
Ultrasensitive and rapid detection of ochratoxin A in agro ...
Upload
alessandro-bottonCategory
view
215download
2
International Journal of Food Microbiology 127 (2008) 105–115
Contents lists available at ScienceDirect
International Journal of Food Microbiology
j ourna l homepage: www.e lsev ie r.com/ locate / i j foodmicro
A cDNA-AFLP approach to study ochratoxin A production in Aspergillus carbonarius
Alessandro Botton a, Davide Ferrigo b, Cristina Scopel b, Roberto Causin b, Claudio Bonghi a, Angelo Ramina a,⁎a Department of Environmental Agronomy and Crop Science, University of Padova, Campus of Agripolis — Viale dell’Università 16, 35020 Legnaro, Padova, Italyb T.E.S.A.F. — Dip. Territorio e Sistemi Agro-Forestali, University of Padova, Campus of Agripolis — Viale dell’Università 16, 35020 Legnaro, Padova, Italy
⁎ Corresponding author. Fax: +39 049 8272850.E-mail address: [email protected] (A. Ramina)
0168-1605/$ – see front matter © 2008 Elsevier B.V. Aldoi:10.1016/j.ijfoodmicro.2008.06.037
a b s t r a c t
a r t i c l e i n f oArticle history:
Aspergillus carbonarius is re Received 17 February 2008Received in revised form 23 April 2008Accepted 19 June 2008Keywords:Aspergillus carbonariusOchratoxin A biosynthesiscDNA-AFLP differential displayG proteinCa2+/calmodulin dependent phosphorylationand dephoshorylationZinc finger transcription factors
sponsible for the majority of mycotoxin contaminations in grapes and its deri-vatives. Most of A. carbonarius strains are ochratoxin A (OTA) producers, even though at very different levels.This broad variability was used to identify genes whose expression is linked with the ability of producingOTA. A cDNA-AFLP differential display screening was performed in two strains of A. carbonarius, antagonistsfor the ability of producing OTA, allowing the identification of 119 differentially expressed sequences puta-tively involved in the regulation of OTA biosynthesis. A likely connection was pointed out between thebiosynthesis of the toxin, vegetative growth and sexual/asexual developmental progression, along withcommon signalling pathways involving G protein and Ca2+/calmodulin dependent phosphorylation anddephoshorylation cascades.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
Ochratoxins (OTs) are secondary metabolites produced by severalspecies of filamentous fungi belonging to the genera Aspergillus, Pe-tromyces, Neopetromyces and Penicillium (Perrone et al., 2006). Thebasic chemical structure ofOTs consists of an isocoumarin group linkedto a phenylalanine moiety by an amide bond. Ochratoxin A (OTA), thechlorinated form, is the most toxic and the most prevalent, whereasochratoxin B and C, the non-chlorinated and esterified forms, respec-tively, are less common and toxic (Bayman and Baker, 2006). OTAcontamination may be affected by different factors such as tempera-ture, water activity, pH and substrate composition. Therefore the toxi-genicity of an ochratoxigenic fungus may vary according to the cropand the geographical area. In temperate locations, OTA is mainly pro-duced by Penicillium species contaminating cereals, whereas in thetropical regions it is synthesized mainly by A. ochraceus. Two otherspecies, namely A. niger var. niger and A. carbonarius, are responsiblefor themajorityof contaminations in cereals and grapes (Battilani et al.,2003; Ponsone et al., 2007; Atoui et al., 2007a), respectively, in thewarm zones (Ringot et al., 2006).
The OTA biosynthetic pathway has not yet been completely eluci-dated. So far, the majority of the studies have been focused on Peni-cillium species (Färber and Geisen, 2004) and A. ochraceus (Harris andMantle, 2001), pointing out that the phenylalanine moiety originatesfrom the shikimic acid pathway, whereas the isocoumarin is formedfrom acetate units via the pentaketide pathway. Polyketide synthase
.
l rights reserved.
(PKS) is the key enzyme catalyzing the first step of OTA biosynthesisand the related gene (pks) has been previously cloned and character-ized in different species, showing a high degree of sequence variability(O'Callaghan et al., 2003; Varga et al., 2003; Geisen et al., 2004). A pksgene was isolated also in A. ochraceus and shown to be expressed onlyunder OTA permissive conditions. Moreover, insertional mutagenesisleading to a disruption of the pks gene impaired OTA production(O'Callaghan et al., 2003). The following steps of OTA biosynthesisinclude the formation of the lactone ring originating mellein, the ad-dition of a carboxyl group to form ochratoxin β (OTβ), and the chlo-rination by means of a chloroperoxidase synthesizing ochratoxin α(OTα). Finally, the linking of OTα to phenylalanine is carried out bymeans of the enzyme OTA synthetase. Inhibition of OTA productioncan be achieved with various natural and synthetic compounds, in aspecies-dependent manner. This may indicate that the biosynthesis ofthis mycotoxin is not conserved in all the producing fungi (Baymanand Baker, 2006).
Additional studies were focused also on the biosynthetic regula-tion of many other mycotoxins, such as aflatoxins, fumonisins, sterig-matocystin and tricothecenes (Brodhagen and Keller, 2006), andnumerous signalling pathways, most of which are shared by differentmycotoxins and diverse species, were outlined. A strong link betweensexual and asexual sporulation and mycotoxin biosynthesis was alsoevidenced. Particularly, the G protein signal transduction pathway hasbeen shown to regulate both secondary metabolism and sporulation(Han et al., 2004), along with lipid signalling (Tsitsigiannis and Keller,2006). Several transcription factors have been shown to bind pro-moter elements of genes involved in mycotoxin biosynthesis and se-condary metabolism, as for Zn(II)2Cys6 zinc finger proteins (Ehrlich

106 A. Botton et al. / International Journal of Food Microbiology 127 (2008) 105–115
et al., 1999) and Cys2His2 (Borneman et al., 2001). Important regu-latory roles involve also the Ca2+/calmodulin-dependent phosphor-ylation/dephosphorylation pathways (Rao and Subramanyam, 2000;Jayashree et al., 2000). However, few reports concerning theregulation of OTA biosynthesis are available (Basìlico and Basìlico,1999; Mitchell et al., 2004; Bellì et al., 2007; O'Callaghan et al., 2003;Pardo et al., 2006; Pateraki et al., 2007; Valero et al., 2007a,b), and thegenetic characterization was mainly focused on either Penicilliumspecies (Färber and Geisen, 2004) or the genes directly involved in thebiosynthetic pathway (O'Callaghan et al., 2003; Varga et al., 2003).Moreover, further studies were carried out only for diagnostic pur-poses, to allow the identification of OTA-producing species contam-inating foods and feeds (Geisen et al., 2004; Schmidt et al., 2004; Bauet al., 2005; Niessen et al., 2005; Dao et al., 2005; Patiño et al., 2005;Bogs et al., 2006; Perrone et al., 2006; Zanzotto et al., 2006; Atoui et al.,2007b; Martìnez-Culebras and Ramòn, 2007). Since OTA represents achallenging problem both for food and feed producers and consumers,the full elucidation of the biosynthetic pathway and its regulationwould allow the setting up of innovative tools to control and/or pre-vent the crop contamination. In this view, biocompetition strategiesusing atoxigenic strains have revealed to be a possible solution, as inthe case of non-toxigenic isolates of A. flavus successfully used to re-duce aflatoxin contamination of cottonseed, corn and peanuts (Ehrlichand Cotty, 2004b).
Among the commodities that can be contaminated by OTA, grapesandwines are thosewith themajor content of toxin, next only to cereals(Bau et al., 2005). Wine is an important beverage in the world trade, ofwhich France, Italy and Spain are themainproducers and exporters. Dueto the actual importance of the wine industry, several studies werecarried out to assess the relevance of OTA contamination in wine. Morethan 50% of Spanish wines were shown to be contaminated, with thehighest OTA concentration in dessert wines (12.25 ng/mL) (Lòpez deCerain et al., 2002; Bellì et al., 2004; Blesa et al., 2004;Mateo et al., 2006;Hernàndez et al., 2006). In Italy, 78.4% of red wines were contaminated,with a maximum of 7.63 ng/mL (Visconti et al., 2000; Pietri et al., 2001;Brera et al., 2005; Bacaloni et al., 2005). In the other countries, thecontamination of wines by OTA is less serious (Mateo et al., 2007).
A. carbonarius is the main responsible organism for the majority ofcontaminations in grapes and its derivatives (Battilani et al., 2003;Ponsone et al., 2007; Atoui et al., 2007a). Most of A. carbonarius strainsare OTA producers, even though at very different levels (Bau et al.,2005; Perrone et al., 2006). This broad variability may be successfullyused to characterize the genetic regulation of OTA biosynthesis, iso-lating the genes differentially expressed in high- and low-producingisolates.
Differential display techniques, such as Differential Display ReverseTranscriptase-PCR (DDRT-PCR) and microarrays, were successfully usedfor aflatoxin biosynthesis studies in A. flavus and A. parasiticus (OBrianet al., 2003), as well as for OTA biosynthesis studies in P. nordicum(Färber and Geisen, 2004). The differential conditions were achieved bygrowth on different minimal media, either supporting or inhibiting OTAproduction. However, with such an approach, several genes were dif-ferentially expressed because of the different media, generating con-fusion in the interpretation of results.
Among the differential display techniques, cDNA-AFLP has provedto be the most popular procedure because of its ability in displayingseveral transcripts simultaneously and detecting differentially ex-pressed genes (Bachem et al., 1996; Botton et al., 2008). Both reliabilityand sensitivity of amplification products proved to be very high, andexpression patterns visualized by cDNA-AFLP well correlated withnorthern blot analysis (Durrant et al., 2000; Jones et al., 2000; Donsonet al., 2002; Cnudde et al., 2006; Albertini et al., 2004).
In the present research, cDNA-AFLP differential display was used toisolate genes differentially expressed in OTA high- and low-producingstrains of A. carbonarius. In the present study, two strains previouslycharacterized by Perrone et al. (2006), namely ITEM 5005 and ITEM
5012, were used because of their differential production of OTA. Thesequences isolated were analyzed for homology and classified ac-cording to the gene ontology criteria (Ashburner et al., 2000). Finally, acomparison between the annotations of up- and down-regulatedsequences was carried out to highlight the differentially expressedfunctional categories. The involvement of such genes in the OTA bio-synthesis is critically assessed and a possible model for its regulationproposed.
2. Materials and methods
2.1. Strains, media and growth conditions
Fungal strains ITEM 5005 and ITEM 5012 of A. carbonarius wereselected from a list of 20 strains obtained from Agri-Food ToxigenicFungi Culture Collection (I.S.P.A. — Institute of Sciences of Food Pro-duction, Bari, Italy) and characterized for OTA production by Perroneet al. (2006).
Strains were grown on Petri dishes containing Czapek DOX Agar(OXOID Ltd., Basingstoke, Hampshire, England) in dark condition at25 °C for 6 days to achieve spore production. Spores were collectedwith a sterile solution of 0.1% (v/v) Tween 20 (J.T. Baker, Deventer,Holland) and stored at −20 °C in 25% (v/v) of glycerol (Sigma-AldrichChemie Gmbh, Steinheim, Germany) before use. Monoconidial cul-tures of A. carbonarius were obtained as reported by Tuite (1969).Monoconidial isolates were subcultured in YES liquid medium (2%yeast extract, 15% sucrose, 1 g/L MgSO4) pH 6.5 (O'Callaghan et al.,2003) in motionless and dark condition at 25 °C for 14 days and OTAproduction monitored by ELISA assays (see below) to detect theoptimum growth phase to collect the mycelia. An aliquot of culturebroth containing conidia (about 110mL) was collected for each isolate,inoculated into 250 mL Erlenmeyer flasks containing 100 mL of YESliquid medium, and grown as described above for 7 days. During thisphase the isolates showed an exponential increase of OTA production.The mycelia were harvested by filtration through a 0.45 μm Milliporefilter (Millipore Corporation, USA), immediately frozen in liquid ni-trogen and then stored at −80 °C before nucleic acid extraction.
2.2. Quantification of OTA production
ELISA determination of OTA content in A. carbonarius isolates wasperformed on 5 mL of liquid culture for each strain. Samples wereblended, extracted in 15 mL of a solution 1:2 HCl 1 M:dichloremethan,shaken for 15 min, centrifuged for 15 min at 2200 ×g, and 5 mL oflower organic phase collected and neutralized with 2.5 mL of a 0.13 Msodium bicarbonate solution at pH 8.1. The solutions were shaken andcentrifuged as previously described, the upper aqueous phase con-taining OTA collected and quantified using the I'screen OCHRA kit forquantitative ELISA (Tecna Diagnostic and Biotechnology, Trieste, Italy)following the manufacturer's instructions. Absorbance was recordedat 450 nm, at room temperature, with a 318MC Micro ELISA platereader (Sanco Instruments Co., Ltd, Shanghai, China). Standard curveswere obtained by plotting the seven standard values of OTA con-centration against optical density at 450 nm. OTA concentration insample isolates was determined from the standard curve.
Themonoconidial isolates 5005E (6 ng/mL) and 5012A (2500 ng/mL),were analyzed by HPLC as described by Varga et al. (2005) to confirmELISA results, and selected for the following cDNA-AFLP analyses.
2.3. RNA extraction and cDNA synthesis
Total RNAwas isolated from50mgofmycelia using theRNAMinispinkit (GE Healthcare, Chalfont St. Giles, UK) following the manufacturerinstructions, quantified with a spectrophotometer (Biomate 3, ThermoScientific, Waltman, WA, USA), and run on 1% agarose gel to check itsintegrity. Twenty µg of total RNAwere treated with 10 U of RQ1 RNase-

107A. Botton et al. / International Journal of Food Microbiology 127 (2008) 105–115
free DNase (Promega, Milan, Italy) and 1 Unit of RNase Inhibitor(RNAguard, Amersham Biosciences, Piscataway, NY, USA) for 30 min,then purified by phenol–chloroform extraction and precipitated inisopropyl-alcohol as described by Sambrook and Russel (2001). Double-stranded cDNAwas synthesized from 10 µg of DNA-free total RNA usingthe Universal Riboclone cDNA Synthesis SystemKit (Promega, Madison,USA) according to the manufacturer's instructions.
2.4. cDNA-AFLP
Double-stranded cDNA was used for cDNA-AFLP analyses as pre-viously described (Vos et al., 1995; Bachem et al., 1996) with somemodifications. Restriction and adaptor ligationwere performed simul-taneously, as described by Barcaccia et al. (2001), in a total volume of30 μL containing 1× One-phor-all buffer (Amersham Biosciences, Pis-cataway, NY, USA), DTT 5 mM, BSA 50 ng/μL, 50 pmol of MseI adaptor,5 pmol of EcoRI adaptor, ATP 10 mM, 5 Units of MseI and 5 Units ofEcoRI, 1 Unit of T4 DNA Ligase and 10 μg of double-stranded cDNA. Themixture was incubated for 4 h at 37 °C. Preamplification was per-formed in a total volume of 50 μL containing 5 μL of restriction–ligation mixture, 1.5 ng/µL of primers without selective nucleotides,dNTPs 200 μM each, 1× PCR buffer and 1 Unit of Taq polymerase(Amersham Biosciences, Piscataway, NY, USA). PCR conditions were asfollows: an initial hold of 5 min at 94 °C, 25 cycles composed by 30 s at94 °C, 30 s at 55 °C and 60 s at 72 °C, and a final hold of 7 min at 72 °C.The reaction was checked on 1% agarose gel and diluted 1:5 withddH2O. The hot-amplification was carried out on 5 µL of diluted pre-amplification mixture in a total volume of 20 μL with 0.4 Units of Taqpolymerase (Amersham Biosciences, Piscataway, NY, USA), 5 ng of 33P-labeled EcoRI primer, 30 ng of MseI primer, dNTPs 200 μM each and 1×PCR buffer, using the following touch-down profile (Barcaccia et al.,1999): an initial hold of 3 min at 94 °C and 14 cycles composed of 30 sat 94 °C, 30 s at 54.5 °C and 60 s at 72 °C with an annealing tem-perature decrease of 0.7 °C/cycle, followed by 18 cycles composed by30 s at 94 °C, 30 s at 55.2 °C and 60 s at 72 °C, and final hold of 10min at72 °C. The final amplification was performed with 88 different com-binations of EcoRI and MseI primers (Table 1). An equal volume of GelLoading Buffer (98% formamide, EDTA 10 mM, 0.05% w/v of bro-mophenol blue and xylene cyanol) was added to the PCR prior to adenaturing step of 5 min at 98 °C. All the reactions were performed ina GeneAmp PCR system 9700 (Applied Biosystems, Foster City, CA,USA).
The amplification products were separated in a 5% polyacrylamidegel with 8 M urea at 90 W constant power using the SequiGen GTsystem (Biorad, Hercules, CA, USA), dried-blotted onto a Whatman3MM paper at 80 °C for 1 h and visualized by autoradiogram afterovernight exposure on Kodak MR-I X-ray film (Botton et al., 2005). Allexperiments were carried out in triplicates.
Table 1Primers used in the cDNA-AFLP experiments
EcoRI primers MseI primers(Eco = AGACTGCGTACCAATTC) (Mse = GACGATGAGTCCTGAGTAA)
01 Mse+AAGA Eco+AAC 02 Mse+ACTB Eco+ACC 03 Mse+AGCC Eco+AGG 04 Mse+AGGD Eco+CAG 205 Mse+AGTE Eco+CCA 06 Mse+ATAF Eco+GCA 07 Mse+ATCG Eco+TAA 08 Mse+CAAH Eco+TGA 09 Mse+CAC
10 Mse+CAT11 Mse+CCA
All the 88 possible combinations between EcoRI and MseI primers were adopted.
2.5. Purification and sequencing
The autoradiogram films were scanned and analyzed with KODAK1D v 3.6 software (Scientific Imaging Systems, Eastman Kodak Com-pany) to quantify the net intensity of the bands. Amplicons showing atleast a 3-fold difference in terms of intensity between the two samplesin all the replicates were excised from the blots and immersed over-night in 100 μL of PCR-grade water. 5 μL were used in a standard PCRreaction with the same EcoRI and MseI primers used in the pre-amplification step described above. The amplified products were se-parated in a 1.5% agarose gel, and the bands excised and purified bymeans of Minelute Gel Extraction Kit (Qiagen, Milano, Italy). Thesequencing reaction was performed with the Big Dye Terminator v3.1kit (Applied Biosystems, Foster City, CA, USA) on 5 μL of purified bandfollowing the manufacturer's instructions, and analyzed at the BMRGenomics (University of Padova, www.bmr-genomics.com).
2.6. Annotation of sequences
BlastX analyses were performed by means of Blast2GO softwarev2.0.1 (http://www.blast2go.de; Conesa et al., 2005; Aparicio et al.,2006). The parameters were differentiated according to the length ofthe nucleotide records and four ranges were defined for this purpose:0–99 bp, 100–199 bp, 200–399 bp, ≥400 bp. Blast expectation valuethreshold was constantly set to 10, whereas HSP length cut-off was setto 10, 15, 20 and 33, respectively. This configuration was chosen toimprove the E-value scores in the Blast analyses, allowing to retrievedatabase matches also for short sequences with short local alignments(Botton et al., 2008). The remaining parameterswere kept as default. Inparticular, the default Blast remote server (http://www.ncbi.nlm.nih.gov/BLAST/Blast.cgi) was used to achieve updated database matches.The Mapping tool of Blast2GO software was used to obtain geneontology (GO) information from retrieved database matches. Annota-tion of all sequences was performed with different parameters on tworanges of length, 0–199 bp and ≥200 bp. E-value Hot Filter was set to 0and 3, respectively, and GO weight constantly to 5. A furtherdifferential annotation parameter was adopted concerning similarity,setting up the threshold cutoff at 60% and 30% for sequences b200 bpand ≥200 bp, respectively. This configuration allowed a stricterannotation of shorter sequences representing also a filter towardsless reliable BlastX matches. Two following steps were carried out toenrich the annotation. InterProScan (ZdobnovandApweiler, 2001)wasperformed byenabling all the possiblemotif database searches and theresulted GO terms added to the annotation. The annotation analysiswas then implemented with the ‘Augment Annotation by ANNEX’function. Basically, this approach uses univocal relationships betweenGO terms from the different GO categories to add implicit annotation(http://www.goat.no; for a detailed description seeMyhre et al., 2006).TheGOSlimgoslim_yeast.obowas used to achieve specific GO terms bymeans of a fungi-specific reduced version of the gene ontology (http://geneontology.org/GO.slims.html). Statistical analyses to comparesequence sets were performed with the integrated GOSSIP tool of theBlast2GO software.
3. Results
3.1. cDNA-AFLP
The cDNA-AFLP technique allowed to identify a total of 383 dif-ferentially expressed amplicons by using 88 different combinations ofEcoRI/MseI primers. Only the bands showing at least a three-folddifference in terms of intensity between the two antagonist sampleswere excised from the blots and sequenced (Fig. 1). A total of 142sequences with an average length of 313 bp were obtained and suc-cessively clustered with strict parameters to eliminate redundancyresulting equal to 16%. Among the 119 ESTs obtained after clustering, 55

Fig. 1. cDNA-AFLP autoradiogram of nine of the most informative primer combinationsamplifying differentially expressed genes in OTA low- (LP) and high- (HP) producingstrains. The combinations of primers used were indicated according to the codes re-ported in Table 2. The boxes show some of the differentially expressed fragments.
108 A. Botton et al. / International Journal of Food Microbiology 127 (2008) 105–115
were down-regulated and 64 were up-regulated in the low-producing(LP) strain.
3.2. Analysis of differentially expressed genes
A BlastX analysis was performed on the isolated sequences pointingout a 42.9% of clones without similarity with known proteins, distri-buted as37.5%among theup-regulated and49.1% in thedown-regulatedfragments, that were annotated as “unknown function/hypotheti-cal protein” (Tables 2 and 3). The matching sequences showed a meanE-value equal to 2.96×10−1 and an average similarity with blast hits ashigh as 75%. Following the mapping process, during which the softwarerecovers the gene ontology (GO) terms mainly by using the accessionnumbers of the blast hits, three GO terms were assigned on average to
each sequence (data not shown). Enzyme codes (EC)were also found for33 sequences on the total 68 annotated ones (Tables 2 and 3).
Concerning the final annotation sets, the GO terms were groupedaccording to cellular component, molecular function and biologicalprocess, by up- and down-regulated clones separately. In Fig. 2, the piecharts show the gene ontology at level 4 in the case of the biologicalprocess, and at level 3 in the case of the cellular component and mo-lecular function categories. Different levels of ontology were chosenfor being the most informative.
Among the LP strain up-regulated genes, the majority were classifiedas related to the intracellular components, in particular 22% in ‘in-tracellular part’, 22% generally as ‘intracellular’, and 17% as ‘intracellularorganelle’, accounting for a total of 61%. The remaining annotationsconcerned the membrane- and non-membrane-bound organelle, for 13%and 4%, respectively (Fig. 2A, left). Regarding the molecular function, the‘hydrolase activity’ was the GO term most frequently encountered (28%),followed by the transferase (20%), and ligase activities (12%). The re-maining categories spanned from 8% of ‘nucleic acid binding’, ‘signaltransducer activity’, ‘helicase activity’, and ‘protein binding’, to 4% ofoxidoreductase and lyase activities (Fig. 2B, left). As far as the biologicalprocess is concerned, the majority of annotations were related to‘carbohydrate metabolic processes’ as in 20% of the cases, followed by‘nucleobase, nucleoside, nucleotide and nucleic acid metabolic process’(13%), ‘biopolymermetabolic process’ (11%), and other different categoriesof the same level. Among the most interesting were the ‘signal trans-duction’ (4%), ‘lipid metabolic process’ (2%), ‘meiotic cell cycle’ (2%), andthe ‘cell cycle phase’ (2%) terms (Fig. 2C, left).
Concerning the sequences down-regulated in LP, the intracellularcomponent was prevalent also in this case with a total of 68% of GOterms, distributed in 24% for ‘intracellular part’, 24% for ‘intracellular’,and 20% for ‘intracellular organelle’ (Fig. 2A, right). As pointed out inFig. 2B (right), the ‘hydrolase activity’was themost abundant categoryof themolecular function annotations also in this case, reaching 34% ofthe total. It was followedby the ‘oxidoreductase activity’ (24%), ‘nucleicacid binding’ (12%), ‘protein binding’ (12%), and lyase, transferase, andligase activities, equally at 6%. In the biological process classification, amore fragmentary situation was pointed out. ‘Cellular metabolic pro-cess’ (11%), ‘protein metabolic process’ (11%) and ‘biopolymer meta-bolic process’ (12%) were the most represented GO terms. Otherimportant categories were ‘lipid metabolic process’ (9%) and ‘signaltransduction’ (6%). For a more detailed description, see Fig. 2C (right).An enrichment analysis allowed to compare the annotation sets of up-and down-regulated genes, bymeans of a statistical approach pointingout the GO categories for which the two groups significantly differed.The categories showing the best statistical scoreswere ‘cytoplasm’ and‘oxidoreductase activity’, belonging to the cellular component andmolecular function ontologies, respectively, both differing of 5.7%(Fig. 3 and data not shown). Among the other GO terms, the mostinteresting are ‘carbohydrate metabolic process’, ‘lipid metabolic pro-cess’, ‘membrane’, ‘transferase activity’, ‘generation of precursormeta-bolites and energy’, ‘cell cycle’, and ‘signal transducer activity’.
3.3. Genes encoding enzymes putatively involved in the generation and/or metabolism of OTA precursors
All the metabolic pathways involving a potential precursor of OTAwere searched in the KEGG pathway database (www.genome.ad.jp/kegg/pathway.html), and compared with the pathways in which theproteins encoded by the differentially expressed genes may be puta-tively involved. Severalmatcheswere found that are reported inTable 4.
The clone 2.28.07, which was shown to be down-regulated in theLP strain, may be involved in numerous pathways, since it putativelyencodes an aldehyde dehydrogenase. Such enzyme is active in gly-colysis, fatty acid metabolism, lysine degradation, and in metabolismof phenylalanine, β-alanine, and pyruvate. In glycolysis, it is involvedin the interconversion of acetate into acetaldehyde, as well as in the
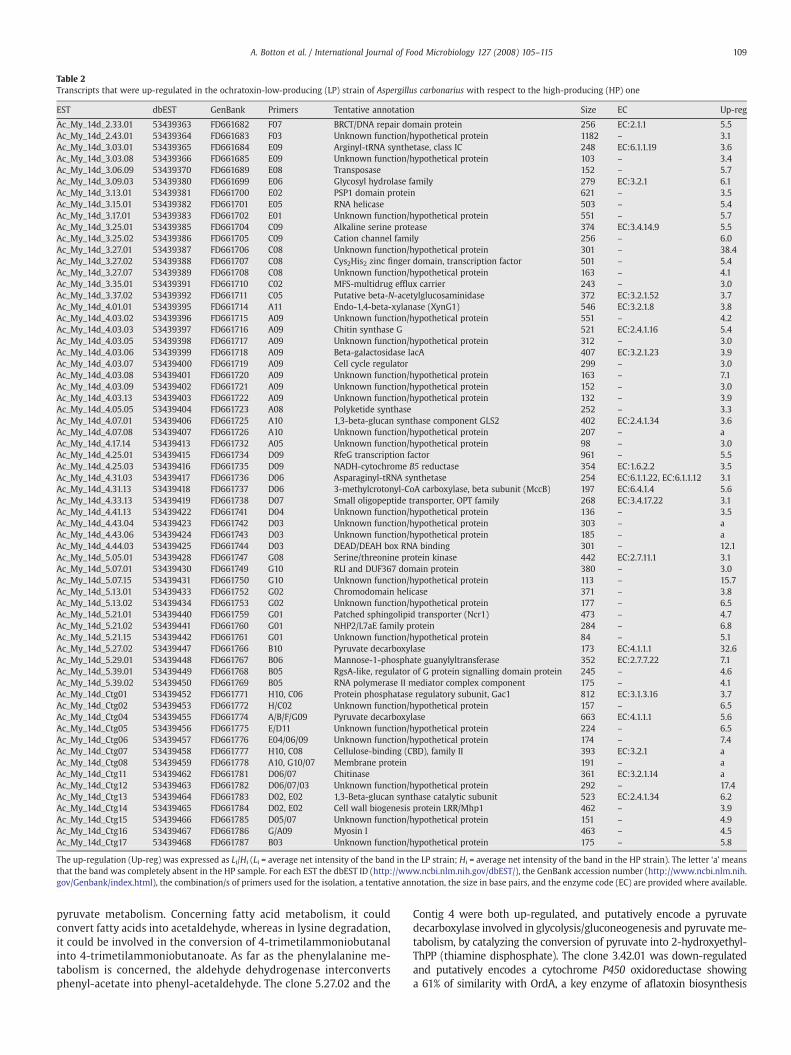
Table 2Transcripts that were up-regulated in the ochratoxin-low-producing (LP) strain of Aspergillus carbonarius with respect to the high-producing (HP) one
EST dbEST GenBank Primers Tentative annotation Size EC Up-reg
Ac_My_14d_2.33.01 53439363 FD661682 F07 BRCT/DNA repair domain protein 256 EC:2.1.1 5.5Ac_My_14d_2.43.01 53439364 FD661683 F03 Unknown function/hypothetical protein 1182 – 3.1Ac_My_14d_3.03.01 53439365 FD661684 E09 Arginyl-tRNA synthetase, class IC 248 EC:6.1.1.19 3.6Ac_My_14d_3.03.08 53439366 FD661685 E09 Unknown function/hypothetical protein 103 – 3.4Ac_My_14d_3.06.09 53439370 FD661689 E08 Transposase 152 – 5.7Ac_My_14d_3.09.03 53439380 FD661699 E06 Glycosyl hydrolase family 279 EC:3.2.1 6.1Ac_My_14d_3.13.01 53439381 FD661700 E02 PSP1 domain protein 621 – 3.5Ac_My_14d_3.15.01 53439382 FD661701 E05 RNA helicase 503 – 5.4Ac_My_14d_3.17.01 53439383 FD661702 E01 Unknown function/hypothetical protein 551 – 5.7Ac_My_14d_3.25.01 53439385 FD661704 C09 Alkaline serine protease 374 EC:3.4.14.9 5.5Ac_My_14d_3.25.02 53439386 FD661705 C09 Cation channel family 256 – 6.0Ac_My_14d_3.27.01 53439387 FD661706 C08 Unknown function/hypothetical protein 301 – 38.4Ac_My_14d_3.27.02 53439388 FD661707 C08 Cys2His2 zinc finger domain, transcription factor 501 – 5.4Ac_My_14d_3.27.07 53439389 FD661708 C08 Unknown function/hypothetical protein 163 – 4.1Ac_My_14d_3.35.01 53439391 FD661710 C02 MFS-multidrug efflux carrier 243 – 3.0Ac_My_14d_3.37.02 53439392 FD661711 C05 Putative beta-N-acetylglucosaminidase 372 EC:3.2.1.52 3.7Ac_My_14d_4.01.01 53439395 FD661714 A11 Endo-1,4-beta-xylanase (XynG1) 546 EC:3.2.1.8 3.8Ac_My_14d_4.03.02 53439396 FD661715 A09 Unknown function/hypothetical protein 551 – 4.2Ac_My_14d_4.03.03 53439397 FD661716 A09 Chitin synthase G 521 EC:2.4.1.16 5.4Ac_My_14d_4.03.05 53439398 FD661717 A09 Unknown function/hypothetical protein 312 – 3.0Ac_My_14d_4.03.06 53439399 FD661718 A09 Beta-galactosidase lacA 407 EC:3.2.1.23 3.9Ac_My_14d_4.03.07 53439400 FD661719 A09 Cell cycle regulator 299 – 3.0Ac_My_14d_4.03.08 53439401 FD661720 A09 Unknown function/hypothetical protein 163 – 7.1Ac_My_14d_4.03.09 53439402 FD661721 A09 Unknown function/hypothetical protein 152 – 3.0Ac_My_14d_4.03.13 53439403 FD661722 A09 Unknown function/hypothetical protein 132 – 3.9Ac_My_14d_4.05.05 53439404 FD661723 A08 Polyketide synthase 252 – 3.3Ac_My_14d_4.07.01 53439406 FD661725 A10 1,3-beta-glucan synthase component GLS2 402 EC:2.4.1.34 3.6Ac_My_14d_4.07.08 53439407 FD661726 A10 Unknown function/hypothetical protein 207 – aAc_My_14d_4.17.14 53439413 FD661732 A05 Unknown function/hypothetical protein 98 – 3.0Ac_My_14d_4.25.01 53439415 FD661734 D09 RfeG transcription factor 961 – 5.5Ac_My_14d_4.25.03 53439416 FD661735 D09 NADH-cytochrome B5 reductase 354 EC:1.6.2.2 3.5Ac_My_14d_4.31.03 53439417 FD661736 D06 Asparaginyl-tRNA synthetase 254 EC:6.1.1.22, EC:6.1.1.12 3.1Ac_My_14d_4.31.13 53439418 FD661737 D06 3-methylcrotonyl-CoA carboxylase, beta subunit (MccB) 197 EC:6.4.1.4 5.6Ac_My_14d_4.33.13 53439419 FD661738 D07 Small oligopeptide transporter, OPT family 268 EC:3.4.17.22 3.1Ac_My_14d_4.41.13 53439422 FD661741 D04 Unknown function/hypothetical protein 136 – 3.5Ac_My_14d_4.43.04 53439423 FD661742 D03 Unknown function/hypothetical protein 303 – aAc_My_14d_4.43.06 53439424 FD661743 D03 Unknown function/hypothetical protein 185 – aAc_My_14d_4.44.03 53439425 FD661744 D03 DEAD/DEAH box RNA binding 301 – 12.1Ac_My_14d_5.05.01 53439428 FD661747 G08 Serine/threonine protein kinase 442 EC:2.7.11.1 3.1Ac_My_14d_5.07.01 53439430 FD661749 G10 RLI and DUF367 domain protein 380 – 3.0Ac_My_14d_5.07.15 53439431 FD661750 G10 Unknown function/hypothetical protein 113 – 15.7Ac_My_14d_5.13.01 53439433 FD661752 G02 Chromodomain helicase 371 – 3.8Ac_My_14d_5.13.02 53439434 FD661753 G02 Unknown function/hypothetical protein 177 – 6.5Ac_My_14d_5.21.01 53439440 FD661759 G01 Patched sphingolipid transporter (Ncr1) 473 – 4.7Ac_My_14d_5.21.02 53439441 FD661760 G01 NHP2/L7aE family protein 284 – 6.8Ac_My_14d_5.21.15 53439442 FD661761 G01 Unknown function/hypothetical protein 84 – 5.1Ac_My_14d_5.27.02 53439447 FD661766 B10 Pyruvate decarboxylase 173 EC:4.1.1.1 32.6Ac_My_14d_5.29.01 53439448 FD661767 B06 Mannose-1-phosphate guanylyltransferase 352 EC:2.7.7.22 7.1Ac_My_14d_5.39.01 53439449 FD661768 B05 RgsA-like, regulator of G protein signalling domain protein 245 – 4.6Ac_My_14d_5.39.02 53439450 FD661769 B05 RNA polymerase II mediator complex component 175 – 4.1Ac_My_14d_Ctg01 53439452 FD661771 H10, C06 Protein phosphatase regulatory subunit, Gac1 812 EC:3.1.3.16 3.7Ac_My_14d_Ctg02 53439453 FD661772 H/C02 Unknown function/hypothetical protein 157 – 6.5Ac_My_14d_Ctg04 53439455 FD661774 A/B/F/G09 Pyruvate decarboxylase 663 EC:4.1.1.1 5.6Ac_My_14d_Ctg05 53439456 FD661775 E/D11 Unknown function/hypothetical protein 224 – 6.5Ac_My_14d_Ctg06 53439457 FD661776 E04/06/09 Unknown function/hypothetical protein 174 – 7.4Ac_My_14d_Ctg07 53439458 FD661777 H10, C08 Cellulose-binding (CBD), family II 393 EC:3.2.1 aAc_My_14d_Ctg08 53439459 FD661778 A10, G10/07 Membrane protein 191 – aAc_My_14d_Ctg11 53439462 FD661781 D06/07 Chitinase 361 EC:3.2.1.14 aAc_My_14d_Ctg12 53439463 FD661782 D06/07/03 Unknown function/hypothetical protein 292 – 17.4Ac_My_14d_Ctg13 53439464 FD661783 D02, E02 1,3-Beta-glucan synthase catalytic subunit 523 EC:2.4.1.34 6.2Ac_My_14d_Ctg14 53439465 FD661784 D02, E02 Cell wall biogenesis protein LRR/Mhp1 462 – 3.9Ac_My_14d_Ctg15 53439466 FD661785 D05/07 Unknown function/hypothetical protein 151 – 4.9Ac_My_14d_Ctg16 53439467 FD661786 G/A09 Myosin I 463 – 4.5Ac_My_14d_Ctg17 53439468 FD661787 B03 Unknown function/hypothetical protein 175 – 5.8
The up-regulation (Up-reg) was expressed as Li/Hi (Li = average net intensity of the band in the LP strain; Hi = average net intensity of the band in the HP strain). The letter ‘a’ meansthat the band was completely absent in the HP sample. For each EST the dbEST ID (http://www.ncbi.nlm.nih.gov/dbEST/), the GenBank accession number (http://www.ncbi.nlm.nih.gov/Genbank/index.html), the combination/s of primers used for the isolation, a tentative annotation, the size in base pairs, and the enzyme code (EC) are provided where available.
109A. Botton et al. / International Journal of Food Microbiology 127 (2008) 105–115
pyruvate metabolism. Concerning fatty acid metabolism, it couldconvert fatty acids into acetaldehyde, whereas in lysine degradation,it could be involved in the conversion of 4-trimetilammoniobutanalinto 4-trimetilammoniobutanoate. As far as the phenylalanine me-tabolism is concerned, the aldehyde dehydrogenase interconvertsphenyl-acetate into phenyl-acetaldehyde. The clone 5.27.02 and the
Contig 4 were both up-regulated, and putatively encode a pyruvatedecarboxylase involved in glycolysis/gluconeogenesis and pyruvateme-tabolism, by catalyzing the conversion of pyruvate into 2-hydroxyethyl-ThPP (thiamine disphosphate). The clone 3.42.01 was down-regulatedand putatively encodes a cytochrome P450 oxidoreductase showinga 61% of similarity with OrdA, a key enzyme of aflatoxin biosynthesis

Table 3Transcripts that were down-regulated in the ochratoxin-low-producing (LP) strain of Aspergillus carbonarius with respect to the high-producing (HP) one
EST dbEST GenBank Primers Tentative annotation Size EC Down-reg
Ac_My_14d_2.08.10 53439350 FD661669 H10 Zn(II)2Cys6 zinc finger domain, transcription factor 110 – 3.0Ac_My_14d_2.16.10 53439351 FD661670 H05 Unknown function/hypothetical protein 98 – 9.2Ac_My_14d_2.20.02 53439352 FD661671 H04 Unknown function/hypothetical protein 192 – 4.2Ac_My_14d_2.28.01 53439353 FD661672 F08 Molecular chaperone Mod-E/Hsp90 758 – aAc_My_14d_2.28.02 53439354 FD661673 F08 Phosphatidylinositol 4-kinase (PI4K) 561 EC:2.7.1.67 aAc_My_14d_2.28.04 53439355 FD661674 F08 CPA2 (Cation/Proton antiporter) transporter 305 – aAc_My_14d_2.28.05 53439356 FD661675 F08 FHA domain protein 275 – aAc_My_14d_2.28.06 53439357 FD661676 F08 Unknown function/hypothetical protein 269 – 3.5Ac_My_14d_2.28.07 53439358 FD661677 F08 Aldehyde dehydrogenase 165 EC:1.2.1.3, EC:1.2.1.5 3.7Ac_My_14d_2.28.08 53439359 FD661678 F08 30 kDa heat shock protein (HSP30) 150 – aAc_My_14d_2.28.10 53439360 FD661679 F08 Unknown function/hypothetical protein 115 – 5.0Ac_My_14d_2.28.11 53439361 FD661680 F08 Unknown function/hypothetical protein 74 – aAc_My_14d_2.28.12 53439362 FD661681 F08 Unknown function/hypothetical protein 48 – aAc_My_14d_3.06.01 53439367 FD661686 E08 Ubiquitin-like activating enzyme 333 – 3.2Ac_My_14d_3.06.02 53439368 FD661687 E08 Unknown function/hypothetical protein 186 – 9.8Ac_My_14d_3.06.08 53439369 FD661688 E08 Transcription initiation factor TFIId subunit TSM1 125 – 4.3Ac_My_14d_3.06.11 53439371 FD661690 E08 Unknown function/hypothetical protein 53 – 4.6Ac_My_14d_3.08.01 53439372 FD661691 E10 Phosphogluconate-2-dehydrogenase 869 EC:1.1.1.43 11.5Ac_My_14d_3.08.02 53439373 FD661692 E10 Arylsulfatase 543 EC:3.1.6.1 12.7Ac_My_14d_3.08.03 53439374 FD661693 E10 Translation elongation factor 1-alpha (EF-1 alpha) 1185 – 3.1Ac_My_14d_3.08.04 53439375 FD661694 E10 Cytochrome C-type haem-binding periplasmic protein 262 – 4.4Ac_My_14d_3.08.05 53439376 FD661695 E10 Calmodulin 203 – 10.7Ac_My_14d_3.08.06 53439377 FD661696 E10 5-oxo-L-prolinase 199 EC:3.5.2.9 9.7Ac_My_14d_3.08.08 53439378 FD661697 E10 Unknown function/hypothetical protein 217 – aAc_My_14d_3.08.09 53439379 FD661698 E10 Unknown function/hypothetical protein 127 – aAc_My_14d_3.24.09 53439384 FD661703 C11 Unknown function/hypothetical protein 108 – aAc_My_14d_3.28.09 53439390 FD661709 C08 Lysyl-tRNA synthetase 78 EC:6.1.1.6 3.7Ac_My_14d_3.42.01 53439393 FD661712 C04 Cytochrome P450 oxidoreductase 318 EC:1.6.2.4, EC:1.14.14.1 26.1Ac_My_14d_3.44.01 53439394 FD661713 C03 Unknown function/hypothetical protein 172 – 5.9Ac_My_14d_4.06.01 53439405 FD661724 A08 5-oxo-L-prolinase 550 EC:3.5.2.9 3.1Ac_My_14d_4.07.10 53439408 FD661727 A10 Pre-rRNA processing protein 159 – aAc_My_14d_4.07.12 53439409 FD661728 A10 Unknown function/hypothetical protein 155 – aAc_My_14d_4.10.01 53439410 FD661729 A06 Nuclear distribution protein nudE 632 – 5.3Ac_My_14d_4.12.01 53439411 FD661730 A07 Heat shock protein 70 (HSP70) 601 – 4.6Ac_My_14d_4.16.02 53439412 FD661731 A05 Unknown function/hypothetical protein 162 – 18.5Ac_My_14d_4.22.13 53439414 FD661733 A03 Unknown function/hypothetical protein 115 – 9.2Ac_My_14d_4.37.13 53439420 FD661739 D05 Unknown function/hypothetical protein 158 – 3.4Ac_My_14d_4.37.16 53439421 FD661740 D05 Unknown function/hypothetical protein 47 – 6.0Ac_My_14d_5.02.17 53439426 FD661745 G11 Diguanylate cyclase phosphodiesterase (GGDEF/EAL) 321 EC:4.6.1.2 4.4Ac_My_14d_5.02.18 53439427 FD661746 G11 Unknown function/hypothetical protein 121 – 11.3Ac_My_14d_5.06.03 53439429 FD661748 G08 DUF1749 protein 145 234.2Ac_My_14d_5.08.02 53439432 FD661751 G10 N-acetylglucosamine-6-phosphate deacetylase 300 EC:3.5.1.25 6.3Ac_My_14d_5.16.01 53439435 FD661754 G05 Unknown function/hypothetical protein 469 – 4.4Ac_My_14d_5.16.02 53439436 FD661755 G05 Protein phosphatase PP2A regulatory subunit B 350 – 4.0Ac_My_14d_5.16.15 53439437 FD661756 G05 Unknown function/hypothetical protein 137 – 6.3Ac_My_14d_5.16.17 53439438 FD661757 G05 Unknown function/hypothetical protein 91 – 30.3Ac_My_14d_5.16.19 53439439 FD661758 G05 Unknown function/hypothetical protein 93 – 3.3Ac_My_14d_5.24.01 53439443 FD661762 B11 Phosphogluconate-6-dehydrogenase (decarboxylating) 535 EC:1.1.1.44 10.2Ac_My_14d_5.24.19 53439444 FD661763 B11 Unknown function/hypothetical protein 77 – 3.6Ac_My_14d_5.24.22 53439445 FD661764 B11 Unknown function/hypothetical protein 75 – 3.2Ac_My_14d_5.24.25 53439446 FD661765 B11 Unknown function/hypothetical protein 67 – 4.2Ac_My_14d_5.44.15 53439451 FD661770 B03 Unknown function/hypothetical protein 65 – 32.5Ac_My_14d_Ctg03 53439454 FD661773 H04/06 Unknown function/hypothetical protein 204 – 3.7Ac_My_14d_Ctg09 53439460 FD661779 A07/02 Protocatechuate 4,5-dioxygenase 336 EC:1.13.11.8 5.4Ac_My_14d_Ctg10 53439461 FD661780 D11, F04 DJ-1/PfpI family protein 521 – 3.0
The down-regulation (Down-reg) was expressed as Hi/Li (Hi = average net intensity of the band in the HP strain; Li = average net intensity of the band in the LP strain). The letter ‘a’means that the band was completely absent in the LP sample. For each EST the dbEST ID (http://www.ncbi.nlm.nih.gov/dbEST/), the GenBank accession number (http://www.ncbi.nlm.nih.gov/Genbank/index.html), the combination/s of primers used for the isolation, a tentative annotation, the size, and the enzyme code (EC) are provided where available.
110 A. Botton et al. / International Journal of Food Microbiology 127 (2008) 105–115
(Ehrlich et al., 2004a). Moreover, it may be involved in fatty acid andtryptophan metabolism. The sequence 4.31.03, encoding a putativeasparaginyl-tRNA-synthetase, was up-regulated andmay be involved inalanine/aspartate metabolism and in aminoacyl-tRNA-biosynthesisby conjugating the specific tRNA to L-asparagine. The differentiallyexpressed EST 4.31.13 was up-regulated and putatively encodes theβ-subunit of a 3-methylcrotonyl-CoA carboxylase involved in fatty acid,pyruvate and propanoate metabolism. This enzyme is responsible forthe conversion of acetyl-CoA into malonyl-CoA. Contig 9 was down-regulated and encodes a putative protocatechuate-4,5-dioxygenase in-volved in the degradation of benzoate. The clones 3.08.01 and 3.08.06putatively encode a phosphogluconate-2-dehydrogenase and a 5-oxo-L-prolinase, respectively. Bothwere down-regulated andmay be involved
in glutathione metabolism, either in the conversion of NADP+ intoNADPH, or in the conversion of 5-oxoproline into L-glutamate. In thelatter reaction also the protein encoded by the down-regulated clone4.06.01 is involved. Finally, the sequence 3.03.01, that was up-regulatedin the LP strain, putatively encodes an arginyl-tRNA-synthetase con-jugating the specific tRNA to L-arginine.
3.4. Genes encoding proteins putatively involved in the regulation of OTAproduction
The regulation of OTA production may be exerted at different levels,both transcriptionally and post-transcriptionally. Among the differen-tially expressed clones, several transcription factorswere present among
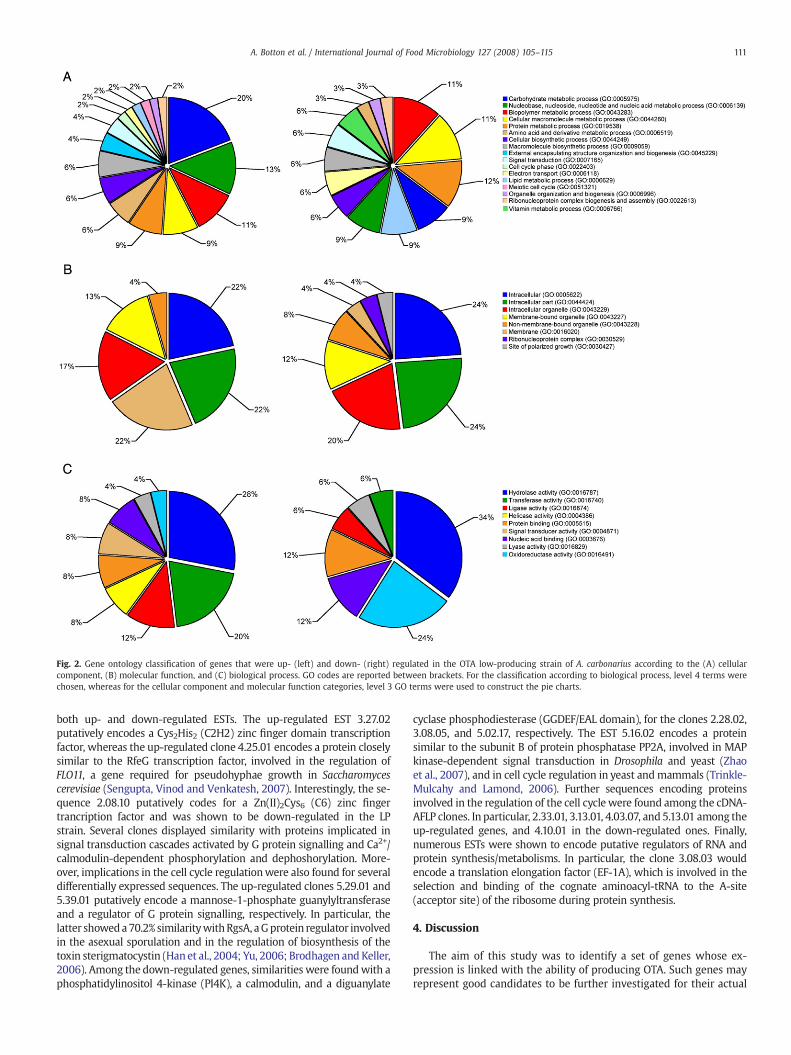
Fig. 2. Gene ontology classification of genes that were up- (left) and down- (right) regulated in the OTA low-producing strain of A. carbonarius according to the (A) cellularcomponent, (B) molecular function, and (C) biological process. GO codes are reported between brackets. For the classification according to biological process, level 4 terms werechosen, whereas for the cellular component and molecular function categories, level 3 GO terms were used to construct the pie charts.
111A. Botton et al. / International Journal of Food Microbiology 127 (2008) 105–115
both up- and down-regulated ESTs. The up-regulated EST 3.27.02putatively encodes a Cys2His2 (C2H2) zinc finger domain transcriptionfactor, whereas the up-regulated clone 4.25.01 encodes a protein closelysimilar to the RfeG transcription factor, involved in the regulation ofFLO11, a gene required for pseudohyphae growth in Saccharomycescerevisiae (Sengupta, Vinod and Venkatesh, 2007). Interestingly, the se-quence 2.08.10 putatively codes for a Zn(II)2Cys6 (C6) zinc fingertrancription factor and was shown to be down-regulated in the LPstrain. Several clones displayed similarity with proteins implicated insignal transduction cascades activated by G protein signalling and Ca2+/calmodulin-dependent phosphorylation and dephoshorylation. More-over, implications in the cell cycle regulationwere also found for severaldifferentially expressed sequences. The up-regulated clones 5.29.01 and5.39.01 putatively encode a mannose-1-phosphate guanylyltransferaseand a regulator of G protein signalling, respectively. In particular, thelatter showeda70.2% similaritywithRgsA, aGprotein regulator involvedin the asexual sporulation and in the regulation of biosynthesis of thetoxin sterigmatocystin (Han et al., 2004; Yu, 2006; Brodhagen and Keller,2006). Among the down-regulated genes, similarities were foundwith aphosphatidylinositol 4-kinase (PI4K), a calmodulin, and a diguanylate
cyclase phosphodiesterase (GGDEF/EAL domain), for the clones 2.28.02,3.08.05, and 5.02.17, respectively. The EST 5.16.02 encodes a proteinsimilar to the subunit B of protein phosphatase PP2A, involved in MAPkinase-dependent signal transduction in Drosophila and yeast (Zhaoet al., 2007), and in cell cycle regulation in yeast andmammals (Trinkle-Mulcahy and Lamond, 2006). Further sequences encoding proteinsinvolved in the regulation of the cell cycle were found among the cDNA-AFLP clones. In particular, 2.33.01, 3.13.01, 4.03.07, and 5.13.01 among theup-regulated genes, and 4.10.01 in the down-regulated ones. Finally,numerous ESTs were shown to encode putative regulators of RNA andprotein synthesis/metabolisms. In particular, the clone 3.08.03 wouldencode a translation elongation factor (EF-1A), which is involved in theselection and binding of the cognate aminoacyl-tRNA to the A-site(acceptor site) of the ribosome during protein synthesis.
4. Discussion
The aim of this study was to identify a set of genes whose ex-pression is linked with the ability of producing OTA. Such genes mayrepresent good candidates to be further investigated for their actual

Fig. 3. Bar chart showing the GO categories for which the up- and down-regulated sequences were significantly different (Pb0.05). The categories are ordered from the less (top) tothe most (down) significant.
Table 4Sequences encoding proteins putatively involved in the metabolism of ochratoxin Aprecursors
cDNA-AFLP clones Regulation KEGG map Pathway
Ac_My_14d_2.28.07, 5.27.02,Ctg4
Down, up,up
map00010 Glycolysis/gluconeogenesis
Ac_My_14d_4.31.13 Up map00061 Fatty acid biosynthesisAc_My_14d_2.28.07, 3.42.01 Down,
downmap00071 Fatty acid metabolism
Ac_My_14d_4.31.03 Up map00252 Alanine and aspartatemetabolism
Ac_My_14d_2.28.07 Down map00310 Lysine degradationAc_My_14d_2.28.07 Down map00360 Phenylalanine metabolismAc_My_14d_Ctg9 Down map00362 Benzoate degradation via
hydroxylationAc_My_14d_3.42.01 Down map00380 Tryptophan metabolismAc_My_14d_2.28.07 Down map00410 β-Alanine metabolismAc_My_14d_3.08.01, 3.08.06 Down,
downmap00480 Glutathione metabolism
Ac_My_14d_2.28.07, 4.31.13,5.27.02, Ctg4
Down, up,up, up
map00620 Pyruvate metabolism
Ac_My_14d_4.31.13 Up map00640 Propanoate metabolismAc_My_14d_3.03.01, 4.31.03 Up, up map00970 Aminoacyl-tRNA
biosynthesis
The codes of KEGG maps and the relative pathways are also reported.
112 A. Botton et al. / International Journal of Food Microbiology 127 (2008) 105–115
involvement in OTA biosynthesis and genetic regulation. The screen-ing was performed in two strains of A. carbonarius, antagonists for theability in producing OTA. This experimental plan, similar to thatadopted by OBrian et al. (2003), was chosen as an alternative to the“OTA permitting medium” approach (Färber and Geisen, 2004), toavoid the contamination due to genes whose expression is inducedby the differential growing conditions. The cDNA-AFLP techniquewas successfully adopted in Aspergillus for transcriptome profiling,allowing the identification of 383 differentially expressed amplicons,119 of which were sequenced and partially characterized by meansof bioinformatics analyses. A similar approach was adopted also byMühlencoert (2003) in A. ochraceus.
The sequences were classified according to the gene ontologycriteria, allowing to identify the cellular component (CC) in which thegene products exert their biological role, their molecular function(MF), and the biological processes (BP) in which they are putativelyinvolved. The Fisher's exact test was performed by using the GOSSIPtool integrated in the Blast2GO software, allowing a comparison of theannotation sets of up- and down-regulated genes by pointing outthe GO categories for which the two groups significantly differed. Theresults of this analysis should be discussed together with theconsiderations regarding the genes encoding enzymes putativelyinvolved in the biosynthesis and those encoding regulative elements.Therefore, a possible model for OTA biosynthesis regulation may behypothesized, as displayed in Fig. 4, according to the up- and down-regulation patterns pointed out by means of cDNA-AFLP differentialdisplay. According to this model, OTA biosynthesis may depend bothon the availability of precursors and on the developmental stage, aspointed out for several other mycotoxins such as fumonisins (Shimand Woloshuk, 2001), aflatoxins, and sterigmatocystin (Wilkinsonet al., 2004). Several proteins among those putatively encoded by thedifferentially expressed clones herein isolated may regulate the avail-ability of OTA precursors. The aldehyde dehydrogenase encoded bythe clone 2.28.07 may deprive OTA biosynthesis both of the acetate
and, more indirectly, of the phenylalanine necessary for synthesizingOTA, since it may be active in glycolysis, and in fatty acid, pyruvate andphenylalanine metabolism. Interestingly, the EST 3.42.01 was shownto encode a cytochrome P450 oxidoreductase 61% similar to OrdA,which is required for aflatoxin biosynthesis (Ehrlich et al., 2004a). Thisenzyme may be involved both indirectly and directly in OTA pro-duction, since it may synthesize precursors from the fatty acids meta-bolism and be responsible for catalyzing limiting reactions in the OTAbiosynthetic pathway as well. In particular, an oxidation step is re-quired to convert mellein into OTβ, and no candidate enzyme has

Fig. 4. Putative model for OTA biosynthesis regulation. Dotted lines represent possiblelinks between processes, whereas arrowheads and blocked lines indicate positive andnegative effects, respectively. Transcription factors are reported in ellipses. Red andgreen indicate up- and down-regulated elements with matches among the differen-tially expressed cDNA-AFLP clones (see Tables 2 and 3). Cyan boxes represent the mainfactors supposed to directly or indirectly regulate OTA biosynthesis.
113A. Botton et al. / International Journal of Food Microbiology 127 (2008) 105–115
been yet identified for this limiting step. Both the latter genes weredown-regulated in the OTA-low-producing strain (LP), as for themajority of the differentially expressed sequences putatively involvedin generation of OTA precursors. Another interesting down-regulatedclone was 4.31.13, coding for a carboxylase subunit involved in theinterconversion of acetyl-CoA into malonyl-CoA, both potential pre-cursors of OTA. As a general remark, the ‘oxidoreductase activity’ wasone of the GO categories most significantly differing between theannotation sets of up- and down-regulated genes, along with ‘cyto-plasm’, ‘carbohydrate metabolic process’, and ‘generation of precursormetabolites and energy’. All this GO terms may concern the avail-ability of precursors for the OTA pathway.
Further GO annotations were shown to significantly differ in thetwo groups of sequences, namely ‘membrane’, ‘transferase activity’,‘signal transducer activity’, and ‘cell cycle’. Several putative transcrip-tion factors were identified and their orthologs shown to be involvedin the regulation of mycotoxin biosynthesis and sexual/asexual spo-rulation. Since there are common signalling pathways connectingmycotoxin biosynthesis and sporulation (Brodhagen and Keller, 2006),a possible role may be also hypothesized for the clones 3.27.02,4.25.01, and 2.08.10. The former, encoding a C2H2 zinc finger trans-cription factor, was up-regulated in the LP strain, and its orthologsfrequently found to trigger cAMP/PKA signal cascades involved in theregulation of sexual/asexual fungi development (Görner et al., 1998;Hurtado and Rachubinski, 1999; Coppin, 2002). The second was up-regulated as well, and encodes a protein closely similar to the RfeGtranscription factor, involved in the regulation of FLO11, a gene re-quired for pseudohyphae formation in S. cerevisiae (Sengupta et al.,2007). Finally, the latter, encoding a putative C6 zinc finger trans-cription factor, was shown to be down-regulated in the LP strain. InA. nidulans as in other Aspergillus species, this family of proteins hasbeen claimed to be involved in the mycotoxin biosynthesis (Fer-nandes, Keller and Adams, 1998: Brodhagen and Keller, 2006). Thus,the C2H2 and RfeG transcription factors may inhibit OTA biosynthesis,
since they are both up-regulated in the low-producing strain, whereasthe C6 zinc finger may act as positive regulator (Fig. 4). A possibleregulation may also be exerted throughout the G protein signalling,since many sequences matched with elements involved in suchtransduction pathways and in the Ca2+/calmodulin-dependent phos-phorylation and dephoshorylation cascades. Three co-regulated geneswere identified as encoding a Gac1-like phosphatase, a Ser/Thr proteinkinase, and a RgsA-like transcription factor. The latter showed a highdegree of similarity (70.2%) with RgsA (regulator of G protein sig-nalling A) of A. nidulans, involved in the biosynthesis of sterigmato-cystin and in asexual sporulation (Han et al., 2004). On one hand,these elements may negatively regulate OTA biosynthesis, since theywere all encoded by genes that were up-regulated in the strainproducing low levels of the toxin. On the other hand, further genesmay activate the biosynthetic pathway, because their expressionpatterns were positively correlated with the production of OTA. TheESTs 3.08.05, 2.28.02, 5.02.17, and 5.16.02 were all down-regulatedin LP, and putatively encode a calmodulin, a PI4 kinase, a proteininvolved in GTP metabolism, and a PP2A phosphatase (Fig. 4). The se-veral genes involved in the regulation of cell cycle, mostly up-regulated,may be linked to sexual/asexual sporulation involving meiosis and/ormitosis. During these processes, the high number of genes encodingproteinswith DNA repairing functionmay guarantee the integrity of theDNA. Nevertheless, it is worth noting that the clone 3.08.03, encoding atranslation elongation factor (EF-1A), showed a regulation patternsimilar to a gene identified by Färber and Geisen (2004) in a screeningfor OTA-related genes in P. nordicum. A positive correlation was foundbetween its expression pattern and OTA production.
Concluding, the transcriptional profiling carried out in the presentresearch allowed the identification of 119 differentially expressed genesputatively involved in the biosynthesis of OTA and its regulation. A draftmodel was proposed according to previous findings concerning theirorthologs. Moreover, a likely connection was pointed out between OTAbiosynthesis and sexual/asexual sporulation, along with common sig-nalling pathways. Indeed, the involvement of G protein and Ca2+/calmo-dulin signalling was proposed, based upon the high number of clonesencoding putative elements of such transduction pathways. Importanttranscription factors were also identified, such as the Cys2His2 and theZn(II)2Cys6 zinc fingers, possibly acting antagonistically. Concluding, thepresent study gives an important contribution to the understandingof OTA biosynthesis, even though specific studies should be addressedto the functional characterization of the genes herein isolated, eitherby silencing or knock-out approaches. Such experiments are currently inprogress and may have important implications in the full comprehen-sion of the biosynthesis of OTA, a toxin that may become a seriousproblem not only for the European Community, but also for theworldwide wine industry.
Acknowledgements
This research was supported by the Azione Biotech 2 of ISIB-CNR,titled “Biotechnology to improve crop quality, safety and sustain-ability”, funded by the Veneto Region.
References
Albertini, E., Marconi, G., Barcaccia, G., Raggi, L., Falcinelli, M., 2004. Isolation of candidategenes for apomixis in Poa pratensis L. Plant Molecular Biology 56, 879–894.
Aparicio, G., Gotz, S., Conesa, A., Segrelles, D., Blanquer, I., Garcia, J.M., Hernandez, V., Robles,M., Talon, M., 2006. Blast2GO goes grid: developing a grid-enabled prototype for func-tional genomics analysis. Studies in Health Technology and Informatics 120, 194–204.
Ashburner, M., Ball, C.A., Blake, J.A., Botstein, D., Butler, H., Cherry, J.M., Davis, A.P., Dolinski,K., Dwight, S.S., Eppig, J.T., Harris, M.A., Hill, D.P., Issel-Tarver, L., Kasarskis, A., Lewis, S.,Matese, J.C., Richardson, J.E., Ringwald, M., Rubin, G.M., Sherlock, G., 2000. Geneontology: tool for the unification of biology. The Gene Ontology Consortium. NatureGenetics 25, 25–29.
Atoui, A., Mathieu, F., Lebrihi, A., 2007a. Targeting a polyketide synthase gene for As-pergillus carbonarius quantification and ochratoxin A assessment in grapes usingreal-time PCR. International Journal of Food Microbiology 115, 313–318.

114 A. Botton et al. / International Journal of Food Microbiology 127 (2008) 105–115
Atoui, A., Mitchell, D., Mathieu, F., Magan, N., Lebrihi, A., 2007b. Partitioning of ochratoxinA in mycelium and conidia of Aspergillus carbonarius and the impact on toxincontamination of grapes and wine. Journal of Applied Microbiology 103, 961–968.
Bacaloni, A., Cavaliere, C., Faberi, A., Pastorini, E., Samperi, R., Laganà, A., 2005. Auto-mated on-line solid-phase extraction-liquid chromatography–electrospray tandemmass spectrometry method for the determination of ochratoxin A inwine and beer.Journal of Agricultural and Food Chemistry 53, 5518–5525.
Bachem, C.W., van der Hoeven, R.S., de Bruijn, S.M., Vreugdenhil, D., Zabeau, M., Visser,R.G., 1996. Visualization of differential gene expression using a novel method ofRNA fingerprinting based on AFLP: analysis of gene expression during potato tuberdevelopment. The Plant Journal 9, 745–753.
Barcaccia, G., Albertini, E., Tavoletti, S., Falcinelli, M., Veronesi, F., 1999. AFLP finger-printing in Medicago spp.: its development and application in linkage mapping.Plant Breeding 118, 335–340.
Barcaccia, G., Varotto, S., Albertini, E., Porceddu, A., Meneghetti, S., Parrini, P., Lucchin,M., 2001. Analysis of gene expression during flowering in apomeiotic mutants ofMedicago spp.: cloning ESTs and candidate genes for apomeiosis. Sexual PlantReproduction 14, 233–238.
Basílico, M.Z., Basílico, J.C., 1999. Inhibitory effects of some spice essential oils on Asper-gillus ochraceus NRRL 3174 growth and ochratoxin A production. Letters in AppliedMicrobiology 29, 238–241.
Battilani, P., Giorni, P., Pietri, A., 2003. Epidemiology of toxin-producing fungi andochratoxinAoccurrence in grape. European Journal of Plant Pathology 109, 715–722.
Bau, M., Castellá, G., Bragulat, M.R., Cabañes, F.J., 2005. DNA-based characterization ofochratoxin-A-producing and non-producing Aspergillus carbonarius strains fromgrapes. Research in Microbiology 156, 375–381.
Bayman, P., Baker, J.L., 2006. Ochratoxins: a global perspective. Mycopathologia 162,215–223.
Belli, N., Marín, S., Coronas, I., Sanchis, V., Ramos, A.J., 2007. Skin damage, high tem-perature and relative humidity as detrimental factors for Aspergillus carbonariusinfection and ochratoxin A production in grapes. Food Control 18, 1343–1349.
Belli, N., Marín, S., Duaigues, A., Ramos, A.J., Sanchis, V., 2004. Ochratoxin A in wines,musts and grape juices from Spain. Journal of the Science of Food and Agriculture84, 591–594.
Blesa, J., Soriano, J.M., Molto, J.C., Manes, J., 2004. Concentration of ochratoxin A in winesfrom supermarkets and stores of Valencian Community (Spain). Journal of Chro-matography A 1054, 397–401.
Bogs, C., Battilani, P., Geisen, R., 2006. Development of a molecular detection and dif-ferentiation system for ochratoxin A producing Penicillium species and its applicationto analyse the occurrence of Penicilliumnordicum in curedmeats. International Journalof Food Microbiology 107, 39–47.
Borneman, A.R., Hynes, M.J., Andrianopoulos, A., 2001. An STE12 homolog from theasexual, dimorphic fungus Penicillium marneffei complements the defect in sexualdevelopment of an Aspergillus nidulans steA mutant. Genetics 157, 1003–1014.
Botton, A., Barcaccia, G., Cappellozza, S., Da Tos, R., Bonghi, C., Ramina, A., 2005. DNAfingerprinting sheds light on the origin of introduced mulberry (Morus spp.)accessions in Italy. Genetic Resources and Crop Evolution 52, 181–192.
Botton, A., Galla, G., Conesa, A., Bachem, C., Ramina, A., Barcaccia, G., 2008. Large-scaleGene Ontology analysis of plant transcriptome-derived sequences retrieved byAFLP technology. BMC Genomics 9, 347.
Brera, C., Soriano, J.M., Debegnach, F., Miraglia, M., 2005. Exposure assessment toochratoxin A from the consumption of Italian and Hungarian wines. MicrochemicalJournal 79, 109–113.
Brodhagen, M., Keller, N.P., 2006. Signalling pathways connectingmycotoxin productionand sporulation. Molecular Plant Pathology 7, 285–301.
Cnudde, F., Hedatale, V., de Jong, H., Pierson, E.S., Rainey, D.Y., Zabeau, M., Weterings, K.,Gerats, T., Peters, J.L., 2006. Changes in gene expression during male meiosis inPetunia hybrida. Chromosome Research 14, 919–932.
Conesa, A., Goetz, S., García-Gómez, J.M., Terol, J., Talon, M., Robles, M., 2005. Blast2GO:a universal tool for annotation, visualization and analysis in functional genomicsresearch. Bioinformatics 21, 3674–3676.
Coppin, E., 2002. The FLE1 gene encoding a C2H2 zinc finger protein co-ordinates maleand female sexual differentiation in Podospora anserina. Molecular Microbiology43, 1255–1268.
Dao, H.P., Mathieu, F., Lebrihi, A., 2005. Two primer pairs to detect OTA producers byPCR method. International Journal of Food Microbiologo 104, 61–67.
Donson, J., Fang, Y., Espiritu-Santo, G., Xing, W., Salazar, A., Miyamoto, S., Armendarez,V., Volkmuth, W., 2002. Comprehensive gene expression analysis by transcriptprofiling. Plant Molecular Biology 48, 75–97.
Durrant, W.E., Rowland, O., Piedras, P., Hammond-Kosack, K.E., Jones, J.D., 2000. cDNA-AFLP reveals a striking overlap in race-specific resistance andwound response geneexpression profiles. The Plant Cell 12, 963–977.
Ehrlich, K.C., Cotty, P.J., 2004b. An isolate of Aspergillus flavus used to reduce aflatoxincontamination in cottonseed has a defective polyketide synthase gene. AppliedMicrobiologo and Biotechnology 65, 473–478.
Ehrlich, K.C., Chang, P.K., Yu, J.J., Cotty, P.J., 2004a. Aflatoxin biosynthesis cluster genecypA is required for G aflatoxin formation. Applied and Environmental Micro-biology 70, 6518–6524.
Ehrlich, K.C., Montalbano, B.G., Cary, J.W., 1999. Binding of the C6-zinc cluster protein,AFLR, to the promoters of aflatoxin pathway biosynthesis genes in Aspergillusparasiticus. Gene 230, 249–257.
Färber, P., Geisen, R., 2004. Analysis of differentially-expressed ochratoxinAbiosynthesisgenes of Penicillium nordicum. European Journal of Plant Pathology 110, 661–669.
Fernandes, M., Keller, N.P., Adams, T.H., 1998. Sequence-specific binding by Aspergillusnidulans AflR, a C6 zinc cluster protein regulating mycotoxin biosynthesis.Molecular Microbiology 28, 1355–1365.
Geisen, R., Mayer, Z., Karolewiez, A., Färber, P., 2004. Development of a real time PCRsystem for detection of Penicillium nordicum and for monitoring ochratoxin Aproduction in foods by targeting the ochratoxin polyketide synthase gene.Systematic & Applied Microbiology 27, 501–507.
Görner, W., Durchschlag, E., Martinez-Pastor, M.T., Estruch, F., Ammerer, G., Hamilton, B.,Ruis, H., Schüller, C.,1998.Nuclear localization of theC2H2zincfingerproteinMsn2p isregulated by stress and protein kinase A activity. Genes & Development 12, 586–597.
Han, K.H., Seo, J.A., Yu, J.H., 2004. Regulators of G-protein signalling in Aspergillusnidulans: RgsA downregulates stress response and stimulates asexual sporulationthrough attenuation of GanB (Gα) signalling. Molecular Microbiology 53, 529–540.
Harris, J.P., Mantle, P.G., 2001. Biosynthesis of ochratoxins by Aspergillus ochraceus.Phytochemistry 58, 709–716.
Hernandez, M.J., Garcia-Moreno,M.V., Duran, E., Guillen, D., Barroso, C.G., 2006. Validationof two analytical methods for the determination of ochratoxin A by reversed-phasedhigh-performance liquid chromatography coupled to fluorescence detection inmustsand sweet wines from Andalusia. Analytica Chimica Acta 566, 117–121.
Hurtado, C.A., Rachubinski, R.A., 1999. MHY1 encodes a C2H2-type zinc finger proteinthat promotes dimorphic transition in the yeast Yarrowia lipolytica. Journal ofBacteriology 181, 3051–3057.
Jayashree, T., Rao, J.P., Subramanyam, C., 2000. Regulation of aflatoxinproduction byCa2+/calmodulin-dependent protein phosphorylation and dephosphorylation. FEMSMicrobiology Letters 183, 215–219.
Jones, C.S., Davies, H.V., Taylor, M.A., 2000. Profiling of changes in gene expressionduring raspberry (Rubus idaeus) fruit ripening by application of RNA fingerprintingtechniques. Planta 211, 708–714.
Lopez de Cerain, A., González-Peñas, E., Jiménez, A.M., Bello, J., 2002. Contribution to thestudy of ochratoxin A in Spanish wines. Food Additives and Contaminants 19,1058–1064.
Martinez-Culebras, P.V., Ramon, D., 2007. An ITS-RFLP method to identify black Asper-gillus isolates responsible for OTA contamination in grapes and wine. InternationalJournal of Food Microbiology 113, 147–153.
Mateo, R., Medina, A., Mateo, E.M., Mateo, F., Jimenez, M., 2007. An overview ofochratoxin A in beer and wine. International Journal of Food Microbiology 119,79–83.
Mateo, R., Medina, A., Mateo, F., Valle-Algarra, F.M., Gimeno-Adelantado, J.V., Jiménez,M., 2006. Optimization of themethodology for the determination of ochratoxin A inwine and study of its occurrence in wines consumed in Spain. Proceedings of the29th World Congress of the Vine and Wine. CD Edition, Logroño (Spain).
Mitchell, D., Parra, R., Aldred, D., Magan, N., 2004. Water and temperature relations ofgrowth and ochratoxin A production by Aspergillus carbonarius strains from grapesin Europe and Israel. Journal of Applied Microbiology 97, 439–445.
Müelencoert, E., 2003. Ochratoxin A Production by Aspergillus ochraceus. Doctoral thesis(http://tumb1.biblio.tu-muenchen.de/publ/diss/ww/2004/muehlencoert.pdf).
Myhre, S., Tveit, H., Mollestad, T., Laegreid, A., 2006. Additional gene ontology structurefor improved biological reasoning. Bioinformatics 22, 2020–2027.
Niessen, L., Schmidt, H., Muhlencoert, E., Färber, P., Karolewiez, A., Geisen, R., 2005.Advances in the molecular diagnosis of ochratoxin A-producing fungi. FoodAdditives and Contaminants 22, 324–334.
O'Callaghan, J., Caddick, M.X., Dobson, A.D.W., 2003. A polyketide synthase generequired for ochratoxin A biosynthesis in Aspergillus ochraceus. Microbiology 149,3485–3491.
OBrian, G.R., Fakhoury, A.M., Payne, G.A., 2003. Identification of genes differentiallyexpressed during aflatoxin biosynthesis in Aspergillus flavus and Aspergillusparasiticus. Fungal Genetics and Biology 39, 118–127.
Pardo, E., Sanchis, V., Ramos, A.J., Marín, S., 2006. Non-specificity of nutritional substratefor ochratoxin A production by isolates of Aspergillus ochraceus. Food Microbiology23, 351–358.
Pateraki, M., Dekanea, A., Mitchell, D., Lydakis, D., Magan, N., 2007. Influence of sulphurdioxide, controlled atmospheres and water availability on in vitro germination,growth and ochratoxin A production by strains of Aspergillus carbonarius isolatedfrom grapes. Postharvest Biology and Technology 44, 141–149.
Patiño, B., González-Salgado, A., González-Jaén, M.T., Vazquez, C., 2005. PCR detectionassays for the ochratoxin-producing Aspergillus carbonarius and Aspergillusochraceus species. International Journal of Food Microbiologo 104, 207–214.
Perrone, G., Mule, G., Susca, A., Battilani, P., Pietri, A., Logrieco, A., 2006. Ochratoxin Aproduction and amplified fragment length polymorphism analysis of Aspergilluscarbonarius, Aspergillus tubingensis, and Aspergillus niger strains isolated fromgrapes in Italy. Applied and Environmental Microbiology 72, 680–685.
Pietri, A., Bertuzzi, T., Pallaroni, L., Piva, G., 2001. Occurrence of ochratoxin A in Italianwines. Food Additives and Contaminants 18, 647–654.
Ponsone, M.L., Combina, M., Dalcero, A., Chulze, S., 2007. Ochratoxin A and ochra-toxigenic Aspergillus species in Argentinean wine grapes cultivated under organicand non-organic systems. International Journal of Food Microbiology 114, 131–135.
Rao, J.P., Subramanyam, C., 2000. Calmodulin mediated activation of acetyl-CoA car-boxylase during aflatoxin production by Aspergillus parasiticus. Letters in AppliedMicrobiology 30, 277–281.
Ringot, D., Chango, A., Schneider, Y.J., Larondelle, Y., 2006. Toxicokinetics and toxicody-namics of ochratoxin A, an update. Chemico-Biological Interactions 159, 18–46.
Sambrook, J., Russell, W.D., 2001. Molecular Cloning: a Laboratory Manual. CSHL Press,Woodbury NY, USA.
Schmidt, H., Taniwaki, M.H., Vogel, R.F., Niessen, L., 2004. Utilization of AFLP markers forPCR-based identification of Aspergillus carbonarius and indication of its presence ingreen coffee samples. Journal of Applied Microbiology 97, 899–909.
Sengupta, N., Vinod, P.K., Venkatesh, K.V., 2007. Crosstalk between cAMP-PKA and MAPkinase pathways is a key regulatory design necessary to regulate FLO11 expression.Biophysical Chemistry 125, 59–71.

115A. Botton et al. / International Journal of Food Microbiology 127 (2008) 105–115
Shim, W.B., Woloshuk, C.P., 2001. Regulation of fumonisin B1 biosynthesis andconidiation in Fusarium verticillioides by a cyclin-like (C-type) gene, FCC1. Appliedand Environmental Microbiology 67, 1607–1612.
Trinkle-Mulcahy, L., Lamond, A.I., 2006. Mitotic phosphatases: no longer silent partners.Current Opinion in Cell Biology 18, 623–631.
Tsitsigiannis, D.I., Keller, N.P., 2006. Oxylipins act as determinants of natural productbiosynthesis and seed colonization in Aspergillus nidulans. Molecular Microbiology59, 882–892.
Tuite, J., 1969. Plant Pathological Methods: Fungi and Bacteria. Burgess PublishingCompany, Minneapolis, MN, USA.
Valero, A., Marín, S., Ramos, A.J., Sanchis, V., 2007a. Effect of preharvest fungicides andinteracting fungi on Aspergillus carbonarius growth and ochratoxin A synthesis indehydrating grapes. Letters in Applied Microbiology 45, 194–199.
Valero, A., Olivan, S., Marin, S., Sanchis, V., Ramos, A.J., 2007b. Effect of intra andinterspecific interaction on OTA production by A. section Nigri in grapes duringdehydration. Food Microbiology 24, 254–259.
Varga, J., Péteri, Z., Tábori, K., Téren, J., Vágvölgyi, C., 2005. Degradation of ochratoxin A andother mycotoxins by Rhizopus isolates. International Journal of Food Microbiology 99,321–328.
Varga, J., Rigó, K., Kocsubé, S., Farkas, B., Pal, K., 2003. Diversity of polyketide synthasegene sequences in Aspergillus species. Research in Microbiology 154, 593–600.
Visconti, A., Pascale, M., Centonze, G., 2000. Determination of ochratoxin A in domesticand imported beers in Italy by immunoaffinity clean-up and liquid chromato-graphy. Journal of Chromatography A 888, 321–326.
Vos, P., Hogers, R., Bleeker, M., Reijans, M., van de Lee, T., Hornes, M., Frijters, A., Pot, J.,Peleman, J., Kuiper, M., 1995. AFLP: a new technique for DNA fingerprinting. NucleicAcids Research 23, 4407–4414.
Wilkinson, H.H., Ramaswamy, A., Sim, S.C., Keller, N.P., 2004. Increased conidiation as-sociated with progression along the sterigmatocystin biosynthetic pathway. Mycolo-gia 96, 1190–1198.
Yu, J.H., 2006. Heterotrimeric G protein signaling and RGSs in Aspergillus nidulans.Journal of Microbiology 44, 145–154.
Zanzotto, A., Burruano, S., Marciano, P., 2006. Digestion of DNA regions to discriminateochratoxigenic and non-ochratoxigenic strains in the Aspergillus niger aggregate.International Journal of Food Microbiology 110, 155–159.
Zdobnov, E.M., Apweiler, R., 2001. InterProScan — an integration platform for thesignature-recognition methods in InterPro. Bioinformatics 17, 847–848.
Zhao, B., Sun, L., Haas, M., Denenberg, A.G., Wong, H.R., Shanley, T.P., 2007. PP2A regu-lates upstream members of the C-jun N-terminal kinase mitogen-activated proteinkinase signaling pathway. Shock 29, 181–189.


















