
A cellular mechanis fomr the palatal shelf reorientation from a … · Palatal shelf reorientation...
14
/. Embryo/, exp. Morph. Vol. 53, pp. 1-13, 1979 Printed in Great Britain © Company of Biologists Limited 1979 A cellular mechanism for the palatal shelf reorientation from a vertical to a horizontal plane in hamster: light and electron microscopic study By RAVINDRAM. SHAH 1 From the Department of Oral Biology, Faculty of Dentistry, The University of British Columbia SUMMARY Cellular and subcellular events during reorientation of the palatal shelf in hamster fetuses are described. An alteration in the morphology of the epithelial and the mesenchymal cells is observed during shelf realignment from a vertical to a horizontal plane. The mesenchymal cells elongate and subsequently appear to protrude in the medially bulging palatal shelf. Microtubules, microfilaments and close contacts are associated with elongation of the mes- enchymal cells. The cells of the thickened epithelium may play a mechanical role in providing direction to the mesenchymal cells during palatal shelf reorientation. The altered morphology of the mesenchymal cells may be associated with the intrinsic shelf force implicit in Walker and Fraser's theory of palatal shelf reorientation. INTRODUCTION Following the publication of Dursy's study in 1869 on the development of the secondary palate, one of the much discussed questions was how the vertical palatine processes, at first separated by the tongue, are able to move up to a horizontal position. Several possibilities were suggested and disputed regard- ing the mechanism underlying reorientation of the palatal shelf (see Ferguson, 1978 for details). However, the mechanism still remains unclear. A literature review of the electron microscopic observations of palatal develop- ment in rodents and human fetuses indicates that most studies were concerned with differentiation and fate of midline epithelial cells after the palatal shelves were realigned from a vertical to a horizontal plane. However, few studies describing ultrastructural aspects of both the epithelial and the mesenchymal cells during palatal shelf reorientation have been made (Walker, 1961; Babiarz, Allenspach & Zimmerman, 1975; Innes, 1978; Ferguson, 1978). These were carried out in mice and rats and the results were conflicting. Shah & Travill (1976 a) observed that the reorientation of palatal shelves from a vertical to a horizontal plane in hamster fetuses occurred between days 1 Author's address: Department of Oral Biology, Faculty of Dentistry, The University of British Columbia, Vancouver, B.C. V6T 1W5, Canada.
Transcript of A cellular mechanis fomr the palatal shelf reorientation from a … · Palatal shelf reorientation...

/ . Embryo/, exp. Morph. Vol. 53, pp. 1-13, 1979Printed in Great Britain © Company of Biologists Limited 1979
A cellular mechanism for the palatal shelfreorientation from a vertical to a horizontal planein hamster: light and electron microscopic study
By RAVINDRAM. SHAH1
From the Department of Oral Biology, Faculty of Dentistry, The Universityof British Columbia
SUMMARYCellular and subcellular events during reorientation of the palatal shelf in hamster fetuses
are described. An alteration in the morphology of the epithelial and the mesenchymal cells isobserved during shelf realignment from a vertical to a horizontal plane. The mesenchymalcells elongate and subsequently appear to protrude in the medially bulging palatal shelf.Microtubules, microfilaments and close contacts are associated with elongation of the mes-enchymal cells. The cells of the thickened epithelium may play a mechanical role in providingdirection to the mesenchymal cells during palatal shelf reorientation. The altered morphologyof the mesenchymal cells may be associated with the intrinsic shelf force implicit in Walkerand Fraser's theory of palatal shelf reorientation.
INTRODUCTION
Following the publication of Dursy's study in 1869 on the development ofthe secondary palate, one of the much discussed questions was how the verticalpalatine processes, at first separated by the tongue, are able to move up toa horizontal position. Several possibilities were suggested and disputed regard-ing the mechanism underlying reorientation of the palatal shelf (see Ferguson,1978 for details). However, the mechanism still remains unclear.
A literature review of the electron microscopic observations of palatal develop-ment in rodents and human fetuses indicates that most studies were concernedwith differentiation and fate of midline epithelial cells after the palatal shelveswere realigned from a vertical to a horizontal plane. However, few studiesdescribing ultrastructural aspects of both the epithelial and the mesenchymalcells during palatal shelf reorientation have been made (Walker, 1961; Babiarz,Allenspach & Zimmerman, 1975; Innes, 1978; Ferguson, 1978). These werecarried out in mice and rats and the results were conflicting.
Shah & Travill (1976 a) observed that the reorientation of palatal shelvesfrom a vertical to a horizontal plane in hamster fetuses occurred between days
1 Author's address: Department of Oral Biology, Faculty of Dentistry, The University ofBritish Columbia, Vancouver, B.C. V6T 1W5, Canada.

2 R. M. SHAH12:00 and 12:04 (12 days 4 h) of gestation. They further noted that the modeof palatal shelf reorientation in hamster fetuses resembled that in humans(Anderson & Matthiessen, 1967; Aronov, 1970) and mice (Walker & Fraser,1956; Greene & Kochhar, 1973). The purpose of the present investigation is tostudy cellular and subcellular events taking place at the time of change in positionof the palatal shelf from a vertical to a horizontal plane in the hamster fetus.Further, an effort will be made to relate the observations of the present studyto those reported in the literature to propose a hypothetical mechanism of shelfreorientation.
MATERIALS AND METHODS
Maintenance and breeding of Golden Syrian hamster have been describedelsewhere (Shah & Travill, 1976a). The pregnant hamsters, in a group of five,were killed at 1 h intervals between days 12:00 and 12:04 of gestation. Half thefetuses from each litter were fixed in Bouin's solution for light microscopy andthe remainder in 3 % phosphate-buffered cold glutaraldehyde for electronmicroscopy. Fetuses for light microscopy were processed for parafin embedding.Frontal serial sections, 7/tm thick, were stained with hematoxylin and eosin. Themethod for processing glutaraldehyde fixed palates for electron microscopy wassimilar to one described by Shah & Travill (19766). Prior to embedding, thepalates were divided into anterior, middle and posterior thirds and carefullyoriented to procure 1 jam frontal sections. Thin sections were obtained from themiddle third of the secondary palate and stained with methanolic uranylacetate (Stempak & Ward, 1964) followed by lead citrate (Reynolds, 1963). Thesections were observed in a Philips 300 or Hitachi HU-11E-1 electronmicroscope.
RESULTS
Light microscopy
The vertical palatal shelf (Fig. 1) was composed of mesenchymal tissuecovered by one to three cell layers of epithelium (Fig. 2). Examination of serialsections revealed a focal epithelial thickening on the medial aspect of the verticalshelf. The epithelial thickening was localized and was seen only in the middlethird of the secondary palate. The thickening was composed of three to fivelayers of cells. The palatal mesenchyme was composed of stellate cells scatteredin an amorphous ground substance. An occasional cell undergoing mitosis wasseen in the mesenchyme. The epithelium on the medial aspect of the verticalshelf, i.e. the~prospective midline epithelium that is destined to fuse and dis-integrate, showed no mitotic figures.
Reorientation of the shelf from a vertical to a horizontal plane started in themiddle third of the secondary palate (Fig. 3). In the frontal section, a medialbulge was observed above the area of epithelial thickening (Fig. 4). The prospec-tive midline fusion epithelium showed no histological change from that of the

Palatal shelf reorientation in hamster
Fig. I. Ventral view of hamster palate at day 12:00 of gestation. Both the lower jawand tongue are removed. The palatal shelves are in vertical position, x 16.Fig. 2. A vertical palatal shelf at day 12:00 of gestation showing thickening of theepithelium on its medial aspect (arrow). Epon thick section, x 76.Fig. 3. Ventral view of hamster palate at day 12:02 of gestation. Both the lower jawand tongue are removed. The palatal shelves are reorienting in the middle third butare still vertical in the posterior third, x 16.Fig. 4. A reorienting palatal shelf at day 12:02 of gestation. Some of the mes-enchymal cells are elongated (arrows). Note the epithelial thickening (E). Epon thicksection. x52.

R. M. SHAH
Figs 5-7. For legends see facing page.

Palatal shelf reorientation in hamster 5
vertical shelf. Some of the mesenchymal cells were, however, elongated andappeared to be polarized along the transverse axis of the medially bulging palatalshelf. Other mesenchymal cells were stellate or appeared to be undergoingelongation.
Subsequently, when the palatal shelf became horizontal (Fig. 5), the focalthickening of the epithelium was on its medial edge (Fig. 6). The mesenchymalcells were either stellate or elongated and resembled those of the vertical orreorienting shelf. However, as the opposing horizontal shelves approachedone another toward the midline all mesenchymal cells were stellate.
Electron microscopy
The epithelium covering the medial aspect of the vertical palatal shelf(Fig. 7) was separated from the underlying mesenchyme by a continuous basallamina. Numerous spaces were present between the epithelial cells. The super-ficial epithelial cells were flat. They were attached to one another and to thesubjacent epithelial cells by desmosomes. They contained a flattened nucleussurrounded by polyribosomes, mitochondria, a few cisternae of rough endo-plasmic reticula and bundles of tonofilaments. The basal cells were roughlycuboidal. Each contained a relatively large, oval or irregularly shaped nucleus,polyribosomes, a few cisternae of rough endoplasmic reticula, mitochondria,coated vesicles and a small Golgi complex.
The cells of focal epithelial thickening (Fig. 8) varied in their shapes. The cellsnear the basal lamina were generally cuboidal and those away were flat. A fewspaces intervened between the cells of thickened epithelia which were otherwiseattached to one another by desmosomes. The cells in thickened epithelium con-tained a spherical or oval nucleus surrounded by numerous polyribosomes, a smallGolgi complex, few strands of rough endoplasmic reticula and mitochondria.
The mesenchymal cells were stellate and contained a large nucleus surroundedby few cytoplasmic organelles and a well developed Golgi complex.
During reorientation of the palatal shelf (Fig. 9), the basal lamina remained
F I G U R E S 5-7
Fig. 5. Ventral view of hamster palate at day 12:04 of gestation. Both the lower jawand tongue are removed. The palatal shelves are horizontal, x 16.Fig. 6. The horizontal palatal shelves at day 12:04 of gestation. The thickened epi-thelium is present on the medial edge of the shelves (arrows). Epon thick section.x42.Fig. 7. Electron micrograph of a vertical palatal shelf at day 12:00 of gestation.A continuous basal lamina (BL) separates the epithelium from the mesenchyme.Numerous spaces (JCS) are present between the superficial and the basal cells andbetween the latter. The superficial cell is characterized by a flat nucleus (N) sur-rounded by polyribosomes, mitochondria (M) and tonofilaments (Tf). The basalcell, in addition, contains a small Golgi complex (GC) and a few cisternae of roughendoplasmic reticula (RER). The nucleus (N) in the basal cell is irregular. Thesuperficial cell is attached to the basal cell by desmosomes (D). x 13200.

R. M. SHAH
Fig. 8. Electron micrograph of a vertical palatal shelf at day 12:00 of gestation.Thickened epithelium. The cells near the basal lamina (BL) are cuboidal and thoseaway are flat. Intercellular spaces (ICS) are few and the cells are attached to oneanother by desmosomes (D). The cells of thickened epithelium contain a sphericalor oval nucleus (N), numerous polyribosomes, rough endoplasmic reticulum (RER)and mitochondria (M). x 8800.
continuous. The spaces between the epithelial cells were increased both innumber and size from that of the earlier stage. The basal cells appeared to haveflattened as compared to that of the vertical shelf. Lysosomes appeared for thefirst time in both the superficial and the basal cells. Other cytoplasmic featuresremained unchanged.
The thickened epithelium on the reorienting palatal shelf (Fig. 10), on theother hand, showed a reduction in number and size of the spaces between itscells. Also the cells near the basal lamina were roughly columnar rather thancuboidal as seen on the vertical shelf. Other cells were flat. The Golgi complexwas well developed. As in the other epithelial cells, lysosomes were recognizedfor the first time in the cells of the thickened epithelium.

Palatal shelf reorientation in hamster
Fig. 9. Electron micrograph of a reorienting palatal shelf at day 12:02 of gestationshowing numerous intercellular spaces (ICS) in the epithelium. The basal lamina(BL) is intact. Lysosomes (Ly) are present in both the superficial and the basal cells,x 8800.
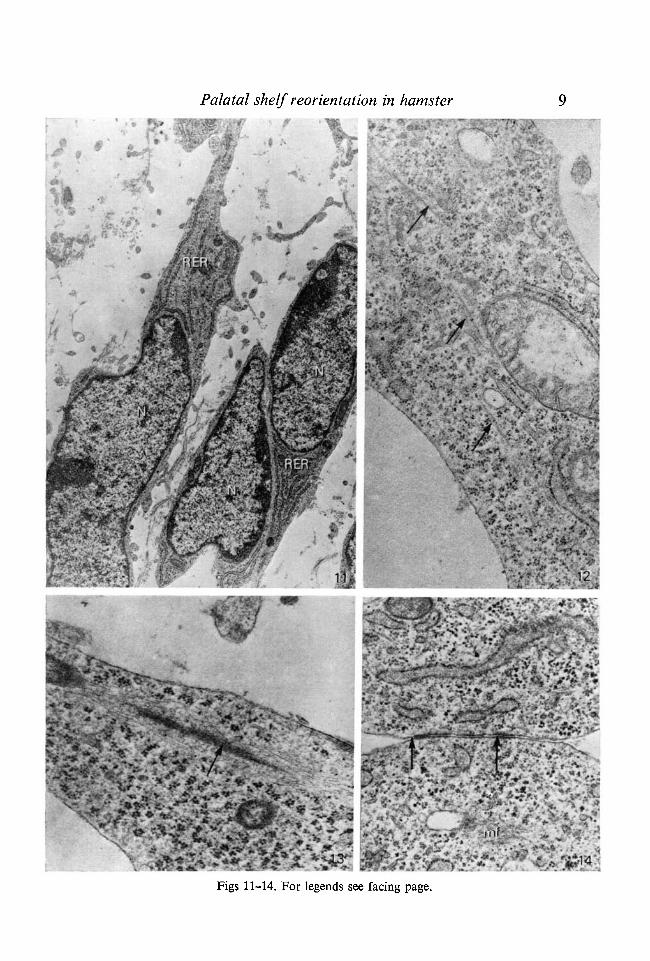
The mesenchymal cells of reorienting palatal shelves were elongated (Fig. 11).The nuclei of the elongated cells were flattened. In addition, microtubules(Fig. 12) and condensation of microfilaments (13) were also present. Often twoneighbouring elongated cells were attached to one another by close junctions(Fig. 14).
The epithelium covering the horizontal palatal shelf was separated from thesubjacent mesenchyme by a continuous basal lamina. The spaces between theepithelial cells were reduced in number and size. The basal cells were roughlycuboidal. The cytoplasm showed an increase in organelles. The morphology andcontent of the thickened epithelium remained unchanged.
DISCUSSION
The mechanism by which the palatal shelf changes its position from a verticalto a horizontal plane is not understood. External factors such as muscularpressure by the tongue (Humphrey, 1969, 1971; Walker, 1969, 1971) and analteration at the cranial base (Harris, 1967; Taylor, 1978) have been consideredto be responsible for the shelf reorientation. However, palatal closure has been

R. M. SHAH
Fig. 10. Electron micrograph of a reorienting palatal shelf at day 12:03 of gestationshowing few intercellular spaces (ICS) (compare with Fig. 9) in the thickenedepithelium. The basal cells (Bas) are roughly columnar (compare with Fig. 8).Mitochondria (M), Golgi complex (GC), rough endoplasmic reticulum (RER),nucleus (N), lysosome (Ly). x 8800.
F I G U R E S 11-14
Fig. 11. Electron micrograph of a reorienting palatal shelf at day 12:02 of gestation.The elongated mesenchymal cells contain a flattened nucleus (N), polyribosomesand numerous cisternae of rough endoplasmic reticula (RER). x 8800.Fig. 12. Electron micrograph of a reorienting palatal shelf at day 12:03 of gestation.The elongated mesenchymal cell contains microtubules (arrows) which are orientedalong the long axis of the cell, x 31000.Fig. 13. Electron micrograph of a reorienting palatal shelf at day 12:03 of gestation.The elongated mesenchymal cell contains microfilaments (arrow) which are orientedalong the long axis of the cell, x 32300.Fig. 14. Electron micrograph of a reorienting palatal shelf at day 12:03 of gestation.The two neighbouring mesenchymal cells are attached to one another by closejunctions (arrows). Microfilaments (mf). x 28600.
Palatal shelf reorientation in hamster
Figs 11-14. For legends see facing page.

10 R. M. SHAH
noted in human cases of aglossia and microglossia (see review by Shah, 1977)and no substantial alterations at the cranial base have been noted at the time ofshelf reorientation (Hart, Smiley & Dixon, 1972; Diewert, 1974). Further,in vitro elevation of shelves has also been observed in the absence of the tongueand cranial base (Brinkley, Basehoar, Branch & Avery, 1975; Wee, Wolfson &Zimmerman, 1976). Thus both clinical and experimental observations seem toindicate that external factors may not be necessary for shelf elevations.
Walker & Fraser (1956) have suggested that reorientation of the palatalshelves by the mode of medial bulging may be due to an 'intrinsic shelf force'.The nature of such a force, if any, is still undetermined. The contention thatan increased acid mucopolysaccharide concentration in the palatal shelf at thetime of elevation may provide the necessary internal shelf force (Walker, 1961;Larsson, 1962; Jacobs, 1964; Pratt, Goggins, Wilk & King, 1973; Ferguson,1978) has been disputed (Nanda, 1971; Andrew & Zimmerman, 1971).Suggestions that the intrinsic shelf force may reside in the elastic fibers (Walker& Fraser, 1956; Clark, 1956) have been discarded by subsequent histochemicaland ultrastructural observations in which no evidence of elastic fibers in the shelftissue was found (Walker, 1961; Isaacson & Chaudhry, 1962; Frommer &Monroe, 1969). Suggestions that an increase in cellular proliferation within thepalatal shelf tissue at the time of elevation may be the source of the shelf force(Schorr, 1908; Mott, Toto & Hilgers, 1969; Jelinek & Dostal, 1974; Nanda &Romeo, 1975) have also been disputed (Walker & Fraser, 1956; Cleaton-Jones,1976). Indeed it has been recently indicated that such growth does not playa direct role in the shelf reorientation (Greene & Pratt, 1976).
The most recent hypothesis to explain the shelf reorientation through intrinsicmechanism is the molecular interactions of the contractile proteins, actin andmyosin. These proteins are synthesized in the mesenchymal cells just prior toreorientation (Lessard, Wee & Zimmerman, 1974) and are represented bya cytoplasmic filament system (Babiarz et ah 1975; Innes, 1978). The results ofthe present study are in accordance with this hypothesis. In addition, ourfindings showed that there is an alteration in the morphology of both the mesen-chymal and the epithelial cells during palatal shelf reorientation. Themesenchymal cells undergo elongation and appear to protrude into the mediallybulging palatal shelf. The epithelial cells also flatten and appear to adaptthemselves to the subjacent growth forces. These morphological alterations maybe associated with the necessary power implicit in Walker & Fraser's intrinsicforce theory.
The alterations in the morphology of mesenchymal cells coincide with theappearance of microtubules and microfilaments. Microtubules play an importantrole in the determination of cell shape during organogenesis (Byers & Porter,1964; Handel & Roth, 1971), and thus may have a similar role in elongationof mesenchymal cells. Presence of microfilaments in non-muscle cells is con-sidered to be associated with cellular movement during morphogenesis (Wessels

Palatal shelf reorientation in hamster 11
et al. 1971; Pichichero & Avers, 1973) since they contain contractile proteins(Goldman, Lazarides, Pollak & Weber, 1975). Ultrastructural identification ofmicrofilaments in the elongated mesenchymal cells of the palate, as observed inthe present study, may correspond to the contractile proteins identified byLessard et al. (1974) and Krawczky & Gillon (1976) in mouse palate. Morpho-logically, however, they do not resemble the myofibrils described by Babiarzet al. (1975) and Innes (1978). The lack of resemblance may be due to differencesin the areas of observation. In the present study observations were made in themiddle third of the hard palate whereas the other authors observed the musclecells in the posterior part of the palate. In any case, the presence of cytoplasmicfilaments in palate mesenchyme may permit marshalling of internal stressesfor movement of mesenchymal cells, and thus in general our results supportthe hypothesis that internal shelf force may reside in the mesenchymal cells ofthe palatal shelf.
Close contacts observed between the elongated mesenchymal cells duringreorientation of the palatal shelves may also be important in generating internalshelf force since Dehaan (1963), Hay (1968) and Sanders (1973) have shown thatsuch contacts play an important role in cellular movement during morphogenesis.
An interesting finding during the present study was occurrence of focalepithelial thickening on the medial aspect of the vertical palatal shelf. The cellsin the thickening were closely packed. The reorientation of the palatal shelfstarted immediately above the thickened epithelium. A similar thickening ofepithelium, however, has not been reported during mouse or human palataldevelopment and its significance is, therefore, entirely speculative. The epithelialthickening may provide necessary guidance for the elongated mesenchymal cellsto move into the medially bulging palatal shelf during the reorientation stage.The focal epithelial thickening could then act as a 'mechanotactic' pivot aroundwhich the mesenchymal cells can glide. Similar mechanotactic guidance has alsobeen suggested during corneal development (Nelson & Revel, 1975).
In brief, the foregoing Results and Discussion indicate that reorientation ofthe hamster palatal shelf occurs through the mode of medial bulging and thusresembles that of humans and mice. A transient elongation of the mesenchymalcells, which subsequently appears to protrude in the medially bulging shelf, maybe associated with generation of the intrinsic force for the palatal shelf re-alignment. Microtubules, microfilaments and close contacts are all associatedwith the morphological changes of the mesenchymal cells. The epithelial changesmay be adaptive and guiding in nature.
The work was supported by a grant from the Medical Research Council of Canada. Theauthor extends his gratitude to Mrs V. Koulouris and Miss Ruth Scheuing for theirassistance.

12 R. M. SHAH
REFERENCES
ANDERSON, H. & MATTHIESSEN, M. (1967). Histochemistry of the early development of thehuman face and nasal cavity with special reference to the movement and fusion of thepalatine processes. Acta Anat. 68, 473-508.
ANDREW, F. D. & ZIMMERMAN, E. F. (1971). Glucocorticoid induction of cleft palate inmice: No correlation with inhibition of mucopolysaccharide synthesis. Teratology 4, 31-38.
ARONOV, A. (1970). Development of the soft and hard palate in man. IX InternationalCongress of Anatomy, 70-73.
BABIARZ, B. S., ALLENSPACH, A. L. & ZIMMERMAN, E. F. (1975). Ultrastructural evidence ofcontractile systems in mouse palates prior to rotation. Devi Biol. 47, 32-44.
BRINKLEY, L., BASEHOAR, G., BRANCH, A. & AVERY, J. (1975). New in vitro system for study-ing secondary palatal development. J. Embryol. exp. Morph. 34, 485-495.
BYERS, B. & PORTER, K. R. (1964). Oriented microtubules in elongating cells of the developinglens rudiment after induction. Proc. natn. Acad. Sci., U.S.A. 52, 1091-1099.
CLARK, K. H. (1956). Histological investigation of cortisone induced cleft palate in mice.Genetics 41, 637-638.
CLEATON-JONES, P. (1976). Radioautographic study of mesenchymal cell activity in thesecondary palate of the rat. / . dent. Res. 55, 437-441.
DEHAAN, R. L. (1963). Oriented cell movements in embryogenesis. In Biological Organizationat the Cellular and Supracellular Level (ed. R. J. C. Harris), pp. 147-165. New York:Academic Press.
DIEWERT, V. M. (1974). A cephalometric study of orofacial structures during secondarypalate closure in the rat. Archs Oral Biol. 19, 303-315.
DURSY, E. (1869). Zur Entwicklungsgeschichte des kopfes des Menshen und der hoherenwirbeltiere. Verlag der H. Lauppschen Buchhandlung, Tubigen.
FERGUSON, M. W. J. (1978). Palatal shelf elevation in the wistar rat fetus. / . Anat. 125,555-577.
FROMMER, J. & MONROE, C. (1969). Further evidence for the absence of elastic fibers duringmovement of the palatal shelves in mice. / . dent. Res. 48, 155-156.
GOLDMAN, R. D., LAZARIDES, E., POLLAK, R. & WEBER, K. (1975). The distribution of actinin non-muscle cells. The use of actin antibody in the localization of actin within themicrofilament bundles of mouse 3T3 cells. Expl Cell Res. 90, 333-344.
GREENE, R. M. & KOCHHAR, D. M. (1973). Spatial relations in the oral cavity of cortisonetreated mouse fetuses during the time of secondary palate closure. Teratology 8, 153-162.
GREENE, R. M. & PRATT, R. M. (1976). Developmental aspects of secondary palate formation.J. Embryol. exp. Morph. 36, 225-245.
HANDEL, M. A. & ROTH, L. E. (1971). Cell shape and morphology of the neural tube:Implications for microtubule function. Devi Biol. 25, 78-95.
HARRIS, J. W. S. (1967). Experimental studies on closure and cleft formation in the secondarypalate. Sci. Basis. Med. Ann. Rev. 354-370.
HART, J., SMILEY, G. & DIXON, A. D. (1972). Sagittal growth trends of the cranio-facialcomplex during formation of the secondary palate in mice. Teratology 6, 43-50.
HAY, E. D. (1968). Organization and fine structure of epithelium and mesenchyme in thedeveloping chick embryo. In Epithelial-Mesenchymal Interactions (ed. R. Fleischmajer &R. E. Billingham), pp. 31-55. Baltimore: Williams & Wilkins Co.
HUMPHREY, T. (1969). The relation between human fetal mouth opening reflexes and closureof the palate. Amer. J. Anat. 125, 317-344.
HUMPHREY, T. (1971). Development of oral and facial motor mechanisms in human fetusesand their relation to craniofacial growth. / . dent. Res. 50, 1428-1441.
INNES, P. B. (1978). The ultrastructure of the mesenchymal element of the palatal shelves ofthe fetal mouse. / . Embryol. exp. Morph. 43, 185-194.
ISAACSON, R. & CHAUDHRY, A. P. (1962). Cleft palate induction in strain A mice withcortisone. Anat. Rec. 142, 479-486.
JACOBS, R. M. (1964). Histochemical study of morphogenesis and teratogenesis of the palatein mouse embryos. Anat. Rec. 149, 691-698.

Palatal shelf reorientation in hamster 13JELINEK, R. &DOSTAL, M. (1974). Morphogenesis of cleft palate induced by exogenous factors.
VII. Mitotic activity during formation of the mouse secondary palate. Folia Morph. 22,94-101.
KRAWCZYK, W. & GILLON, D. (1976). Immunofluorescent detection of actin in non-musclecells of the developing mouse palatal shelf. Archs Oral Biol. 21, 503-508.
LARSSON, K. S. (1962). Studies on the closure of the secondary palate. III. Autoradiographicand histochemical studies in the normal mouse embryo. Acta morph. neer.-scand. 4,349-367.
LESSARD, J. L., WEE, E. L. & ZIMMERMAN, E. F. (1974). Presence of contractile proteins inmouse fetal palate prior to shelf elevation. Teratology 9, 113-126.
MOTT, W., TOTO, P. & HILGERS, D. (1969). Labelling index and cellular density in palatineshelves of cleft palate mice. / . dent. Res. 48, 263-269.
NANDA, R. (1971). The role of sulfated mucopolysaccharides in cleft palate formation.Teratology 3, 237-244.
NANDA, R. & ROMEO, D. (1975). Differential cell proliferation of embryonic rat palatalprocesses as determined by incorporation of tritiated thymidine. Cleft Palate J. 12, 436-442.
NELSON, G. A. & REVEL, J. P. (1975). Scanning electron microscopic study of cell movementsin the corneal endothelium of the avian embryo. Devi Biol. 42, 315-333.
PICHICHERO, M. E. & AVERS, C. J. (1973). The evolution of cellular movement in eukaryotes:The role of microfilaments and microtubules. Sub-cell. Biochem. 2, 97-105.
PRATT, R. M., GOGGINS, J. F., WILK, A. L. & KING, C. T. G. (1973). Acid mucopoly-saccharide synthesis in the secondary palate of the developing rat at the time of rotationand fusion. Devi Biol. 32, 230-237.
REYNOLDS, E. S. (1963). The use of lead citrate at high pH as an electron opaque stain inelectron microscopy. /. Cell Biol. 17, 208-213.
SANDERS, E. J. (1973). Intercellular contact in the unincubated chick embryo. Z. Zellforsch.Mikrosk. Anat. 141, 459-468.
SCHORR, G. (1908). Zur Entwicklungsgeschichte der sekundaren gaumens bei cinigensaugetierer und beim menschen. Anat. Hefte. 36, 69-104.
SHAH, R. M. (1977). Palatomandibular and maxillo-mandibular fusion, partial aglossia andcleft palate in a human embryo. Report of a case. Teratology 15, 261-272.
SHAH, R. M. & TRAVILL, A. A. (1976a). Morphogenesis of the secondary palate in normaland hydrocortisone treated hamsters. Teratology 13, 71-84.
SHAH, R. M. & TRAVILL, A. A. (19766). Light and electron microscopic observations onhydrocortisone induced cleft palate in hamsters. Amer. J. Anat. 145, 149-166.
STEMPAK, J. G. & WARD, R. T. (1964). An improved staining method for electron microscopy.J. Cell Biol. 22, 691-10\.
TAYLOR, G. R. (1978). Craniofacial growth during closure of the secondary palate in thehamster. / . Anat. 125, 361-370.
WALKER, B. E. (1961). The association of mucopolysaccharides with morphogenesis of thepalate and other structures in mouse embryos. / . Embryol. exp. Morph. 9, 22-31.
WALKER, B. E. (1969). Correlation of embryonic movement with palatal closure in mice.Teratology 2, 191-198.
WALKER, B. E. (1971). Palate morphogenesis in the rabbit. Archs Oral Biol. 16, 275-286.WALKER, B. E. & FRASER, F. C. (1956). The embryology of cortisone induced cleft palate.
J. Embryol. exp. Morph. 5, 201-209.WEE, E., WOLFSON, L. & ZIMMERMAN, E. (1976). Palate shelf movement in mouse embryo
culture: Evidence for skeletal and smooth muscle contractility. Devi Biol. 48, 91-103.WESSELS, N. K., SPOONER, B. S., ASH, J. F., BRADLEY, M. O., LUDUENA, M. A., TAYLOR,
E. L., WRENN, J. T. & YAMADA, K. M. (1971). Microfilaments in cellular and develop-mental processes. Science, N.Y. Ill, 135-143.
(Received 22 November 1978, revised 22 March 1979)
EMB 53



















