat
eE
ctop
tery
goid
rea
ches
Doe
s no
t (0)
, doe
s (1
)In
Pel
oneu
stes
, the
ect
opte
rygo
id e
xten
ds m
edia
lly to
med
ially
to la
tera
lne
ar th
e la
tera
l mar
gin
of th
e po
ster
ior
inte
rpte
rygo
idm
argi
n of
pos
terio
rva
cuity
, ove
rlapp
ing
the
pter
ygoi
d. F
ig. 1
1.in
terp
tery
goid
vac
uitie
s
66P
alat
eC
olum
nar
ecto
pter
ygoi
dN
o co
ntac
t (0)
, con
tact
(1)
Illus
trat
ed b
y C
arpe
nter
,T
he e
ctop
tery
goid
in m
any
ples
iosa
uroi
d ta
xa r
ises
cont
acts
pos
torb
ital b
ar19
97do
rsal
ly o
ut o
f the
pla
ne o
f the
pal
ate,
and
con
tact
sth
e ve
ntro
-med
ial m
argi
n of
the
post
orbi
tal b
ar. T
his
char
acte
r is
sha
red
by e
lasm
osau
rs, c
rypt
oclid
ids,
and
poly
coty
lids.
Fig
s. 1
6 an
d 17
.
67pa
late
Dis
hed
pter
ygoi
dsA
bsen
t (0)
, pre
sent
(1)
In p
olyc
otyl
ids,
the
pter
ygoi
ds a
nter
o-la
tera
l and
late
ral t
o th
e po
ster
ior
inte
rpte
rygo
id v
acui
ties
are
broa
dly
dish
ed o
r de
pres
sed
shal
low
ly a
long
thei
rle
ngth
. Thi
s ch
arac
ter
was
obs
erve
d in
Dol
icho
rhyn
chop
s, T
rinac
rom
erum
, and
MO
R 7
51,
and
is n
ot a
pre
serv
atio
nal a
rtifa
ct. F
ig. 1
7.
Con
tinue
d

O’Keefe • ACTA ZOOL. FENNICA No. 21344A
pp
end
ix 1
. Con
tinue
d.—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—C
har.
type
Cha
ract
erS
tate
s/C
odin
gC
itatio
nR
emar
ks—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—68
Pal
ate
Pos
terio
r pt
eryg
oid/
Abs
ent (
0), p
rese
nt (
1)Il l
ustr
ated
by
And
rew
s,In
Tric
leid
us a
nd th
e po
lyco
tyl id
s, th
e pt
eryg
oids
sen
dpa
rasp
heno
id c
onta
ct19
10ou
t dis
tinct
med
ial p
roce
sses
that
con
tact
the
post
erio
rm
argi
n of
the
para
sphe
noid
. The
se p
roce
sses
mee
t in
a m
edia
n su
ture
in th
e po
lyco
tyl id
s, b
ut I
do n
otbe
lieve
this
con
ditio
n is
hom
olog
ous
to th
e co
nditi
on in
char
acte
r 62
. Fig
s. 1
6 an
d 17
.
69P
alat
eE
ctop
tery
goid
and
Do
not f
orm
flan
ges
(0),
form
flan
ges
(1),
Sta
te tw
o fig
ured
by
The
form
atio
n of
acc
esso
ry fl
ange
s ar
ound
the
pter
ygoi
d fo
rm la
tera
lfla
nges
mee
t in
shor
t, di
shed
con
tact
at
Whi
te 1
935
post
erio
r in
tern
pte
rygo
id v
acui
ty is
cha
ract
eris
tic o
ffla
nges
ven
tro-
late
ral
mid
l ine
(2),
mee
t in
broa
d co
ntac
t (3)
alm
ost a
l l pl
iosa
urs
excl
usiv
e of
the
rhom
aleo
saur
ids.
to p
oste
rior
pter
ygoi
dS
tate
one
occ
urs
in A
ttenb
oros
auru
s. S
tate
two
occu
rsva
cuity
in P
elon
eust
es a
nd K
rono
saur
us; s
tate
thre
e oc
curs
inLi
ople
urod
on, P
liosa
urus
, and
Bra
chau
chen
ius.
Fig
s. 1
1 an
d 13
.
70B
rain
case
Par
asph
enoi
dLo
ng, t
aper
ing
ante
riorly
(0)
, sho
rt a
ndA
ndre
ws
1910
p. 8
1–89
;T
he a
nter
ior
port
ion
of th
e pa
rasp
heno
id r
uns
mor
phol
ogy
blun
t(1)
Will
isto
n 19
03 p
.218
ante
riorly
in a
long
, tap
erin
g cu
ltrifo
rm p
roce
ss in
Ara
eosc
elis
, oth
er p
rimiti
ve d
iaps
ids,
and
Ple
sios
auru
s . In
the
cryp
tocl
idid
s an
d po
lyco
tylid
s, th
ean
terio
r po
rtio
n of
the
para
sphe
noid
term
inat
es in
ash
ort,
blun
t pro
cess
with
pro
min
ent l
ater
alar
ticul
atio
ns fo
r th
e pt
eryg
oids
loca
ted
at e
ither
sid
eon
the
pala
te s
urfa
ce. F
igs.
6, 1
6, a
nd 1
7.
71P
alat
eP
aras
phen
oid
keel
Not
kee
led
(0),
sha
rp k
eel (
1), k
eele
dT
he v
entr
al s
urfa
ce o
f the
par
asph
enoi
d, v
isib
lean
terio
rly (
2)th
roug
h th
e po
ster
ior
inte
rpte
rygo
id v
acui
ties,
has
ash
arp
keel
in v
ario
us ta
xa. C
arpe
nter
(19
97)
illus
trat
esst
ate
one
in L
ibon
ecte
s. P
elon
eust
es a
nd o
ther
plio
saur
s po
sses
s st
ate
two.
Fig
s. 7
and
11.
72P
alat
eP
aras
phen
oid
Ant
erio
r pa
rasp
heno
id n
ot e
xpos
ed o
nT
he a
nter
ior
port
ion
of th
e pa
rasp
heno
id is
eith
erex
posu
re a
nter
ior
topa
late
sur
face
(0)
, exp
osed
via
ext
ensi
onex
pose
d on
the
pala
te s
urfa
ce o
r co
vere
d by
the
post
erio
r in
terp
tery
goid
of p
oste
rior
inte
rpte
rygo
id v
acui
ties
(1)
pter
ygoi
ds. S
tate
one
ref
ers
to e
xpos
ure
of th
eva
cuiti
esex
pose
d w
ith la
tera
l pte
rygo
id s
utur
es (
2)pa
rasp
heno
id b
elow
the
pala
te s
urfa
ce d
ue to
enla
rgem
ent o
f the
pos
terio
r in
terp
tery
goid
vac
uitie
s;st
ate
two
refe
rs to
exp
osur
e on
the
pala
te s
urfa
ce,
acco
mpa
nied
by
late
ral s
utur
es w
ith th
e pt
eryg
oids
.F
igs.
6, 7
, 11,
and
13.

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 4573
Pal
ate
Pos
sess
ion
of c
rista
eP
rese
nt (
0), a
bsen
t (1)
Vau
ghn
1955
, Rei
szA
s de
scrib
ed a
bove
, the
ple
sios
aur
brai
ncas
e is
vent
rola
tera
les
1981
prim
itive
in th
e re
tent
ion
of c
rista
e ve
ntro
late
rale
s of
the
para
sphe
noid
, at l
east
in L
ower
Jur
assi
c ta
xa. T
heco
nfigu
ratio
n in
ear
ly p
lesi
osau
rs d
iffer
s fr
om e
arl ie
rdi
apsi
ds in
that
the
basi
sphe
noid
foss
a is
rep
lace
d by
a di
sc o
f end
ocho
ndra
l bon
e (b
asis
phen
oid)
con
fluen
tw
ith th
e m
edia
l mar
gins
of t
he c
rista
e ve
ntro
late
rale
s.F
ig. 1
.
74B
rain
case
Par
asph
enoi
d/A
bsen
t (0)
, pre
sent
(1)
Giv
en th
e co
nfigu
ratio
n de
scrib
ed in
cha
ract
er 7
3, th
eba
sioc
cipi
tal c
onta
ctpa
rasp
heno
id is
sep
arat
ed fr
om th
e ba
sioc
cipi
tal o
non
mid
l ine.
the
mid
l ine
by th
e ba
sisp
heno
id in
ear
ly p
lesi
osau
rs.
In s
ome
cryp
tocl
idid
s an
d al
l pol
ycot
ylid
s, th
epa
rasp
heno
id e
xten
ds b
ackw
ard
over
this
are
a an
dco
ntac
ts th
e ve
ntro
-ant
erio
r m
argi
n of
the
basi
occi
pita
lon
the
mid
l ine.
Fig
s. 1
6 an
d 17
.
75P
alat
eP
osse
ssio
n of
bas
alP
rese
nt (
0), a
bsen
t (1)
Illus
trat
ed fo
r T
hala
ssio
drac
on in
Sto
rrs
and
Tay
lor
artic
ulat
ion
(199
4) a
nd p
rese
nt in
Ple
sios
auru
s , th
e ba
sal
artic
ulat
ion
is p
rese
nt a
nd p
rom
inen
t in
all
ples
iosa
uroi
ds. T
he a
rtic
ulat
ion
appe
ars
abse
nt in
mos
t plio
saur
s; h
owev
er, o
ne s
kull
of P
elon
eust
es(B
MN
H R
. 380
3) p
rese
rves
this
feat
ure.
The
basi
sphe
noid
and
the
basa
l art
icul
atio
n ar
isin
g fr
om it
are
very
poo
rly o
ssifi
ed in
mos
t plio
saur
ids;
the
Pel
oneu
stes
sku
ll m
entio
ned
here
is u
nusu
al in
the
pres
erva
tion
of a
n ex
trem
ely
delic
ate
and
poor
lyos
sifie
d ba
sisp
heno
id in
whi
ch th
e ar
ticul
atio
n is
visi
ble.
The
are
a of
art
icul
atio
n w
ith th
e do
rsal
sur
face
of th
e pa
late
is u
sual
ly v
isib
le, h
owev
er, i
n pl
iosa
urm
ater
ial i
n w
hich
the
skul
l roo
f has
bee
n re
mov
ed.
The
fact
that
the
char
acte
r is
pre
sent
in a
ll ta
xa m
akes
this
cha
ract
er u
ninf
orm
ativ
e; it
is in
clud
ed d
ue to
prev
ious
con
fusi
on a
bout
the
pres
ence
of t
he b
asal
artic
ulat
ion.
Fig
s. 6
, 16,
and
17.
76P
alat
eB
asio
ccip
ital t
uber
sN
ot r
educ
ed (
0), r
educ
ed/ t
uber
face
tsA
ndre
ws
(191
0) il
lust
rate
s th
e ba
sioc
cipi
tal i
nre
duce
dco
nflue
nt w
ith b
asis
phen
oid
Tric
leid
us; i
n th
is ta
xon
and
in th
e po
lyco
tylid
s, th
ear
ticul
atio
n (1
)ba
sioc
cipi
tal t
uber
s ar
e re
duce
d, a
nd th
eir
pter
ygoi
dar
ticul
ar fa
cets
are
con
fluen
t with
the
med
ian
basi
sphe
noid
art
icul
atio
n. F
ig. 1
6.
Con
tinue
d
O’Keefe • ACTA ZOOL. FENNICA No. 21346A
pp
end
ix 1
. Con
tinue
d.—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—C
har.
type
Cha
ract
erS
tate
s/C
odin
gC
itatio
nR
emar
ks—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—77
Pal
ate
Juga
l has
sm
all
No
expo
sure
(0)
, exp
osur
e (1
)In
Pel
oneu
stes
and
som
e ot
her
plio
saur
s, th
e ju
gal
expo
sure
on
pala
teha
s a
smal
l exp
osur
e on
the
pala
te s
urfa
ce b
etw
een
surf
ace
the
max
i l la
and
squa
mos
al.
78P
alat
eLa
tera
l pal
atal
Abs
ent (
0), p
rese
nt (
1)S
ome
plio
saur
s ha
ve s
mal
l fen
estr
a in
the
pala
te,
fene
stra
tion
bord
ered
form
ed b
y th
e la
ck o
f a s
utur
e be
twee
n th
e pt
eryg
oid
by p
alat
ine
and
and
pala
tine
near
the
post
erio
r m
argi
n of
the
pala
tine.
pter
ygoi
dF
igs.
7 a
nd 8
.
79P
alat
eP
alat
ine/
inte
rnal
nar
isP
alat
ine
ente
rs in
tern
al n
aris
bor
der
(0),
The
pal
atin
e is
exc
lude
d fr
om th
e in
tern
al n
aris
by
aex
clud
ed b
y vo
mer
/max
i l la
cont
act (
1)su
ture
bet
wee
n th
e vo
mer
and
max
i l la
in L
iopl
euro
don
and
Plio
saur
us.
80P
alat
eP
alat
ines
app
roac
hD
o no
t mee
t (0)
, clo
se a
ppro
ach
or m
eet
Will
isto
n 19
03, 1
910
Fig
. 13.
clos
ely
or m
eet a
tat
mid
line
(1)
mid
line
81P
alat
eP
rem
axill
a/ a
nter
ior
Pre
max
illa
ente
rs a
nter
ior
bord
er (
0), i
sR
iepp
el 1
994a
, 199
7b,
bord
er o
f int
erna
l nar
isex
clud
ed b
y vo
mer
/ max
illa
cont
act (
1)ch
ars.
35,
45
82P
alat
eS
ub-o
rbita
l fen
estr
atio
nA
bsen
t (0)
, pre
sent
(1)
Sto
rrs
1991
, cha
r. 9
Sub
-orb
ital f
enes
tra
are
pres
ent i
n m
any
plio
saur
s.F
igs.
8 a
nd 1
3.
83P
alat
eV
omer
s ex
tend
far
Do
not (
0), e
xten
d po
ster
ior
and
mee
tF
igs.
9, 2
, 11,
etc
.po
ster
ior
to in
tern
alpt
eryg
oids
in w
ide
inte
rdig
itatin
g su
ture
(1)
nare
s on
mid
line
84P
alat
eP
rom
inen
t ‘pt
eryg
oid
Abs
ent (
0), p
rese
nt (
1)R
iepp
el 1
994a
, 199
7b,
As
disc
usse
d by
Sto
rrs
(199
7), t
he ‘p
tery
goid
flan
ge’
flang
e’/ e
ctop
tery
goid
char
s. 3
5, 4
4; S
torr
spr
esen
t in
som
e pl
iosa
urs
is n
ot a
true
pte
rygo
idbo
ss19
97fla
nge,
bec
ause
it is
com
pris
ed a
lmos
t ent
irely
by
the
ecto
pter
ygoi
d. S
torr
s th
eref
ore
sugg
ests
the
term
‘ect
opte
rygo
id b
oss’
for
this
feat
ure.
Fig
s. 1
1 an
d 13
.
85P
alat
eE
ctop
tery
goid
bos
sC
onta
ct a
bsen
t (0)
, con
tact
s ju
gal (
1)T
he e
ctop
tery
goid
bos
s ca
n ei
ther
be
free
of t
heha
s w
ide
cont
act w
ithm
edia
l wal
l of t
he s
kull
roof
, or
cont
act t
he ju
gal a
ndju
gal/
squa
mos
alor
squ
amos
al. F
ig. 1
6.
86M
andi
ble
Bow
edm
axill
aA
bsen
t(0)
pres
ent(
1)T
his
char
acte
rre
fers
tom
andi
bles
that
expa
nd
ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 47
86M
andi
ble
Bow
ed m
axill
aA
bsen
t (0)
, pre
sent
(1)
Thi
s ch
arac
ter
refe
rs to
man
dibl
es th
at e
xpan
dno
ticea
bly
ante
rior
of th
e ja
w a
rtic
ulat
ions
; suc
hm
andi
bles
hav
e a
hoop
-like
or
bow
ed a
ppea
ranc
ew
hen
view
ed o
n th
e ve
ntra
l sur
face
. Fig
. 7.
87M
andi
ble
Mec
kel ia
n ca
nal o
pen
Not
ope
n (0
) op
en (
1)T
he l i
ngua
l sur
face
of t
he lo
wer
jaw
is p
oorly
oss
ified
ante
riorly
in p
lesi
osau
rs; h
owev
er, i
n so
me
taxa
the
sple
nial
does
not
oss
ify a
t al l,
leav
ing
the
Mec
kel ia
n ca
nal
open
far
ante
rior
to th
e ja
w a
rtic
ulat
ion.
88M
andi
ble
Ven
tral
man
dibu
lar
No
ridge
(0)
, rid
ge (
1)T
he r
hom
aleo
saur
id ja
w s
ymph
ysis
has
a d
istin
ctrid
ge/ p
edes
tal-l
ike
ridge
or
pede
stal
-l ike
ela
bora
tion
alon
g its
ven
tral
sym
phys
ism
argi
n. T
his
ridge
ext
ends
pos
terio
rly fo
r a
shor
tdi
stan
ce p
ast t
he s
ymph
ysis
alo
ng e
ach
jaw
ram
us.
Fig
. 7.
89M
andi
ble
Man
dibu
lar
sym
phys
isS
hort
(0)
, som
ewha
t enf
orce
d (1
),M
odifi
ed fr
om R
iepp
elT
he le
ngth
of t
he m
andi
bula
r sy
mph
ysis
has
bee
nsc
oopl
ike
(2),
long
(3)
1997
b, c
har.
51;
Bro
wn
hist
oric
ally
impo
rtan
t in
clas
sify
ing
the
plio
saur
s (T
arlo
1981
, cha
r. 5
; Tar
lo19
60).
My
stat
e ‘s
coop
like’
app
lies
to b
oth
the
1960
rhom
aleo
saur
id a
nd L
iopl
euro
don
cond
ition
s; ‘l
ong’
appl
ies
to th
e co
nditi
on in
Pel
oneu
stes
. Fig
s. 7
and
13.
90M
andi
ble
Spl
enia
l par
ticip
ates
inD
oes
not p
artic
ipat
e (0
), d
oes
part
icip
ate
The
spl
enia
l ent
ers
the
man
dibu
lar
sym
phys
is in
all
sym
phys
is(1
), a
ngul
ars
exte
nd p
ast s
ymph
ysis
(2)
long
-sno
uted
taxa
. Sta
te tw
o de
scrib
es th
epo
lyco
tylid
s, in
whi
ch th
e sy
mph
ysis
is s
o lo
ng th
at it
exte
nds
post
erio
r to
the
ante
rior
mar
gin
of th
ean
gula
rs. T
his
char
acte
r is
ord
ered
. Fig
s. 9
and
12.
91M
andi
ble
Ling
ual m
andi
bula
rA
bsen
t (0)
, pre
sent
(1)
Tha
umat
osau
rus
poss
ess
a fe
nest
ra in
this
pos
ition
,fe
nest
rabo
unde
d by
the
sple
nial
, ang
ular
, and
pre
artic
ular
.F
ig. 7
.
92M
andi
ble
Mor
phol
ogy
of d
enta
ry/
Ang
ular
pro
ject
s fo
rwar
d of
sur
angu
lar
inIn
mos
t ple
sios
aurs
, the
sut
ure
of th
e de
ntar
y w
ith th
ean
gula
r- s
uran
gula
rla
tera
l vie
w (
0), s
uran
gula
r an
terio
ran
gula
r an
d su
rang
ular
on
the
late
ral s
urfa
ce o
f the
sutu
repr
oces
s (1
)m
andi
ble
is r
elat
ivel
y st
raig
ht (
Sto
rrs
& T
aylo
r 19
96);
how
ever
in p
liosa
urno
ids
the
sura
ngul
ar s
ends
a p
roce
ssan
terio
rly in
to a
n em
baym
ent i
n th
e de
ntar
y. F
ig. 1
2.
93M
andi
ble
Cor
onoi
dP
rese
nt (
0), a
bsen
t (1)
Cru
icks
hank
199
4b,
The
sta
tus
of th
e co
rono
id a
nd p
rear
ticul
ar is
a lo
ng-
char
. 9st
andi
ng p
robl
em in
ple
sios
aurs
. I b
elie
ve th
e co
rono
idto
be
pres
ent i
n al
l tax
a ex
cept
Kim
mer
osau
rus ,
cont
ra C
ruic
ksha
nk (
1994
b). F
igs.
12,
15,
and
19.
Con
tinue
d

O’Keefe • ACTA ZOOL. FENNICA No. 21348A
pp
end
ix 1
. Con
tinue
d.—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—C
har.
type
Cha
ract
erS
tate
s/C
odin
gC
itatio
nR
emar
ks—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—94
Low
er J
awLo
ng l i
ngua
l cor
onoi
dA
bsen
t (0)
, pre
sent
(1)
The
cor
onoi
d in
pl io
saur
s ru
ns a
nter
iorly
to th
epr
oces
sm
andi
bula
r sy
mph
ysis
. Fig
. 12.
95M
andi
ble
Cor
onoi
d ex
pose
d on
No
expo
sure
(0)
, exp
osur
e (1
)F
ig. 2
4. In
term
edia
te s
tate
in M
urae
nosa
urus
late
ral j
aw s
urfa
cei l l
ustr
ated
by
And
rew
s (1
910)
is c
orre
ct, a
lthou
gh h
eid
entifi
es th
is b
one
as th
e sp
leni
al r
athe
r th
an th
eco
rono
id F
ig 1
9.
96M
andi
ble
Pre
artic
ular
Pre
sent
(0)
, abs
ent (
1)C
ruic
ksha
nk 1
994b
,C
ruic
ksha
nk (
1994
b) b
elie
ved
the
prea
rtic
ular
to b
ech
ar. 9
pres
ent b
ut s
mal
l in
ples
iosa
urs;
I w
as a
ble
to c
onfir
mits
pre
senc
e in
al l
taxa
with
ade
quat
e pr
eser
vatio
n.F
igs.
12
and
19.
97M
andi
ble
Pre
artic
ular
she
lf/A
bsen
t (0)
, pre
sent
(1)
And
rew
s 19
10In
cry
ptoc
lidid
s an
d po
lyco
tylid
s, th
e pr
eart
icul
ar fo
rms
groo
vea
shel
f bel
ow th
e po
ster
ior
end
of th
e M
ecke
lian
cana
l,w
hich
is a
dis
tinct
gro
ove.
The
pre
artic
ular
fits
into
aso
cket
in th
e an
gula
r. N
oted
in M
urae
nosa
urus
by
And
rew
s (1
910)
. Fig
. 19.
98M
andi
ble
Jaw
art
icul
atio
n in
Abo
ve o
r at
col
linea
r w
ith to
oth
row
(0)
,F
igs.
12,
14,
and
15.
rela
tion
to to
oth
row
low
er th
an to
oth
row
(1)
99S
kull
shap
eD
iast
ema
at m
axill
a/A
bsen
t (0)
, pre
sent
(1)
Mod
ified
from
Sto
rrs
Fig
s. 2
and
11.
prem
axill
a su
ture
1991
, cha
r. 3
7
100
Den
titio
nF
irst t
ooth
afte
rLa
rge
(0),
red
uced
(1)
The
firs
t max
illar
y to
oth
is g
ener
ally
muc
h sm
alle
r th
andi
aste
ma
thos
e ca
udal
to it
in ta
xa p
osse
ssin
g a
dias
tem
a.F
igs.
2 a
nd 1
1.
101
Den
titio
nP
rem
axill
a an
d de
ntar
yA
bsen
t (0)
, pre
sent
(1)
Rie
ppel
199
4a, 1
997b
,F
ig. 7
.fa
ngs
char
s. 4
4, 5
5; S
torr
s 19
91,
char
. 39;
Bro
wn
1981
,ch
ar. 8
102
Den
titio
nO
ne o
r tw
o ca
nini
form
Pre
sent
(1)
, abs
ent (
0)R
iepp
el 1
994a
, 199
7b,
teet
h on
max
illa
char
s. 4
5, 5
6; S
torr
s 19
91,
char
. 41
103
Den
titio
nT
ooth
form
Gra
cile
smal
lroo
tna
rrow
now
ear
(0)
Bro
wn
1981
char
3F
igs
45
11an
d14
Too
thty
pes
defin
edby
Mas
sare

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 49
103
Den
titio
nT
ooth
form
Gra
cile
, sm
all r
oot,
narr
ow, n
o w
ear
(0)
Bro
wn
1981
, cha
r. 3
,F
igs.
4, 5
, 11,
and
14.
Too
th ty
pes
defin
ed b
y M
assa
rero
bust
, lar
ge r
oot,
wea
r (1
), v
ery
smal
l/T
arlo
196
0(1
987)
wer
e no
t use
d be
caus
e m
ost p
lesi
osau
rs h
ave
need
le-li
ke (
2)th
e sa
me
type
of t
eeth
in th
is m
ore
gene
ral c
odin
g.
104
Too
th s
hape
Tee
th r
ound
or
with
Rou
nd (
0), p
lana
r fa
ce (
1)T
arlo
196
0re
info
rced
pla
nar
face
105
Too
th o
rnam
ent
Long
itudi
nal s
tria
tions
Str
iatio
ns a
l l ar
ound
(0)
, lin
gual
onl
y (1
),T
arlo
196
0on
teet
hno
ne (
2)
106
Den
titio
nN
umbe
r of
pre
max
i l lar
y5(
0), 6
(1),
7(2
), g
reat
er th
an 7
(3)
Bro
wn
1981
, cha
r. 7
;T
his
char
acte
r is
ord
ered
. Fig
s. 2
and
11.
teet
hC
ruic
ksha
nk 1
994b
,ch
ar. 1
107
Den
titio
nM
axi l l
ary
teet
hLe
ss th
an tw
enty
(0)
, mor
e tw
enty
toC
ruic
ksha
nk 1
994b
,T
his
char
acte
r is
ord
ered
.th
irty
(1),
man
y m
ore
than
thirt
y (2
)ch
ar. 1
108
Atla
s/ A
xis
Num
ber
of a
xis
rib2(
0), 1
(1)
Thi
s ch
arac
ter
is d
ifficu
lt to
sco
re in
man
y ta
xa d
ue to
head
spo
or p
rese
rvat
ion
and/
or in
com
plet
e os
sific
atio
n.
109
Atla
s/ A
xis
Art
icul
atio
n of
axi
s rib
Bro
ad a
rtic
ulat
ion
with
atla
s ce
ntru
mA
ndre
ws
1910
, 191
3T
his
char
acte
r is
var
iabl
e w
ithin
Pel
oneu
stes
, and
may
and/
or o
ther
ele
men
ts (
0), h
ead
confi
ned
be o
ntog
enet
ical
ly v
aria
ble.
Diffi
cult
to s
core
in a
dult
to a
xis
cent
rum
(1)
indi
vidu
als
in w
hich
the
atla
s/ax
is c
ompl
ex is
gen
eral
ly fu
sed.
110
Atla
s/ A
xis
Atla
s/ax
is m
orph
olog
yN
o la
tera
l exp
osur
e of
atla
s ce
ntru
m o
n cu
pA
ndre
ws
1909
, Will
isto
nfa
ce (
0), l
ater
al e
xpos
ure
(1),
no
late
ral
1910
, Bro
wn
1981
, cha
r.ex
posu
re, b
ut a
tlas
and
axis
inte
rcen
tra
13. A
lso
Rom
er, 1
956
excl
ude
atla
s ce
ntru
m v
entr
ally
(2)
111
Ver
tebr
aeN
umbe
r of
cer
vica
lP
rimiti
ve (
0), i
ncre
ased
(1)
, red
uced
(2)
,B
row
n 19
81, c
har.
14
The
num
ber
of c
ervi
cal v
erte
brae
is u
nfor
tuna
tely
vert
ebra
egr
eate
r th
an 5
0un
know
n in
Pis
tosa
urus
(S
ues
1987
); h
owev
er th
is ta
xon
is s
imila
r to
ples
iosa
urs
in p
osse
ssin
g ‘p
ecto
ral’
verte
brae
,de
fined
as
vert
ebra
e in
whi
ch th
e rib
art
icul
atio
n ar
ises
from
a s
hort
tran
sver
se p
roce
ss a
risin
g fr
om b
oth
the
cent
rum
and
neu
ral a
rch
(illu
stra
ted
in B
row
n, 1
981)
.H
ere
I defi
ne c
ervi
cal v
erte
brae
as
vert
ebra
e in
whi
chth
e rib
art
icul
atio
n ar
ises
from
the
cent
rum
onl
y. B
row
n(1
981)
als
o be
lieve
d 28
–32
cerv
ical
s to
be
prim
itive
inpl
esio
saur
s; I
have
ret
aine
d th
is c
riter
ion.
Sta
te o
ne is
33 c
ervi
cals
or
larg
er, w
hile
sta
te tw
o is
few
er th
an 2
8.S
tate
thre
e co
ncer
ns n
ecks
of 5
0 ce
rvic
als
or lo
nger
.T
his
char
acte
r is
ord
ered
usi
ng th
e ch
arac
ter
stat
e tr
ee((
2)1,
3)0.
Con
tinue
d

O’Keefe • ACTA ZOOL. FENNICA No. 21350A
pp
end
ix 1
. Con
tinue
d.—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—C
har.
type
Cha
ract
erS
tate
s/C
odin
gC
itatio
nR
emar
ks—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—11
2M
orph
o-m
etric
Pro
port
ions
of c
ervi
cal
Leng
th e
qual
to h
eigh
t (0)
, len
gth
grea
ter
Bro
wn
1981
, cha
r. 1
5;W
elle
s (1
952)
iden
tified
ont
ogen
etic
var
iatio
n in
this
cent
rath
an h
eigh
t (1)
, len
gth
less
than
Wel
les
1952
char
acte
r, a
s w
ell a
s va
riatio
n al
ong
the
vert
ebra
lhe
ight
(2)
colu
mn,
but
als
o fo
und
that
taxo
nom
ical
ly r
elev
ant
inte
rspe
cific
diff
eren
ces
exis
ted.
I sc
ored
this
char
acte
r lo
osel
y; v
erte
brae
had
to b
e m
arke
dly
com
pres
sed
or e
long
ated
ove
r m
ost o
f the
col
umn
tobe
sco
red
as d
eriv
ed.
113
Ver
tebr
aeD
istin
ct c
hang
e in
No
chan
ge in
ang
le (
0), c
hang
e (1
)N
oted
by
Wel
les
1962
In e
lasm
osau
rs a
nd s
ome
othe
r ta
xa, t
he c
ervi
cal
zygo
poph
ysea
l ang
lezy
gopo
phys
es a
re o
rient
ed d
orso
-ven
tral
ly in
the
alon
g ce
rvic
al c
olum
nan
terio
r pa
rt o
f the
col
umn.
Thi
s or
ient
atio
n sh
ifts
grad
ually
to m
edio
-late
ral i
n th
e po
ster
ior
part
of t
hene
ck.
114
Ver
tebr
aeV
entr
al k
eel o
n ce
rvic
alA
bsen
t (0)
, pre
sent
(1)
Tar
lo 1
960
vert
ebra
e
115
Ver
tebr
aeLa
tera
l rid
ge o
nA
bsen
t (0)
, pre
sent
(1)
Bro
wn
1981
, cha
r. 1
6;A
low
, lat
eral
rid
ge is
pre
sent
on
the
cent
rum
bod
yan
terio
r ce
rvic
alill
ustr
ated
in W
elle
s,ab
ove
the
rib a
rtic
ulat
ion
in e
lasm
osau
rs a
nd s
ome
vert
ebra
e in
adu
lts19
43ot
her
ples
iosa
uroi
ds.
116
Ver
tebr
aeB
inoc
ular
sha
ped
Abs
ent (
0), p
rese
nt (
1)W
elle
s 19
43T
he a
nter
ior
cerv
ical
cen
tra
in s
ome
elas
mos
aurs
ante
rior
cerv
ical
cen
tra
have
a d
istin
ct d
orso
-ven
tral
con
stric
tion
med
ially
,yi
eldi
ng a
cen
trum
that
has
a ‘b
inoc
ular
’ out
line
inan
terio
r or
pos
terio
r vi
ew.
117
Ver
tebr
aeN
umbe
r of
cer
vica
l rib
Tw
o (0
), o
ne (
1)S
ande
r et
al .
1997
;he
ads
Bro
wn
1981
, cha
r. 2
1
118
Ver
tebr
aeV
entr
al fo
ram
ina
inA
bsen
t (0)
, pre
sent
(1)
San
der
et a
l. 19
97;
Ter
med
“fo
ram
ina
subc
entr
alia
” by
Sto
rrs
(199
1), t
hece
rvic
al v
erte
brae
Sto
rrs
1991
, cha
r. 4
7po
sses
sion
of p
aire
d fo
ram
ina
in th
e ve
ntra
l sur
face
of
the
cerv
ical
cen
tra
is a
trai
t sha
red
by p
isto
saur
ids
and
all p
lesi
osau
rs.
119
Ver
tebr
aeF
oram
ina
subc
entr
alia
Med
ial a
nd la
rge
(0),
late
ral a
ndT
arlo
196
0; s
ee a
lso
redu
ced
and
late
ral
redu
ced
(1)
Will
isto
n 19
07
120
Ver
tebr
aeW
idth
of c
ervi
cal
Wid
er th
an c
entr
um (
0), s
ubeq
ual w
ithS
ande
r et
al.
1997
;T
his
char
acte
r is
ord
ered
.zy
gopo
phys
esce
ntru
m (
1), m
ore
narr
ow th
anS
torr
s 19
91 c
har.
48
cent
rum
(2)
ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 51
cent
rum
(2)
121
Ver
tebr
aeP
oste
rior
artic
ulat
ion
Abs
ent (
0), p
rese
nt (
1)N
oted
in P
isto
saur
us b
y (S
ues
1987
) an
d in
for
succ
eedi
ng n
eura
lC
alla
way
asau
rus
(Wel
les
1962
), th
is fe
atur
e is
a s
mal
lsp
ine,
cer
vica
lno
tch
in th
e po
ster
ior
base
of t
he c
ervi
cal n
eura
l spi
neve
rteb
rae
whi
ch a
ccep
ts th
e an
terio
r ed
ge o
f the
suc
ceed
ing
neur
al s
pine
. Thi
s no
tch
may
rep
rese
nt th
e zy
gant
rum
pres
ent i
n m
ore
basa
l sau
ropt
eryg
ians
; how
ever
, no
corr
espo
ndin
g zy
gosp
hene
is e
ver
pres
ent.
Lost
inm
any
plio
saur
s.
122
Ver
tebr
aeC
ervi
cal r
ib a
rtic
ulat
ion
Circ
ular
or
subc
ircul
ar (
0), e
long
ate
(1)
Il lus
trat
ed in
Wel
les
Ant
ero-
post
erio
r el
onga
tion
of th
e ce
rvic
al r
ibs,
and
grea
tly e
long
ate/
1943
elon
gatio
n of
thei
r ar
ticul
atio
n w
ith th
e ce
ntru
m, i
sce
rvic
al r
ibs
expa
nded
com
mon
in lo
ng-n
ecke
d ta
xa in
whi
ch th
e ce
ntru
m is
and
blad
e-l ik
elo
nger
than
wid
e.
123
Ver
tebr
aeA
nter
ior
proc
ess
ofP
rese
nt (
0), a
bsen
t (1)
Il lus
trat
ed in
San
der
A d
istin
ct a
nter
ior
proc
ess
is p
rese
nt o
n th
e ce
rvic
alce
rvic
al r
ibs
et a
l. 19
97, S
torr
s 19
97rib
s of
Aug
usta
saur
us, a
s w
ell a
s T
hala
ssio
drac
on,
Ple
sios
auru
s , a
nd s
ome
othe
r Ju
rass
ic p
lesi
osau
rs.
Thi
s pr
oces
s is
lost
in la
ter
ples
iosa
urs.
124
Ver
tebr
aeA
nter
ior
neur
al fl
ange
Abs
ent (
0), p
rese
nt (
1)M
urae
nosa
urus
and
som
e ot
her
ples
iosa
uroi
ds h
ave
aon
cer
vica
l neu
ral
dist
inct
flan
ge o
n th
e an
terio
r m
argi
n of
the
cerv
ical
spin
esne
ural
spi
nes.
Thi
s ch
arac
ter
is n
ot n
eces
sary
to fo
rmth
e ar
ticul
atio
n de
scrib
ed in
cha
ract
er 1
21.
125
Ver
tebr
aeN
eura
l spi
nes,
Ang
led
back
war
d (0
), n
ot a
ngle
d (1
)B
row
n 19
81, c
har.
20
The
cer
vica
l neu
ral s
pine
s in
ela
smos
aurs
and
som
ece
rvic
al v
erte
brae
othe
r ta
xa a
re n
ot a
ngle
d, w
hile
they
are
ang
led
back
war
d in
Tha
lass
iodr
acon
and
the
plio
saur
oids
.
126
Ver
tebr
aeD
ista
l end
of
No
diam
eter
incr
ease
(0)
, thi
cken
ed (
1)R
iepp
el 1
997b
, cha
r. 6
8T
hick
ened
tran
sver
se p
roce
sses
are
pre
sent
in a
lltr
ansv
erse
pro
cess
es,
ples
iosa
urs,
Pis
tosa
urus
, and
Aug
usta
saur
us.
dors
al v
erte
brae
127
Ver
tebr
aeD
orsa
l neu
ral a
rch
Sub
equa
l to
cent
rum
hei
ght (
0), s
hort
erS
ande
r et
al .
1997
The
neu
ral a
rch
tend
s to
be
smal
l and
sho
rt c
ompa
red
heig
htth
an c
entr
um h
eigh
t (1)
to th
e ce
ntru
m in
ple
sios
aurs
, whi
le it
is la
rger
in m
ore
basa
l sau
ropt
eryg
ians
.
128
Ver
tebr
aeZ
ygos
phen
e/P
rese
nt (
0), a
bsen
t (1)
San
der
et a
l . 19
97;
The
pis
tosa
urid
con
ditio
n is
als
o di
scus
sed
in S
ues
zyga
ntru
m a
rtic
ulat
ion
Rie
ppel
199
4a, 1
997b
,(1
987)
.ch
ars.
53,
67;
Sto
rrs
1991
, cha
r. 4
9
Con
tinue
d
O’Keefe • ACTA ZOOL. FENNICA No. 21352A
pp
end
ix 1
. Con
tinue
d.—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—C
har.
type
Cha
ract
erS
tate
s/C
odin
gC
itatio
nR
emar
ks—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—12
9V
erte
brae
Hei
ght o
f neu
ral
Low
(0)
, low
and
rug
ose
(1),
hig
h (2
)S
ande
r et
al.
1997
The
dor
sal n
eura
l spi
nes
of A
ugus
tasa
urus
are
low
spin
es, d
orsa
lw
ith r
ugos
e to
ps; t
he s
pine
s in
Tha
lass
iodr
acon
are
vert
ebra
elo
w b
ut n
ot r
ugos
e; th
ose
in P
isto
saur
us a
re h
igh.
The
dors
al n
eura
l spi
nes
are
high
in a
l l pl
esio
saur
s.
130
Ver
tebr
aeLa
tera
l com
pres
sion
Not
com
pres
sed
(0),
com
pres
sed
and
San
der
et a
l. 19
97T
he n
eura
l spi
nes
in m
any
ples
iosa
urs
are
late
ral ly
of n
eura
l spi
nes,
blad
e-l ik
e (1
)co
mpr
esse
d an
d bl
ade-
l ike;
the
neur
al s
pine
s of
man
ydo
rsal
and
cer
vica
lpl
iosa
urs
lack
this
feat
ure.
vert
ebra
e
131
Pec
tora
l Gird
leIn
terc
lavi
cle
post
erio
rP
rese
nt (
0), a
bsen
t (1)
Sto
rrs
1991
, cha
r. 5
8T
he in
terc
lavi
cle
was
pro
babl
y pr
esen
t but
is n
otpr
oces
skn
own
in m
any
ples
iosa
urs.
132
Pec
tora
l Gird
leD
orsa
l pro
cess
of
Tap
ers
to b
lunt
tip
(0),
ven
tral
lyR
iepp
el 1
997b
, cha
r. 8
5sc
apul
aex
pand
ed p
oste
riorly
(1)
133
Pec
tora
l Gird
leP
rese
nce
of c
lavi
cles
Pre
sent
(0)
, int
ercl
avic
le a
bsen
t (1)
,M
odifi
ed fr
om B
row
nT
he in
terc
lavi
cle
and
clav
icle
s ar
e sl
ow to
oss
ify a
ndan
d in
terc
lavi
cle
both
abs
ent (
2)19
81, c
hars
. 22
and
23of
ten
do n
ot p
rese
rve
wel
l; P
lesi
osau
rus
is th
e on
lyta
xon
in w
hich
I co
uld
be s
ure
of th
e ab
senc
e of
the
inte
rcla
vicl
e, b
ased
on
Sto
rrs
1997
and
my
own
obse
rvat
ions
.
134
Pec
tora
l Gird
leC
lavi
cle
med
ian
Sym
phys
is (
0), s
epar
ated
by
inte
rcla
vicl
esy
mph
ysis
(1),
mee
t onl
y be
hind
not
ch (
2)
135
Pec
tora
l Gird
leS
capu
lae
mee
t in
Sep
arat
ed b
y cl
avic
les/
inte
rcla
vicl
e (0
),M
odifi
ed fr
om B
row
nT
he s
capu
lae
mee
t in
an a
nter
ior
med
ian
sym
phys
isan
terio
r m
edia
nm
eet m
edia
lly b
ut le
ave
notc
h fo
r de
rmal
1981
, cha
r. 2
4in
man
y la
ter
ples
iosa
urs;
how
ever
, the
ant
ero-
med
ial
sym
phys
isel
emen
ts (
1), m
eet i
n lo
ng s
ymph
ysis
proc
esse
s of
the
scap
ulae
are
usu
ally
sep
arat
ed b
yw
ith n
o no
tch
(2)
the
derm
al g
irdle
ele
men
ts in
plio
saur
s (c
ontr
a B
row
n19
81).
136
Pec
tora
l Gird
leA
nter
ior
intr
asca
pula
rA
bsen
t (0)
, pre
sent
(1)
Som
e cr
ypto
clid
ids
and
som
e po
lyco
tylid
s ha
ve a
fene
stra
smal
l fen
estr
a pe
rfor
atin
g th
e an
terio
r gi
rdle
ele
men
tson
the
mid
line.
137
Pec
tora
l Gird
leLo
ngitu
dina
l pec
tora
lA
bsen
t (0)
, for
med
by
clav
icle
and
Mod
ified
from
Sto
rrs
Thi
s ch
arac
ter
is c
erta
inly
var
iabl
e on
toge
netic
ally
, as
bar
cora
coid
(1)
, for
med
by
scap
ula
and
1991
, cha
r. 6
6; B
row
ndi
scus
sed
by B
row
n (1
981)
. I in
clud
e it
here
bec
ause
cora
coid
(2)
1981
, cha
r. 2
5m
ost o
f the
taxa
in th
is a
naly
sis
are
adul
ts, a
nd fo
r th
esa
ke o
f com
plet
enes
s.
138
Pec
tora
lGird
leS
prac
orac
oid
Pre
sent
(0)
abse
nt(1
)R
iepp
el19
97b
char
64T
hes
prac
orac
oid
fora
men
isab
sent
inpi
stos
arid
s

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 5313
8P
ecto
ral G
irdle
Sup
raco
raco
idP
rese
nt (
0), a
bsen
t (1)
Rie
ppel
199
7b, c
har.
64;
The
sup
raco
raco
id fo
ram
en is
abs
ent i
n pi
stos
aurid
sfo
ram
en/ n
otch
Sto
rrs
1991
, cha
r. 6
4an
d al
l ple
sios
aurs
.
139
Pec
tora
l Gird
leC
orac
oid
shap
eR
ound
ed c
onto
urs/
not
pla
te-li
ke (
0),
Rie
ppel
199
7b, c
har.
88;
expa
nded
med
ian
sym
phys
is (
1)S
torr
s 19
91, c
har.
63
140
Pec
tora
l Gird
leM
edia
n co
raco
idA
bsen
t (0)
, pre
sent
(1)
Il lus
trat
ed in
Wil l
isto
nM
ost p
olyc
otyl
ids
have
one
or
two
perf
orat
ions
on
perf
orat
ions
1906
, 190
6ei
ther
sid
e of
the
mid
l ine
in th
e po
ster
ior
exte
nsio
ns o
fth
e co
raco
ids.
141
Pec
tora
l Gird
leP
oste
rior
cora
coid
Abs
ent (
0), p
rese
nt (
1)B
row
n 19
81, c
har.
27
Wel
les
(194
3, 1
962)
and
man
y ot
hers
hav
eex
tens
ion
with
dee
pdo
cum
ente
d th
at th
e po
ster
ior
exte
nsio
ns o
f the
med
ian
emba
ymen
tco
raco
ids
are
long
and
sur
roun
d a
deep
med
ian
emba
ymen
t in
man
y el
asm
osau
rs.
142
Pec
tora
l Gird
leP
oste
ro-la
tera
lA
bsen
t (0)
, pre
sent
(1)
Bro
wn
1981
, cha
r. 2
6S
torr
s (1
997)
and
Bro
wn
(198
1) d
iscu
ss th
e cl
ear
cora
coid
win
gson
toge
netic
var
iatio
n in
this
cha
ract
er. I
t is
incl
uded
here
bec
ause
mos
t gen
era
in th
is a
naly
sis
are
adul
ts,
and
for
the
sake
of c
ompl
eten
ess.
143
Pel
vic
Gird
leC
onta
ct b
etw
een
Pre
sent
(0)
, abs
ent (
1)S
torr
s 19
91, c
har.
68;
The
loss
of t
he p
rimiti
ve c
onta
ct b
etw
een
the
ilium
and
ilium
and
pub
isS
ues
1987
pubi
s ha
s lo
ng b
een
used
as
a di
agno
stic
cha
ract
er o
fth
e P
lesi
osau
ria (
i.e. R
omer
, 195
6, p
. 329
).
144
Pel
vic
Gird
leP
ubis
ven
tral
Con
vex
(1),
con
cave
(0)
Rie
ppel
199
7b, c
har.
100
;(m
edia
l) m
argi
nS
torr
s 19
91, c
har.
69;
Bro
wn
1981
, cha
r. 2
8
145
Pel
vic
Gird
leLa
rge,
ven
tral
Abs
ent (
0), p
rese
nt (
1)S
ues
1987
A la
rge
vent
ral p
late
form
ed b
y th
e is
chiu
m a
nd p
ubis
pubo
-isch
iatic
pla
teis
cha
ract
eris
tic o
f ple
sios
aurs
.
146
Pel
vic
Gird
leM
edia
n pe
lvic
bar
Abs
ent (
0), p
rese
nt (
1)B
row
n 19
81, c
har.
30
Ana
logo
us to
the
med
ian
pect
oral
bar
, the
med
ian
pelv
ic b
ar is
slo
w to
to o
ssify
, as
disc
usse
d by
Bro
wn
(198
1). I
incl
ude
it he
re b
ecau
se m
ost o
f the
taxa
inth
is a
naly
sis
are
adul
ts, a
nd fo
r th
e sa
ke o
fco
mpl
eten
ess.
The
cha
ract
er is
als
o di
fficu
lt to
sco
re;
the
pres
ence
of a
sut
ure
betw
een
the
med
ian
proc
esse
s of
the
pubi
s an
d is
chiu
m c
an o
nly
bede
term
ined
by
artic
ulat
ing
the
two
elem
ents
.
147
Pel
vic
Gird
leT
hyro
id fe
nest
raC
lose
d (0
), o
pen
(1)
Rie
ppel
199
7b, c
har.
102
;cl
osed
or
open
in a
dult
Sto
rrs
1991
cha
r. 7
1
Con
tinue
d

O’Keefe • ACTA ZOOL. FENNICA No. 21354
Ap
pen
dix
1. C
ontin
ued.
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
Cha
r. ty
peC
hara
cter
Sta
tes/
Cod
ing
Cita
tion
Rem
arks
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
148
Pel
vic
Gird
leO
btur
ator
fora
men
Pre
sent
(0)
, abs
ent (
1)R
iepp
el 1
997b
, cha
r. 1
01;
The
obt
urat
or fo
ram
en is
lost
in s
ome
pist
osau
roid
sS
torr
s 19
91, c
har.
70
and
all p
lesi
osau
rs; s
ee d
iscu
ssio
n ab
ove.
149
Pel
vic
Gird
leIl i
ac b
lade
Wel
l-dev
elop
ed (
0), r
educ
ed (
1)R
iepp
el 1
994a
, 199
7b,
char
s. 7
9, 9
9; S
torr
s 19
91,
char
. 67;
Bro
wn
1981
, cha
r.31
; Sue
s 19
87
150
Hum
erus
Ang
led
hum
erus
Abs
ent (
1), p
rese
nt (
0)R
iepp
el 1
994a
, 199
7b,
The
ang
led
hum
erus
is c
hara
cter
istic
of s
tem
-gro
upch
ars.
74,
92;
Sto
rrs
saur
opte
rygi
ans,
and
of e
arly
ple
sios
aurs
. See
Sto
rrs
1991
cha
r. 7
3(1
997)
for
the
cond
ition
in P
lesi
osau
rus .
151
Pro
-pod
ials
Exp
ande
d di
stal
Abs
ent (
0), p
rese
nt (
1)S
hort
pro
podi
als
who
se d
ista
l end
s ar
e ex
pand
edpr
opod
ials
with
dor
sal
ante
ro-p
oste
riorly
and
flat
tene
d do
rso-
vent
rally
are
troc
hant
er/ t
uber
osity
char
acte
ristic
of a
ll pl
esio
saur
s. T
he p
osse
ssio
n of
ado
rsal
troc
hant
er o
r tu
bero
sity
is a
lso
char
acte
ristic
of
all p
lesi
osau
r pr
opod
ials
.
152
Hum
erus
Dis
tal e
nd o
f hum
erus
Abs
ent (
0), p
rese
nt (
1)Ill
ustr
ated
by
Will
isto
nT
he h
umer
us in
man
y pl
esio
saur
oids
has
two
dist
inct
has
two
dist
inct
1906
plan
es o
r fa
cets
on
its d
ista
l end
, with
whi
ch th
epl
anes
in a
dult
epip
odia
ls a
rtic
ulat
e.
153
Pro
podi
als
Dis
tinct
face
t on
dist
alA
bsen
t (0)
, pre
sent
(1)
Illus
trat
ed b
y W
illis
ton
Tric
leid
us a
nd P
olyc
otyl
us h
ave
a th
ird d
istin
ct fa
cet
hum
erus
for
1906
on th
e po
ster
o-di
stal
end
of t
he h
umer
us w
hich
supe
rnum
ery
artic
ulat
es w
ith a
sup
ernu
mer
y os
sific
atio
n of
the
ossi
ficat
ion
epip
odia
l row
.
154
Pro
podi
als
Gra
cile
or
mas
sive
Gra
cile
/con
stric
ted
(0),
mas
sive
(1)
Bro
wn
1981
, cha
r. 3
2A
pplic
able
onl
y to
ple
sios
aurs
.
155
Pro
podi
als
Pro
podi
als
rela
tivel
yN
ot e
long
ate
(0),
elo
ngat
e w
ith n
arro
wLa
te p
liosa
urs
such
as
Bra
chau
chen
ius
have
long
,el
onga
tedi
stal
hea
d (1
)na
rrow
pro
podi
als
com
pare
d to
ear
lier
plio
saur
s.
156
Hum
erus
Del
tope
ctor
al c
rest
Pre
sent
(0)
, abs
ent (
1)R
iepp
el 1
997b
, cha
r. 9
3

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 5515
7U
lna
Uln
ar s
hape
Nar
row
(0)
or
broa
d (1
) di
stal
lyS
ande
r et
al 1
997;
A b
road
dis
tal u
lna
is th
ough
t to
be a
syn
apom
orph
yS
torr
s 19
91, c
har.
83
of A
gust
asau
rus
and
Pis
tosa
urus
(S
ande
r et
al.
1997
);ho
wev
er I
bel ie
ve it
form
s pa
rt o
f a tr
ansf
orm
atio
nse
ries
of d
ecre
asin
g pe
richo
ndra
l oss
ifica
tion
inpi
stos
auro
ids
and
ples
iosa
urs.
I ha
ve th
eref
ore
code
dit
as p
rese
nt in
ple
sios
aurs
rat
her
than
inap
plic
able
.
158
Uln
aD
istin
ctly
luna
te u
lna
Abs
ent (
0), p
rese
nt (
1)M
any
early
ple
sios
aurs
hav
e an
uln
a w
hich
isdi
stin
ctly
luna
te in
sha
pe.
159
Fem
urIn
tern
al tr
ocha
nter
Wel
l-dev
elop
ed (
0), r
educ
ed (
1)R
iepp
el 1
997b
, cha
r. 1
05B
oth
this
cha
ract
er a
nd c
hara
cter
160
are
rel
ated
toth
e pr
ogre
ssiv
e lo
ss o
f fem
oral
feat
ures
as
saur
opte
rygi
ans
beca
me
mor
e aq
uatic
.
160
Fem
urIn
ter-
troc
hant
eric
foss
aD
eep
(0),
dis
tinct
but
red
uced
(1)
,R
iepp
el 1
997b
, cha
r. 1
06ru
dim
enta
ry o
r ab
sent
(2)
161
Epi
-pod
ials
Epi
podi
al m
orph
olog
yLo
nger
than
bro
ad (
0), e
qual
or
broa
der
Bro
wn
1981
, cha
r. 3
5.;
than
long
(1)
Sue
s 19
87
162
Epi
-pod
ials
Sup
ernu
mer
yN
one
(0),
epi
podi
al r
ow/p
isifo
rm (
1),
Mod
ified
from
Sto
rrs
Ple
sios
aurs
ofte
n ha
ve a
sup
ernu
mer
y os
sific
atio
n in
ossi
ficat
ions
, for
elim
bpr
opod
ial (
2), b
oth
(3)
1991
,cha
r. 8
4; B
row
nth
e ep
ipod
ial r
ow a
nd/o
r on
a le
vel w
ith th
e di
stal
1981
, cha
r. 3
4pr
opod
ial.
The
se b
ones
are
slo
w to
oss
ify a
nd a
relia
ble
to lo
ss d
urin
g co
llect
ion.
163
Met
a-po
dial
sF
ifth
met
apod
ial
In li
ne w
ith r
est o
f met
apod
ial r
ow (
0),
Ow
en 1
865,
Bro
wn
Ow
en (
1865
) w
as th
e fir
st to
not
e th
at th
e fif
thsh
ifted
into
dis
tal m
esop
odia
l row
1981
, cha
r. 3
7m
etap
odia
l is
shift
ed in
to th
e di
stal
mes
opod
ial r
ow in
all p
lesi
osau
rs. T
his
shift
res
ults
in a
mis
alig
nmen
t of
the
phal
ange
al a
rtic
ulat
ions
of d
igit
five
rela
tive
to th
eot
her
digi
ts.
164
Pha
lang
esH
yper
phal
angy
No
incr
ease
(2–
3–4–
5–3)
(0)
,M
odifi
ed fr
om S
torr
shy
perp
hala
ngy
pres
ent (
1)19
91, c
har.
80
165
Pha
lang
esIn
terlo
ckin
g di
stal
Abs
ent (
0), p
rese
nt (
1)T
he p
hala
nges
of T
rinac
rom
erum
and
rel
ated
taxa
phal
ange
s an
terio
r to
have
join
ts w
hich
are
mis
alig
ned
betw
een
digi
ts in
fifth
pha
lang
eal r
owdi
gits
one
thro
ugh
four
. Thi
s ch
arac
ter
is d
ifficu
lt to
scor
e du
e to
con
cern
s ov
er p
rese
rvat
ion.
166
Rib
sM
edia
n ga
stra
l rib
Alw
ays
one
late
ral p
roce
ss (
0), m
ayR
iepp
el 1
997b
, cha
r. 1
19el
emen
tha
ve tw
o la
tera
l pro
cess
es (
1)—
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
——
—

O’Keefe • ACTA ZOOL. FENNICA No. 21356
Appendix 2. Data matrix for the cladistic analysis in this paper. The first three taxa comprise the outgroup.Inapplicable characters are coded as ‘x’; unknown characters are coded as ‘?’. The ancestral condition isgenerally coded as ‘0’.————————————————————————————————————————————————Taxon 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19————————————————————————————————————————————————Simosaurus 0 0 0 0 2 2 x 0 0 0 0 ? 0 1 2 0 0 0 ?Cymatosaurus 0 ? ? ? ? 2 x 0 1 0 0 ? 0 0 1,2 1 0 1 ?Pistosauridae 0 0 ? 1 ? 0 x 1 0 0 0 ? 0 0 1 1 0 1 ?Calawayasaurus 2 1 2 0 2 0 0 0 0 0 0 0 0 0 x 0 1 1 ?Attenborosaurus 1 0 1 1 0 0 0 1 1 0 0 ? ? 0 0 ? 0 ? ?BMNH R.5488 1 0 1 1 1 0 1 1 1 0 ? 0 1 0 ? ? 0 0 1Brachauchenius 1 2 ? ? ? 1 ? 2 1 0 1 1 0 x x 0 0 0 0Brancasaurus 2 1 2 1 0 0 0 0 0 0 0 0 0 1 x 0 0 1 ?Cryptoclidus 2 0 0 1 2 0 1 0 0 0 0 0 0 0 x 0 0 0 1Dolichorhynchops 1 2 1 1 0 1 1 2 0 0 1 0 0 x x 0 0 0 ?Eurycleidus 0 ? 0 ? ? 0 0 1 1 0 0 ? 0 0 0 1 0 0 ?Hauffiosaurus 1 0 2 1 0 1 0 2 1 0 ? ? ? ? ? ? ? ? ?Kimmerosaurus ? ? ? ? ? 2 ? 3 0 0 0 ? ? 0 x 0 ? 0 ?Kronosaurus 1 2 2 1 ? 1 ? 2 ? ? ? ? ? ? ? ? ? ? ?Leptocleidus ? 2 ? 1 ? 0 1 1 1 0 1 0 1 x x 0 0 0 ?Libonectes 2 1 ? 0 ? 0 0 0 0 0 2 0 0 x x ? 1 ? ?Liopleurodon 1 2 1 1 1 1 1 2 2 0 1 1 0 x x 0 0 0 1Macroplata 1 0 2 1 0 1 0 2 0 0 ? 1 0 x 0 0 ? ? 0Microcleidus 2 1 0 1 0 0 0 0 0 0 0 0 0 ? ? 0 ? 1 ?MOR 751 0 2 ? ? ? 0 1 1 0 0 0 1 0 0 x 0 0 0 1Morturneria ? ? ? ? ? 2 1 3 0 0 ? ? ? ? ? ? ? ? ?Muraenosaurus 2 1 0 1 2 0 1 0 0 0 0 0 0 0 x 0 0 0 0Peloneustes 1 2 1 1 1 1 1 2 1 0 1 1 0 x x 0 0 0 1Plesiosaurus 0 0 2 1 0 0 0 0 0 0 0 0 0 0 ? 1 0 0 0Pliosaurus 1 2 ? ? ? 1 ? 2 2 0 1 1 0 x x 0 0 0 ?Polycotylus ? 2 1 1 0 ? 1 ? ? ? ? ? ? ? ? ? ? ? ?Rhomaleo. megaceph. 0 0 0 1 0 0 0 1 1 0 0 1 0 0 x 1 0 0 1Rhomaleo. victor 1 0 0 1 2 0 0 1 1 ? ? 0 ? ? ? ? ? ? ?Rhomaleo. zetland. 0 2 ? ? 0 1 ? 1 1 0 1 0 1 x ? 0 0 0 1Simolestes 1 2 1 1 1 0 1 1 1 0 1 0 ? x x 0 ? ? ?Styxosaurus 2 1 ? ? ? 0 0 0 0 0 2 0 ? x ? 0 1 0 ?Thalassiodracon 0 0 0 1 0 0 0 1 1 0 0 0 0 0 0 1 0 0 1Tricleidus 2 0 ? 1 0 2 1 0 0 0 0 0 0 0 x ? ? 0 ?Trinacromerum 1 2 1 1 1 1 1 2 0 0 1 0 0 x x 0 ? 0 ?————————————————————————————————————————————————

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 57
————————————————————————————————————————————————20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41————————————————————————————————————————————————
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1 00 0 0 0 0 1 0 0 0 0 1 0 0 0 1 1 0 0 0,1 0 0 01 0 0 0 0 2 0 1 0 1 1 0 0 0 ? 1 0 0 1 0 1 01 1 0 0 0 2 1 1 1 1 2 0 0 0 1 2 x 0 x 1 1 11 0 0 0 0 2 0 1 0 1 1 0 0 0 ? 1 0 ? 0 ? 0 01 ? 0 0 0 2 ? 1 ? 1 0 0 ? 0 ? 1 0 1 ? 0 0 ?1 0 0 0 0 2 0 1 1 1 0 0 1 0 1 1 1 0 0 1 0 01 1 0 0 0 2 1 1 0 1 2 0 0 0 1 2 x 0 x 1 1 11 1 0 0 0 2 1 1 0 1 2 0 0 1 1 2 x 0 x 1 1 01 0 0 0 1 2 1 1 0 1 2 0 0 0 1 2 x 0 x 1 1 21 0 0 0 0 2 0 1 0 1 ? 0 ? 0 1 ? ? 0 ? 0 ? ?? ? ? ? ? ? ? ? ? 1 ? ? ? 0 1 ? ? ? ? ? ? 01 1 ? 0 0 2 1 1 ? 1 ? ? ? 1 1 ? x ? ? ? 1 ?? ? ? ? ? ? 0 1 ? 1 ? ? 1 ? 1 ? ? ? ? ? ? ?1 0 0 0 0 2 0 1 0 1 0 0 ? 0 1 ? ? ? ? 0 ? 11 1 0 0 ? 2 1 ? 1 1 2 0 0 0 1 2 x 0 x 1 1 ?1 0 0 0 0 2 0 1 0 1 0 0 1 0 1 1 0 0 0 0 0 01 0 0 0 0 2 0 1 ? 1 0 0 1 0 1 1 0 1 0 0 ? ?1 1 ? 0 0 2 1 1 ? 1 ? 0 ? 0 1 2 x 0 x ? 1 11 0 0 0 0 2 1 1 0 1 2 0 0 0 1 2 x 0 x 1 1 2? ? ? ? ? ? ? 1 1 1 ? ? ? ? 1 ? x ? ? ? x ?1 1 ? 0 0 2 1 1 0 1 2 0 0 ? ? ? x 0 ? ? 1 ?1 0 0 0 0 2 0 1 0 1 0 0 1 0 1 1 0 0 0 0 0 01 0 0 0 0 2 0 1 0 1 1 0 0 0 1 2 0 0 x 1 1 11 0 0 0 0 2 ? 1 1 1 0 0 1 0 1 1 1 0 ? ? 0 0? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? x ? ? ? ? ?1 0 0 0 0 2 0 1 ? 1 0 0 ? 0 1 1 0 1 0 0 ? 1? ? ? 0 0 ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?1 0 0 0 0 2 0 1 ? 1 0 0 1 0 1 1 0 1 0 0 0 ?1 0 0 0 0 2 0 1 ? 1 0 0 1 0 1 1 0 ? 0 ? ? ?? ? 0 0 0 2 1 1 1 1 2 0 0 0 1 2 x 0 x 1 1 11 0 0 0 0 2 0 1 0 1 1 0 0 0 1 1 0 0 0 0 1 11 0 0 0 0 2 1 1 ? 1 2 0 0 1 1 2 x 0 x ? 1 01 ? 0 0 1 2 1 1 0 1 2 0 0 0 1 2 x 0 x 1 1 2
————————————————————————————————————————————————Continued

O’Keefe • ACTA ZOOL. FENNICA No. 21358
————————————————————————————————————————————————Taxon 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60————————————————————————————————————————————————Simosaurus 0 0 0 0 x 0 x x 0 0 0 x 0 x 0 0 0 0 0Cymatosaurus ? ? 1 1 ? ? ? ? ? ? ? ? 0 x ? ? 2 0 0Pistosauridae ? ? 1 ? ? ? ? ? ? ? 1 ? 0 0 ? ? ? 0 0Calawayasaurus 0 0 1 1 0 2 0 1 1 1 ? ? 0 0 1 1 1 1 0Attenborosaurus ? ? 1 ? 0 ? ? ? ? ? ? ? 1 1 ? ? 2 0 ?BMNH R.5488 0 1 1 ? 1 ? 0 ? ? ? 0 0 ? 1 1 0 1 ? ?Brachauchenius ? ? 1 ? 1 ? 0 ? ? ? 1 1 1 1 ? ? 1 0 0Brancasaurus 0 0 1 ? 0 ? 0 ? ? ? ? ? 0 0 1 1 1 ? 0Cryptoclidus 1 1 1 ? 0 1 0 0 ? 0 ? ? 1 0 1 1 1 0 2Dolichorhynchops 0 0 1 1 0 1 1 1 0 0 0 0 1 0 1 1 1 0 2Eurycleidus 0 1 1 1 0 0 ? ? 0 0 ? ? 1 1 1 0 1 0 2Hauffiosaurus ? 1 ? ? ? ? ? ? 0 0 ? ? 1 ? ? ? 2 0 0Kimmerosaurus 1 1 1 1 0 0 0 0 ? 0 ? ? 1 0 1 1 1 0 2Kronosaurus ? 1 1 1 0 2 0 1 0 0 ? ? ? 1 ? ? 2 0 0Leptocleidus 0 ? 1 ? ? ? ? 1 0 ? 0 0 1 1 ? ? 1 1 2Libonectes 0 0 1 1 0 2 0 1 1 1 1 ? 0 0 1 1 1 1 0Liopleurodon 0 1 1 0 1 0 0 1 1 0 0 1 1 1 ? 0 2 0 1Macroplata 0 1 1 1 0 0 0 0 0 0 ? 0 ? 1 ? ? 1 0 0Microcleidus ? ? ? ? ? ? ? ? ? ? ? ? ? 0 ? ? ? 1 0MOR 751 0 0 1 1 1 ? ? ? ? 0 ? ? ? 0 ? 1 1 0 2Morturneria 0 0 ? 1 1 0 ? 1 ? ? ? ? 1 ? ? 1 ? ? ?Muraenosaurus 0 1 1 ? 0 ? 0 ? ? ? ? ? 1 0 ? 1 1 0 2Peloneustes 0 1 1 0 0 0 0 1 1 0 0 1 1 1 1 0 2 0 0Plesiosaurus ? 0 1 ? 0 ? ? ? ? 1 1 1 1 0 0 0 1 0 1Pliosaurus 0 1 1 ? 1 ? ? 1 ? ? 0 0 1 1 ? ? 2 0 ?Polycotylus ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?Rhomaleo. megaceph. ? 1 1 1 1 ? 0 1 0 1 0 0 ? 1 ? ? 1 1 2Rhomaleo. victor ? 0 ? ? ? ? ? ? ? ? ? ? ? ? ? ? 1 1 2Rhomaleo. zetland. ? 1 1 ? 1 1 1 1 0 1 0 0 1 1 ? ? 1 1 2Simolestes 0 1 1 ? 1 0 0 1 ? ? 0 0 1 1 ? 0 1 ? ?Styxosaurus ? ? 1 ? ? ? ? ? ? 1 1 0 0 0 1 1 ? ? ?Thalassiodracon 0 1 1 1 0 0 0 0 0 1 1 ? 1 1 0 0 1 0 2Tricleidus 0 0 1 1 0 1 1 0 0 0 ? ? 1 0 ? ? 1 0 2Trinacromerum ? 0 1 ? 0 1 1 ? 0 ? 0 0 1 0 ? ? 1 1 2————————————————————————————————————————————————
Appendix 2. Continued.

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 59
————————————————————————————————————————————————61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82————————————————————————————————————————————————
0 x x x 0 0 0 x x ? x x x x 0 x 0 0 0 0 0 00 x x x 0 0 ? x x ? x x x x ? x 0 0 0 0 1 01 0 x x 0 ? 0 0 0 ? 1 0 ? 0 ? ? 0 0 0 0 0 01 1 0 0 0 1 0 0 0 0 1 2 1 ? 0 0 0 0 0 0 1 01 0 1 x 0 0 0 0 1 0 0 2 0 0 0 0 1 ? ? ? ? 11 1 1 ? 0 0 0 0 0 0 0 2 x 0 ? 0 ? ? ? ? ? ?1 1 1 0 0 0 0 0 3 0 2 2 x 0 ? 0 ? 0 ? 1 ? 11 ? ? 0 ? ? 0 0 ? 0 ? ? 1 0 ? 0 0 ? 0 0 1 ?1 0 0 x 0 1 0 0 0 1 0 2 1 0 0 0 0 0 0 0 ? 01 2 0 0 0 1 1 1 0 1 0 2 1 1 0 1 0 0 0 0 1 01 0 1 x 0 0 0 0 0 0 2 2 0 0 0 0 ? ? 0 0 ? ?1 1 1 0 1 0 0 0 2 0 2 2 x 0 ? 0 0 0 0 0 1 01 0 0 x 0 ? 0 1 0 ? 0 2 1 1 ? 0,1 ? ? ? 0 ? ?1 1 ? 0 ? ? ? 0 2 ? ? ? x 0 ? 0 0 ? ? ? ? ?1 1 1 1 0 0 0 0 0 0 1 2 x 0 0 0 1 ? ? 0 1 11 2 0 0 0 1 0 0 0 0 1 2 1 ? 0 0 0 ? 0 0 1 01 1 1 0 0 0 0 0 3 0 2 2 x 0 ? 0 0 1 1 1 1 11 0 1 x ? ? 0 0 0 0 2 2 0 0 ? 0 ? ? 0 0 1 ?1 0 1 0 0 ? 0 0 0 0 ? 2 1 0 0 ? ? 0 ? ? ? 01 1 0 0 0 1 1 1 0 1 ? 2 1 ? ? ? 0 ? ? ? ? 01 ? 0 x ? ? 0 ? ? 1 ? 2 ? 1 0 1 ? ? 0 ? 1 ?1 0 0 x 0 1 0 0 0 1 0 1 1 0 0 0 0 0 1 0 0 01 1 1 0 1 0 0 0 2 0 2 2 x 0 0 0 1 0 0 0 1 11 0 0 x 0 0 0 0 0 0 ? 1 ? 0 0 0 0 ? 0 0 0 01 1 ? 0 ? ? 0 ? 3 ? ? ? x ? ? 0 ? ? 1 ? 1 ?? ? ? 0 ? ? ? ? ? ? ? ? 1 1 ? ? ? ? ? ? ? ?1 1 1 1 0 0 0 0 0 0 1 2 x 0 0 0 ? 1 0 0 1 11 1 1 1 0 ? 0 0 ? ? 1 1 x ? ? 0 ? 1 1 0 1 01 ? 1 ? 0 ? 0 0 0 0 ? 2 x 0 ? ? ? ? ? ? ? ?1 1 1 ? 0 0 0 0 0 0 ? 2 x 0 ? 0 ? 1 ? ? ? ?? ? ? 0 ? ? ? 0 ? ? ? ? ? ? ? ? 0 ? ? ? ? ?1 0 1 x 0 0 0 0 0 0 2 2 0 0 0 0 0 ? ? 0 1 ?1 0 0 x 0 1 0 1 0 1 0 2 1 1 0 1 ? ? ? 0 0 ?1 2 0 0 0 ? 1 1 0 1 0 2 1 1 ? 1 0 0 ? ? ? 0
————————————————————————————————————————————————Continued

O’Keefe • ACTA ZOOL. FENNICA No. 21360
————————————————————————————————————————————————Taxon 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101————————————————————————————————————————————————Simosaurus 0 0 x 0 0 0 0 0 0 0 0 ? 0 0 0 0 0 x 0Cymatosaurus 1 1 0 0 ? ? 2 ? ? ? ? ? ? ? ? ? 1 ? 1Pistosauridae 0 1 ? 0 ? ? 1 ? ? ? ? ? ? ? ? ? 0 x 1Calawayasaurus 1 0 x 1 1 0 1 0 0 0 0 ? 0 0 0 1 0 x 0Attenborosaurus ? 0 x 0 0 1 2 1 ? 0 0 ? ? 0 0 0 1 ? 0BMNH R.5488 ? ? ? 1 ? 1 2 1 ? ? ? ? ? ? ? 0 1 ? ?Brachauchenius 1 1 0 0 0 ? 2 1 ? 1 ? ? 0 ? 0 0 1 ? 0Brancasaurus 1 ? ? 0 ? 0 ? ? ? ? ? ? ? ? ? ? 0 x 0Cryptoclidus 0 0 x 0 1 0 0 0 0 0 0 0 1 0 1 0 0 x 0Dolichorhynchops 0 0 x 0 x 0 3 2 0 0 0 0 1 0 1 1 0 x 0Eurycleidus 0 0 x 0 1 1 2 1 ? ? 0 ? 0 0 0 0 1 1 ?Hauffiosaurus 1 1 1 0 ? 0 3 1 0 ? ? ? ? ? 0 0 1 1 0Kimmerosaurus ? 0 x ? ? 0 0 0 ? 0 1 0 1 ? 1 1 ? ? 0Kronosaurus ? 1 ? ? ? 0 3 ? ? ? ? ? ? ? ? ? ? ? ?Leptocleidus 0 1 0 1 ? ? 2 ? 0 1 0 1 0 0 0 0 1 1 1Libonectes 1 0 x ? 1 0 1 0 1 0 0 0 0 0 0 1 0 x 0Liopleurodon 1 1 1 0 1 0 2 1 0 1 0 1 0 0 0 0 1 1 0Macroplata 1 0 x 0 0 0 3 1 ? 1 ? ? ? 0 0 0 ? ? 0Microcleidus ? 0 x 0 1 0 1 0 ? 0 ? 0 0 ? 0 1 0 x 0MOR 751 ? 0 x 0 x 0 2 1 0 0 0 ? 1 0 1 1 0 x 0Morturneria ? ? ? 0 1 0 ? ? ? ? ? 0 ? ? 0 1 0 x 0Muraenosaurus 1 0 x 0 1 0 0 0 0 0 0 0 1 0 1 0 0 x 0Peloneustes 1 1 2 0 0 0 3 1 0 1 0 1 0 0 0 0 1 1 0Plesiosaurus 0 0 x 0 ? 0 0 ? ? ? ? 0 0 ? 0 0 0 x 0Pliosaurus 1 1 ? 0 0 0 2 1 0 1 0 1 1 0 0 0 1 1 0Polycotylus ? ? ? ? x 0 ? ? 0 ? 0 0 1 0 1 1 ? ? ?Rhomaleo. megaceph. ? 1 1 1 0 1 2 1 1 ? ? ? ? ? 0 0 1 ? 1Rhomaleo. victor 0 1 ? ? 0 1 2 1 1 ? ? ? ? 0 0 0 ? ? 1Rhomaleo. zetland. ? ? ? 1 0 1 2 1 ? ? 0 ? 0 0 0 0 1 ? 1Simolestes ? 1 ? ? 0 1 2 1 ? 1 0 1 0 0 0 0 ? ? 1Styxosaurus ? ? ? 1 ? 0 1 ? ? 0 ? ? 0 ? ? 1 0 x 0Thalassiodracon 1 0 x 0 ? 0 2 1 ? ? ? ? 0 ? 0 0 0 x 0Tricleidus 0 0 x 0 1 0 0 0 0 0 0 0 ? 0 1 1 0 x 0Trinacromerum ? 0 x 0 ? 0 3 2 ? 0 ? 0 1 0 1 1 0 x 0————————————————————————————————————————————————
Appendix 2. Continued.

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 61
————————————————————————————————————————————————102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124————————————————————————————————————————————————
0 0 0 ? 0 0 ? ? ? 0 0 0 0 0 0 0 0 x 0 0 0 ? 01 0 0 ? 0 0 ? ? ? ? ? ? 0 0 ? ? ? x ? ? ? ? ?0 ? 0 ? 0 0 ? ? ? 0 0 0 0 0 0 0 1 0 1 1 0 0 01 0 0 ? 0 0 ? ? ? 3 1 1 ? 0 1 1 1 0 2 1 0 ? 00 1 0 0 0 0 ? ? ? 0 0 0 ? 0 0 0 1 ? 2 ? 0 0 00 1 0 0 1 ? 0 1 0 0 2 0 1 0 0 0 1 0 1 ? 0 ? 00 1 0 ? ? ? ? ? ? 2 2 ? ? 0 0 1 1 1 2 ? 0 ? 00 0 0 ? 0 ? ? 1 0 1 1 ? ? 0 0 1 1 0 2 1 0 ? 00 0 0 0 1 0 1 0 1 0 0 1 0 0 0 1 1 0 2 0 1 1 10 0 0 ? 0 0 1 1 2 2 2 1 ? 0 0 1 1 0 2 1 0 ? 10 0 0 ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?0 0 0 ? 2 1 ? ? ? 0 0 0 ? 0 0 0 ? 0 2 ? 0 ? ?0 2 0 ? 3 2 1 ? ? ? 2 ? 0 0 0 1 1 0 2 ? 1 1 ?0 1 ? ? ? ? ? 1 ? 2 2 ? ? ? ? 1 1 ? 2 ? 0 ? 00 1 0 0 0 0 ? ? ? 2 2 0 1 0 0 1 1 0 2 0 0 ? 01 0 0 ? 0 0 1 ? 0 3 1 1 ? 1 1 1 1 0 2 1 1 ? 00 1 0 1 0 1 ? ? ? 2 2 0 0 0 0 0 1 1 2 1 0 1 00 0 0 0 ? 1 0 ? 0 0 0 ? 1 0 0 0 1 0 2 0 0 ? 0? 0 0 0 ? ? 0 ? ? 1 1 1 0 1 1 0 1 0 2 ? 1 0 01 1 0 ? 1 0 1 1 2 2 2 ? 1 0 0 1 1 0 ? ? 0 ? ?0 2 0 ? 2 2 0 0 2 ? 2 ? ? 0 1 1 1 0 2 ? 0 ? ?0 0 0 0 0 0 1 0 1 1 1 0 0 0 ? 1 1 0 2 1 1 1 10 1 0 0 1 1 0 0 0 2 2 0 1 0 0 0 1 0 2 0 0 1 00 0 0 0 0 ? ? ? 0 1 0 ? 0 0 0 0 1 0 2 1 0 0 00 1 1 1 1 1 ? ? ? ? 2 ? 0 0 0 0 1 1 2 ? 0 ? 0? 1 0 ? ? ? 1 1 2 2 2 0 ? 0 0 1 1 0 1 1 0 ? 10 1 0 0 0 ? ? ? ? 0 2 ? 1 0 0 0 1 0 1 ? 0 0 00 1 0 ? ? ? ? ? ? 0 ? 0 ? 0 0 ? ? 0 ? ? 0 0 ?0 1 0 0 ? ? ? ? ? 2 2 0 1 0 0 0 1 0 1 ? 0 ? 00 1 0 0 0 ? ? ? 0 2 2 ? 0 0 0 0 1 0 1 1 0 1 01 0 0 ? 0 ? ? ? ? 3 1 ? ? 1 1 1 1 0 2 1 1 ? 00 0 0 0 0 0 0 0 0 0 0 0 1 0 0 0 1 0 2 0 0 0 00 0 0 0 0 ? 1 ? 1 0 0 1 0 0 0 1 1 0 2 1 0 1 10 0 0 ? 0 1 1 1 2 2 2 1 ? 0 0 1 1 0 2 1 0 ? 1
————————————————————————————————————————————————Continued

O’Keefe • ACTA ZOOL. FENNICA No. 21362
————————————————————————————————————————————————Taxon 125 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143————————————————————————————————————————————————Simosaurus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 x x x 0Cymatosaurus ? 0 ? ? ? ? ? ? ? ? ? ? ? ? ? x ? ? ?Pistosauridae 0,1 1 0 0 1,2 0 ? 1 0 ? 0 0 0 1 1 x 0 0 0Calawayasaurus 1 1 1 1 2 1 ? 0 0 0 1 0 0,1 1 1 0 1 1 1Attenborosaurus 0 1 1 1 0 0 ? 0 0 0 0 0 0 1 1 0 0 1 1BMNH R.5488 0 1 1 1 ? 0 ? 0 0 0 0 0 1 1 1 0 0 0 1Brachauchenius 1 1 1 1 2 0 ? ? ? ? ? ? ? ? ? ? ? ? ?Brancasaurus 0 1 1 1 2 1 1 0 0 0 1 ? 2 1 1 0 1 1 1Cryptoclidus 1 1 1 1 2 1 ? 0 0 0 1 1 2 1 1 0 0 1 1Dolichorhynchops 1 1 1 1 2 0 x 0 0 2 0 1 1 1 1 1 0 1 1Eurycleidus ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? 0 ? ? 1Hauffiosaurus ? ? 1 1 ? 0 ? 0 ? ? 0 0 ? 1 1 0 0 1 1Kimmerosaurus 1 ? ? 1 ? 0 ? ? ? ? ? ? ? ? ? ? ? ? ?Kronosaurus 0 ? 1 1 2 0 ? ? ? ? 0 ? 0 1 1 ? 0 ? ?Leptocleidus 0 1 1 1 ? 0 0 0 0 0 0 0 1 1 1 0 0 1 ?Libonectes 1 ? 1 1 ? 1 ? 0 0 0 2 0 2 1 1 ? 1 1 ?Liopleurodon 0 1 1 1 2 0 ? 0 ? ? 0 ? ? 1 1 0 0 1 1Macroplata 0 ? 1 1 0 0 ? ? ? ? ? ? ? ? 1 ? ? ? 1Microcleidus 1 1 1 1 2 1 ? 0 ? ? 1 ? 2 ? 1 0 0 1 1MOR 751 ? ? 1 1 ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?Morturneria ? ? 1 1 ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?Muraenosaurus 1 1 1 1 2 1 ? 0 0 1 1 1 2 1 1 0 0 1 1Peloneustes 0 1 1 1 2 0 0 0 ? ? 0 0 0 1 1 0 0 1 1Plesiosaurus 1 1 1 1 2 1 ? 0 1 0 0 0 1 1 1 0 0 1 1Pliosaurus 0 1 1 1 ? 0 ? ? ? ? ? ? ? ? ? 0 ? ? 1Polycotylus 1 1 1 1 2 0 x 0 0 2 0 1 1 1 1 1 ? ? 1Rhomaleo. megaceph. 0 1 1 1 2 0 ? 0 ? ? ? ? ? ? 1 ? 0 ? 1Rhomaleo. victor ? ? ? 1 2 ? ? ? 0 ? 0 0 1 1 1 0 0 1 ?Rhomaleo. zetland. ? 1 1 1 ? 0 ? ? ? ? ? ? ? ? ? ? ? ? ?Simolestes ? 1 1 1 ? 0 ? 0 ? ? 0 ? 0 1 1 0 0 0 1Styxosaurus 1 ? 1 1 2 1 ? ? ? ? ? ? ? ? ? ? ? ? ?Thalassiodracon 0 1 1 1 2 0 ? 0 0 0 0 0 1 1 1 0 0 1 1Tricleidus 1 1 1 1 2 0 0 0 0 2 1 0 2 1 1 0 0 1 1Trinacromerum 1 1 1 1 2 0 x 0 0 2 0 1 1 1 1 1 0 1 1————————————————————————————————————————————————
Appendix 2. Continued.

ACTA ZOOL. FENNICA No. 213 • A cladistic analysis and taxonomic revision of the Plesiosauria 63
————————————————————————————————————————————————144 145 146 147 148 149 150 151 152 153 154 155 156 157 158 159 160 161 162 163 164 165 166————————————————————————————————————————————————
0 0 x 0 0 0 0 0 x 0 x x 0 0 0 1 2 0 0 0 0 0 11 ? ? 1 ? ? 0 0 x ? x x ? ? ? 0 1 ? ? ? ? ? 0? 1 x ? 1 1 0 0 x 0 x x 1 1 0 1 ? 0 0 0 ? ? 00 1 0 1 1 1 1 1 0 0 1 0 1 1 0 1 2 1 0 1 1 1 ?1 1 0 1 1 1 0 1 0 0 0 0 1 1 1 1 2 0 0 1 1 0 01 1 1 1 1 1 0 1 ? 0 0 0 1 1 1 1 2 0 ? ? 1 ? 0? ? ? ? ? ? 1 1 ? ? 0 1 ? ? 0 ? ? 1 ? 1 1 ? ?0 1 1 1 1 1 1 1 ? ? 0 0 1 ? 0 1 2 1 ? 1 1 ? 01 1 0 1 1 1 1 1 1 0 0 0 1 1 0 1 2 1 1 1 1 0 01 1 0 1 1 1 1 1 1 0 0 0 1 1 0 1 2 1 3 1 1 1 ?1 1 0 1 1 1 ? 1 ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?1 1 ? 1 1 ? 1 1 0 0 0 0 1 1 1 1 2 0 2 1 1 0 0? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?1 1 1 1 1 ? ? 1 ? ? 0 0 ? ? 0 1 2 1 ? ? 1 ? ?? ? ? ? ? ? 1 1 0 0 ? 0 1 ? ? ? ? ? ? ? 1 ? ?? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?1 1 1 1 1 1 1 1 0 0 0 0 1 1 0 1 2 1 0 1 1 0 01 1 1 1 1 ? 1 1 0 0 0 0 1 1 ? 1 2 0 ? ? 1 ? ?1 1 1 1 1 1 1 1 1 0 0 0 1 1 1 1 2 0 1 1 1 0 0? ? ? ? ? ? 1 1 1 1 0 0 1 1 0 1 2 1 ? 1 1 1 ?? ? ? ? ? ? 1 1 ? ? ? ? ? ? ? ? ? 1 ? ? 1 ? ?1 1 0 1 1 1 1 1 1 1 0 0 1 1 0 1 2 1 1 1 1 0 01 1 0 1 1 1 1 1 0 0 0 0 1 1 0 1 2 1 0 1 1 0 01 1 1 1 1 1 0 1 0 0 0 0 1 1 1 1 2 0 1 1 1 0 01 1 0 1 1 ? 1 1 0 0 0 1 1 1 0 1 2 1 ? ? 1 ? 01 1 0 1 1 1 1 1 1 1 0 0 1 1 0 1 2 1 3 1 1 ? 01 1 ? 1 1 1 0 1 0 0 0 0 1 1 1 1 2 0 ? ? 1 ? 01 1 1 1 1 ? 0 1 0 0 0 0 1 1 1 1 2 0 3 1 1 0 0? 1 ? ? ? ? 0 1 0 0 0 0 1 1 1 1 2 0 ? ? 1 ? ?1 1 1 1 1 1 1 1 1 0 0 0 1 1 0 1 2 0 0 ? 1 ? ?? ? ? ? ? ? ? 1 ? 0 ? ? ? ? ? ? ? ? ? ? ? ? ?1 1 1 1 1 1 0 1 0 0 0 0 1 1 1 1 2 0 0 1 1 0 01 1 0 1 1 ? 1 1 1 1 0 0 1 1 0 1 2 1 2 1 1 ? 01 1 0 1 1 1 1 1 1 0 0 0 1 1 0 1 2 1 3 1 1 1 0
————————————————————————————————————————————————









![A TAXONOMIC REVISION OF THE GENUS LIPPIA … · A TAXONOMIC REVISION OF THE GENUS LIPPIA [HOUST. EX] LINN. (VERBENACEAE)* IN AUSTRALIA Ahmad Abid Munir ... number (2n = 40 (42?))](https://static.fdocuments.us/doc/165x107/5b88c1117f8b9abf5c8c31fc/a-taxonomic-revision-of-the-genus-lippia-a-taxonomic-revision-of-the-genus-lippia.jpg)








