a case study in a borderline area (Liguria, NW Italy)
16
The influence of climate on the distribution of lichens: a case study in a borderline area (Liguria, NW Italy) Paolo Giordani Guido Incerti Received: 7 June 2006 / Accepted: 24 May 2007 / Published online: 22 June 2007 Ó Springer Science+Business Media B.V. 2007 Abstract The purposes of this article are to quantify the relationship between epiphytic lichen distribution and macroclimatic variables in the study area and to provide a case study for evaluating the predictive role of epiphytic lichens as bioclimatic indicators. The study was carried out in the Liguria region (NW- Italy), a small (5432 km 2 ) borderline area, where phytoclimatic features range from the dry Mediterra- nean to the Alpine in a few kilometers. Epiphytic lichen diversity was sampled using a standardized protocol [Asta et al (2002) In: Nimis et al (eds) Monitoring with lichens: monitoring lichens. Kluwer, Dordrecht]. Abundance of the species in the sampling sites was related to macroclimatic parameters (yearly average temperature and rainfall) and non-parametric multivariate models were calculated to find significa- tive relationships among predictive and response variables. A total of 59 species showed highly significant relation with macroclimatic parameters. Four groups were selected, by means of a cluster analysis, related to four climatic niches (warm-humid, cold-humid, mesothermic-humid, warm-dry). Distri- butional pattern of the groups in the survey area showed a good correspondence with the bioclimatic units of Liguria region described by Nimis [(2003) Checklist of the Lichens of Italy 3.0. University of Trieste, Dept of Biology. http:// www.dbiodbs.univ.trieste.it. Cited 1 Jun 2006]. A significant subset of epiphytic lichen species in the study area have been proved to be efficient bioclimatic indicator and it is supposed to give good results to monitor climatic changes, in a long-term perspective. Keywords Distributional pattern Á Epiphytic lichen Á NPMR Á Bioclimatic indicators Á Poikilohydric organisms Introduction Lichens physiology is strongly related to both micro- and macro-climatic factors, since it is well known that relative air humidity and rainfall are the main sources of water supply for these organisms (e.g., Barkman 1958). The relationship between atmo- spheric water supply and lichen physiology has been investigated by several authors, showing that thallus hydric saturation is an important parameter in regu- lating metabolic processes such as gas exchanges, nitrogen fixation and photosynthesis (Rundel 1988; Nash 1996). In last years several studies have pointed out the role of climatic factors in the relationship between P. Giordani (&) Dipartimento per lo Studio del Territorio e delle sue Risorse, Laboratorio di Biologia e Diversita ` dei Vegetali, Corso Dogali 1M, 16136 Genova, Italy e-mail: [email protected] G. Incerti Dipartimento di Biologia, Universita ` di Trieste, Via L. Giorgieri 10, 34127 Trieste, Italy 123 Plant Ecol (2008) 195:257–272 DOI 10.1007/s11258-007-9324-7
Transcript of a case study in a borderline area (Liguria, NW Italy)

The influence of climate on the distribution of lichens:a case study in a borderline area (Liguria, NW Italy)
Paolo Giordani Æ Guido Incerti
Received: 7 June 2006 / Accepted: 24 May 2007 / Published online: 22 June 2007
� Springer Science+Business Media B.V. 2007
Abstract The purposes of this article are to quantify
the relationship between epiphytic lichen distribution
and macroclimatic variables in the study area and to
provide a case study for evaluating the predictive role
of epiphytic lichens as bioclimatic indicators. The
study was carried out in the Liguria region (NW-
Italy), a small (5432 km2) borderline area, where
phytoclimatic features range from the dry Mediterra-
nean to the Alpine in a few kilometers. Epiphytic
lichen diversity was sampled using a standardized
protocol [Asta et al (2002) In: Nimis et al (eds)
Monitoring with lichens: monitoring lichens. Kluwer,
Dordrecht]. Abundance of the species in the sampling
sites was related to macroclimatic parameters (yearly
average temperature and rainfall) and non-parametric
multivariate models were calculated to find significa-
tive relationships among predictive and response
variables. A total of 59 species showed highly
significant relation with macroclimatic parameters.
Four groups were selected, by means of a cluster
analysis, related to four climatic niches (warm-humid,
cold-humid, mesothermic-humid, warm-dry). Distri-
butional pattern of the groups in the survey area
showed a good correspondence with the bioclimatic
units of Liguria region described by Nimis [(2003)
Checklist of the Lichens of Italy 3.0. University of
T r i e s t e , D e p t o f B i o l o g y . h t t p : / /
www.dbiodbs.univ.trieste.it. Cited 1 Jun 2006]. A
significant subset of epiphytic lichen species in the
study area have been proved to be efficient bioclimatic
indicator and it is supposed to give good results to
monitor climatic changes, in a long-term perspective.
Keywords Distributional pattern � Epiphytic
lichen � NPMR � Bioclimatic indicators �Poikilohydric organisms
Introduction
Lichens physiology is strongly related to both micro-
and macro-climatic factors, since it is well known
that relative air humidity and rainfall are the main
sources of water supply for these organisms (e.g.,
Barkman 1958). The relationship between atmo-
spheric water supply and lichen physiology has been
investigated by several authors, showing that thallus
hydric saturation is an important parameter in regu-
lating metabolic processes such as gas exchanges,
nitrogen fixation and photosynthesis (Rundel 1988;
Nash 1996).
In last years several studies have pointed out the
role of climatic factors in the relationship between
P. Giordani (&)
Dipartimento per lo Studio del Territorio e delle sue
Risorse, Laboratorio di Biologia e Diversita dei Vegetali,
Corso Dogali 1M, 16136 Genova, Italy
e-mail: [email protected]
G. Incerti
Dipartimento di Biologia, Universita di Trieste, Via L.
Giorgieri 10, 34127 Trieste, Italy
123
Plant Ecol (2008) 195:257–272
DOI 10.1007/s11258-007-9324-7

lichen physiology and distribution (Kappen 1988;
McCune et al. 1997; Van Herk et al. 2002; Brunialti
and Giordani 2003). According to Nimis and Losi
(1984), lichen distribution could be an useful tool to
assess bioclimatic features of a territory, since these
symbiotic organisms, with respect to higher plants
and animals, are more selectively distributed along
ecological gradients where the local climate plays a
major role.
A number of epiphytic lichen species have been
proved to be distributed along ecological ranges
defined by the variation of main climatic factors
(Sancho et al. 1997; Fos et al. 1999; Shirazi et al.
1996). To better define bioclimatic variability at
regional scale, the relationship between species
distribution and climatic variability should be eval-
uated. It deeply depends from the scale of observa-
tion: at a strictly local level, lichens distribution is
scarcely affected by the macroclimate variability, and
the ecological response is rather related to microcli-
matic factors (for a case study see Coxson and Coyle
2003), so that narrow-ranging species should not be
considered as bioclimatic indicators. Also broad-
ranging species appear inadequate, since they are too
little sensible to the minimal mesoclimatic differ-
ences observable at regional level. Some species are
distributed according to extremely specific mesocli-
matic conditions, and they are susceptible to give
detailed information (Brunialti and Giordani 2003).
Unfortunately most of them are extremely rare and,
since their spot-like distribution, it is rather compli-
cated to use them to mark borders of bioclimatic
units.
According to Will-Wolf et al. (2002), the inter-
pretation of lichen distribution can be improved by
considering groups of species as bioclimatic indica-
tors, once the ecological features of all the species in
a group are homogeneous and well-defined. Upon
these basis, one could mark bioclimatic borders by
means of an a priori defined group of species with
well-known climatic requirements, such as lichens
with oceanic affinities, as described in Degelius
(1935), Rose (1976) and McCune (1984). Neverthe-
less this approach highly depends on the definition of
species affinities, which are often reported at global
scale, whilst the needed ecological information
should be referred to a regional level.
Furthermore, climatic affinities of species are often
expressed in a qualitative, or at least in a semi-
quantitative way, when some ecological scores along
climatic gradients are attributed to the species and
expressed on ordinal scales (Wirth 1991; Nimis
2003). This approach has the advantage of giving a
synthetic ecological characterization of almost every
taxa in national floras, in spite of a main shortcoming
consisting in the difficult transfer of the information,
which is associated to the taxa at a large geographic
scale, when one study is carried out at local level.
The ecological characterization of epiphytic
lichens, based on a quantitative approach and on
detailed field data collecting, is far from being
completed. Thus, the identification of groups of
species ecologically homogeneous, whose distribu-
tion could be used to mark bioclimatic borders, is a
difficult task to perform in a quantitative way on the
basis of the current knowledge of epiphytic lichens
ecology.
A first proposal to achieve this goal is presented as
a case-study, in which epiphytic lichen distribution
was estimated, through the application of a rigorous
sampling design and a standardized field strategy, in a
climatically heterogeneous area (Liguria, NW-Italy),
and distributional data were related to main climatic
parameters (namely rainfall and temperature) by
means of non-parametric multivariate models. In this
study a quantitative methodology is introduced to
identify groups of species with significantly similar
response to climatic variability. The study has three
main goals: (a) To quantify the relationship between
epiphytic lichens distribution and macroclimatic gra-
dients in the study area; (b) to provide a case study for
evaluating the possible use of epiphytic lichens as
bioclimatic indicators; (c) to compare the actual
bioclimatic subdivision of the study area (Nimis and
Martellos 2002), based on a synthetic and semi-
quantitative approach, with the one resulting from the
application of the proposed methodology.
Methods
Study area
The Liguria region, in north-western Italy, has a
surface area of 5,314 km2. To the south, it borders on
the Ligurian Sea. A continuous mountainous ridge
(Ligurian Alps and Ligurian Apennines) separates the
Tyhrrenian slopes from the Po Valley basin. Liguria
258 Plant Ecol (2008) 195:257–272
123

was selected as the study area because of the
occurrence of significant environmental gradients,
for both natural and anthropogenic variables. Accord-
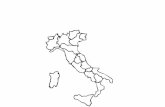
ing to Nimis (2003), the survey area can be subdi-
vided in four main bioclimatic units (Fig. 1A–D): (A)
Montane, including some Alpine and Apennine areas
of the hinterland above 1000 m, dominated by beech
forests; (B) Humid Sub-Mediterranean, occurring
both in the Tyhrrenian and Po valley hinterland
along the coast to the east of Genoa and to the
extreme west of Genoa near the French border,
ranging from 400 m to 1000 m and characterized by
deciduous Quercus-Carpinus forests; (C) Mediterra-
nean, limited to coastal districts (less than 10 km
from the sea) ranging from 0 m to 800 m and (D) Dry
Sub-Mediterranean, occurring mainly in the valleys
of the River Po in the western part of the region.
The population of about 1.5 million inhabitants is
concentrated in the regional capital, Genoa, and in
coastal areas (ca. 2000 persons/km2), where high
levels of atmospheric pollution occur, due mainly to
traffic and industry. On the other hand, many
mountainous areas are scarcely populated (ca. 10 per-
sons/km2) and have no local sources of air pollution.
Data collection
A total of 165 sampling sites were selected by means
of a stratified random sampling design, based on
habitat type and altitude, proportionally to the surface
occupied by each stratum within the survey area. At
each site, consisting of a 30-m radius plot, all suitable
trees (circumference > 60 cm, inclination of the
bole < 108, absence of damage and decorticated areas
on the trunk) were sampled. On each tree, the
frequencies of all epiphytic lichen species in a
sampling grid consisting of four 10 · 50 cm ladders,
each divided into five 10 · 10 cm squares were
considered. The grid was placed systematically on the
cardinal exposures (N, E, S, W) of the bole, at a
height of 1 m above ground, following the standards
suggested by Asta et al. (2002). Nomenclature
follows Nimis (2003).
To explore the relationship between lichen distri-
bution and macroclimatic factors, data were related to
average yearly rainfall (values ranging between
950 mm and 1800 mm) and average yearly temper-
ature (between 108C and 148C), provided by the
Ligurian Regional Council (1999) and calculated on
the basis of local models (Buzzi et al. 1994). These
two simple climatic parameters were selected on the
basis of previous studies (Giordani 2006, 2007) since
they have been proved to be related to epiphytic
lichen diversity in the survey area.
Data analysis
Species abundance was calculated for each plot as the
sum of frequencies of each species within the
sampling grids on a tree, averaged for all trees within
a plot. Quantitative data were subsequently converted
into binary (presence–absence) with respect to the
median of the distribution of the abundance of each
species in the whole dataset.
A
C D
B
Fig. 1 Bioclimatic units of Liguria region, according to Nimis (2003): Montane (A), Humid sub-Mediterranean (B), Mediterranean
(C), Dry sub-Mediterranean (D)
Plant Ecol (2008) 195:257–272 259
123

Two matrices were considered: (1) sample
units · species presence, and (2) sample units · cli-
matic variables. Quantitative variables of matrix 2
were log-transformed (McCune et al. 2002), in order
to reduce the effect of the high variability and of the
different order of magnitude within variables on the
final model. To detect and exclude from analysis
possible outliers, an explorative multivariate analysis
was carried out using PC-ORD method (McCune and
Mefford 1999). To describe the relationship between
species distribution and macroclimatic variables, the
transformed matrix of the whole dataset was analysed
by Non Parametric Multiplicative Regression—
NPMR (Bowman and Azzalini 1997; McCune et al.
2003), using the software Hyperniche 1.0 (McCune
and Mefford 2004) and the Species Occurrence
(SpOcc) NPMR model (McCune et al. 2003).
The model estimates the probability of occurrence
of a given species in a target site, by applying data
from sites that lie close (environmental neighbour-
hood) to the target one in the bi-dimensional space
defined by values of the two macroclimatic variables
(neighbourhood tolerance window).
For each species the best SpOcc-NPMR model
was considered, to describe the contribution of
rainfall and temperature in predicting its distribution.
Following McCune and Mefford (2004), the quality
of a SpOcc-NPMR model (M2) was estimated as its
improvement over an a priori model (M1), given by
the average frequency of the species in the whole
dataset. First, an equal probability was assumed for
both models, so that Pr(M1) = Pr(M2) = ½. Then,
Bayes factors (B12) were calculated as the ratio of the
likelihood of the observed values under the posterior
model M1 to the likelihood of the result under the
a priori model M2. As suggested by Kass and Raftery
(1995), the results were interpreted as log10(B12),
where values > 2 indicate a decisive relationship
between the response variable (species occurrence)
and the predictive ones (temperature and rainfall)
included in each model.
Furthermore, a model evaluation was carried out
with a Monte Carlo permutation test, by comparing
the estimation of the response variable given by the
selected models to the average estimation calculated
by 1,000 random permutations among the dataset.
Finally a matrix of sample units · species presence
was submitted to cluster analysis (Euclidean Distance
and Minimum Variance clustering algorithm), includ-
ing only the species significantly related to climatic
variables, to test if their similar response to macro-
climatic factors do correspond to their local distribu-
tion (i.e., abundance in the sampling sites).
Results
SpOcc-NPMR models
A total of 190 species were recorded in the 165
sampling sites. The abundance of 70 species, out of
the 190, resulted significantly related to yearly
average temperature and rainfall, when evaluated
with Monte–carlo test (P < 0.05). In particular (see
Table 1), 59 species (e.g., Buellia griseovirens)
showed significantly high log B (> 2), indicating a
strong relationship with the predictive variables
included in the model. A statistically significant
model was also found for other 11 species (e.g.,
Amandinea punctata), but, in these cases, a low log B
value (< 2) indicates a weaker relation with climatic
variables included in the model. Finally, 120 species
(e.g., Physcia aipolia) showed no relation with
climatic variables (P > 0.05; log B < 2).
Cluster analysis
Three groups were identified (Fig. 2):
• Cluster A includes 23 species, mainly related to
cold-humid climate (e.g., Parmelina pastillifera,
Hypogymnia physodes, Buellia griseovirens).
• Cluster B includes 18 species, related to humid
conditions, but with a broader ecological range
for temperature, occurring both in warm and in
fresh climate (e.g., Flavoparmelia caperata, Par-
motrema chinense, Normandina pulchella).
• Cluster C includes 18 species occurring in meso-
to-warm areas with humid to dry climate (e.g.,
Xanthoria parietina, Flavoparmelia soredians
Heterodermia obscurata, Parmotrema reticula-
tum).
Geographical distribution vs. ecological responses
of the species
The joint distribution of the species of each cluster is
reported at local scale in Fig. 3. The species of cluster
260 Plant Ecol (2008) 195:257–272
123

Table 1 List of the 190 species releved in the survey area
Taxon logB Cluster Code
Acrocordia gemmata (Ach.) A. Massal. < 2 001
Amandinea punctata (Hoffm.) Coppins & Scheid. < 2* 002
Anaptychia ciliaris (L.) Korb. < 2 003
Arthonia punctiformis Ach. < 2* 004
Arthonia radiata (Pers.) Ach. 3.8* A 005
Arthopyrenia cerasi (Schrad.) A. Massal. < 2 006
Arthrosporum populorum A. Massal. < 2 007
Bacidia arceutina (Ach.) Arnold < 2 008
Bacidia rosella (Pers.) De Not. < 2 009
Bacidia rubella (Hoffm.) A. Massal. < 2 010
Bactrospora patellarioides (Nyl.) Almq. var. patellarioides < 2 011
Bryoria fuscescens (Gyeln.) Brodo & D. Hawksw. < 2 012
Buellia disciformis (Fr.) Mudd < 2 013
Buellia griseovirens (Sm.) Almb. 9.1* A 014
Calicium abietinum Pers. < 2 015
Caloplaca cerina (Hedw.) Th. Fr. var. cerina < 2 016
Caloplaca ferruginea (Huds.) Th. Fr. 5.2* C 017
Caloplaca flavorubescens (Huds.) J. R. Laundon var. flavorubescens < 2 018
Caloplaca herbidella (Hue) H. Magn. < 2 019
Caloplaca luteoalba (Turner) Th. Fr. < 2 020
Caloplaca pollinii (A.Massal.) Jatta 7.7* C 021
Caloplaca pyracea (Ach.) Th. Fr. < 2* 022
Candelaria concolor (Dicks.) Stein < 2* 023
Candelariella reflexa (Nyl.) Lettau <2* 024
Candelariella xanthostigma (Ach.) Lettau < 2* 025
Canoparmelia crozalsiana (de Lesd.) Elix & Hale < 2 026
Catillaria nigroclavata (Nyl.) Schuler < 2 027
Catinaria atropurpurea (Schaer.) Vezda & Poelt < 2 028
Chrysothrix candelaris (L.) J. R. Laundon 2.9* C 029
Cladonia caespiticia (Pers.) Florke < 2 030
Cladonia coniocraea (Florke) Spreng. < 2 031
Cladonia fimbriata (L.) Fr. 2.5* C 032
Cladonia parasitica (Hoffm.) Hoffm. 2.2* B 033
Cladonia peziziformis (With.) J. R. Laundon < 2 034
Cladonia pyxidata (L.) Hoffm. < 2 035
Collema conglomeratum Hoffm. < 2 036
Collema flaccidum (Ach.) Ach. < 2 037
Collema fragrans (Sm.) Ach. < 2 038
Collema furfuraceum (Arnold) Du Rietz 4.2* C 039
Collema ligerinum (Hy) Harm. < 2 040
Collema multipunctatum Degel. < 2 041
Collema nigrescens (Huds.) DC. < 2 042
Collema subflaccidum Degel. < 2 043
Degelia atlantica (Degel.) M. Jørg. & P. James < 2 044
Degelia plumbea (Lightf.) M. Jørg. & P. James < 2 045
Plant Ecol (2008) 195:257–272 261
123

Table 1 continued
Taxon logB Cluster Code
Dendriscocaulon umhausense (Auersw.) Degel. < 2 046
Dimerella pineti (Ach.) Vezda < 2 047
Evernia prunastri (L.) Ach. 2.6* A 048
Flavoparmelia caperata (L.) Hale 2.0* B 049
Flavoparmelia soredians (Nyl.) Hale 3.7* C 050
Fuscidea stiriaca (A. Massal.) Hafellner < 2 051
Fuscopannaria mediterranea (Tav.) M. Jørg. < 2 052
Graphis scripta (L.) Ach. < 2 053
Gyalecta liguriensis (Vezda) Vezda < 2 054
Gyalecta truncigena (Ach.) Hepp < 2 055
Heterodermia obscurata (Nyl.) Trevis. 7.6* C 056
Hyperphyscia adglutinata (Florke) H. Mayrhofer & Poelt 6.1* B 057
Hypocenomyce scalaris (Ach.) M. Choisy < 2 058
Hypogymnia physodes (L.) Nyl. 3.0* A 059
Hypogymnia tubulosa (Schaer.) Hav. < 2 060
Hypotrachyna revoluta (Florke) Hale < 2 061
Japewiella carrollii (Coppins & P. James) Printzen < 2 062
Koerberia biformis A. Massal. < 2 063
Lecania cyrtella (Ach.) Th. Fr. < 2 064
Lecania fuscella (Schaer.) A. Massal. < 2 065
Lecania naegelii (Hepp) Diederich & Van den Boom < 2 066
Lecanographa amylacea (Pers.) Egea & Torrente < 2 067
Lecanora albella (Pers.) Ach. 6.2* A 068
Lecanora allophana Nyl. < 2 069
Lecanora argentata (Ach.) Malme 3.8* A 070
Lecanora carpinea (L.) Vain. 2.5* A 071
Lecanora chlarotera Nyl. < 2* 072
Lecanora cinereofusca H. Magn. < 2 073
Lecanora expallens Ach. < 2* 074
Lecanora glabrata (Ach.) Malme < 2 075
Lecanora hagenii (Ach.) Ach. var. hagenii < 2 076
Lecanora horiza (Ach.) Linds. < 2 077
Lecanora intumescens (Rebent.) Rabenh. 6.7* A 078
Lecanora leptyrodes (Nyl.) Degel. < 2 079
Lecanora meridionalis H. Magn. < 2 080
Lecanora pulicaris (Pers.) Ach. 4.6* A 081
Lecanora rubicunda Bagl. < 2 082
Lecanora saligna (Schrad.) Zahlbr. < 2 083
Lecanora strobilina (Spreng.) Kieff. 3.4* A 084
Lecanora subintricata (Nyl.) Th. Fr. < 2 085
Lecanora symmicta (Ach.) Ach. < 2 086
Lecidea turgidula Fr. < 2 087
Lecidella elaeochroma (Ach.) M. Choisy 2.1* B 088
Lepraria incana (L.) Ach. 3.7* B 089
262 Plant Ecol (2008) 195:257–272
123

Table 1 continued
Taxon logB Cluster Code
Lepraria lobificans Nyl. < 2 090
Leprocaulon microscopicum (Vill.) Gams 9.1* B 091
Leptogium cyanescens (Rabenh.) Korb. < 2 092
Leptogium lichenoides (L.) Zahlbr. < 2 093
Leptogium tenuissimum (Dicks.) Korb. < 2 094
Leptogium teretiusculum (Wallr.) Arnold < 2 095
Leptorhaphis atomaria (Ach.) Szatala < 2 096
Lobaria amplissima (Scop.) Forssell < 2 097
Lobaria pulmonaria (L.) Hoffm. < 2 098
Melanelia elegantula (Zahlbr.) Essl. < 2 099
Melanelia exasperata (De Not.) Essl. < 2 100
Melanelia exasperatula (Nyl.) Essl. < 2 101
Melanelia fuliginosa (Duby) Essl. subsp. glabratula 2.8* B 102
Melanelia glabra (Schaer.) Essl. < 2 103
Melanelia laciniatula (H. Olivier) Essl. < 2 104
Melanelia subaurifera (Nyl.) Essl. 2.4* A 105
Melaspilea urceolata (Fr.) Almb. < 2 106
Micarea prasina Fr. < 2 107
Mycobilimbia pilularis (Korb.) Hafellner & Turk < 2 108
Mycomicrothelia confusa D. Hawksw. < 2 109
Naetrocymbe punctiformis (Pers.) R. C. Harris < 2 110
Nephroma laevigatum Ach. < 2 111
Nephroma parile (Ach.) Ach. < 2 112
Normandina pulchella (Borrer) Nyl. 4.9* B 113
Ochrolechia subviridis (Høeg) Erichsen < 2 114
Opegrapha atra Pers. 3.2* C 115
Opegrapha culmigena Lib. < 2 116
Opegrapha varia Pers. < 2 117
Pachyphiale arbuti (Bagl.) Arnold < 2 118
Pannaria conoplea (Ach.) Bory < 2 119
Parmelia saxatilis (L.) Ach. 7.9* A 120
Parmelia submontana Hale < 2 121
Parmelia sulcata Taylor 3.8* B 122
Parmeliella triptophylla (Ach.) Mull. Arg. < 2 123
Parmelina pastillifera (Harm.) Hale 7.8* A 124
Parmelina quercina (Willd.) Hale 2.5* A 125
Parmelina tiliacea (Hoffm.) Hale 2.6* B 126
Parmelinopsis horrescens (Taylor) Elix & Hale < 2 127
Parmeliopsis ambigua (Wulfen) Nyl. 4.9* A 128
Parmeliopsis hyperopta (Ach.) Arnold < 2 129
Parmotrema chinense (Osbeck) Hale & Ahti 8.2* B 130
Parmotrema crinitum (Ach.) M. Choisy < 2 131
Parmotrema reticulatum (Taylor) M. Choisy 8.4* C 132
Parmotrema stuppeum (Taylor) Hale < 2 133
Plant Ecol (2008) 195:257–272 263
123

Table 1 continued
Taxon logB Cluster Code
Pertusaria albescens (Huds.) M. Choisy & Werner 4.2* A 134
Pertusaria amara (Ach.) Nyl. 3.1* B 135
Pertusaria coccodes (Ach.) Nyl. < 2 136
Pertusaria coronata (Ach.) Th. Fr. < 2 137
Pertusaria flavida (DC.) J. R. Laundon 3.3* A 138
Pertusaria hemisphaerica (Florke) Erichsen 4.7* A 139
Pertusaria hymenea (Ach.) Schaer. 4.2* C 140
Pertusaria leioplaca DC. 2.7* A 141
Pertusaria pertusa (Weigel) Tuck. 2.4* B 142
Pertusaria pustulata (Ach.) Duby < 2* 143
Phaeophyscia chloantha (Ach.) Moberg < 2 144
Phaeophyscia endophoenicea (Harm.) Moberg < 2 145
Phaeophyscia hirsuta (Mereschk.) Essl. 2.8* C 146
Phaeophyscia nigricans (Florke) Moberg < 2 147
Phaeophyscia orbicularis (Neck.) Moberg 3.8* B 148
Phaeophyscia pusilloides (Zahlbr.) Essl. < 2 149
Phlyctis agelaea (Ach.) Flot. < 2 150
Phlyctis argena (Spreng.) Flot. 6.0* B 151
Physcia adscendens (Fr.) H. Olivier 3.1* B 152
Physcia aipolia (Humb.) Furnrh. < 2 153
Physcia biziana (A. Massal.) Zahlbr. var. biziana < 2 154
Physcia clementei (Turner) Maas Geest. 3.3* C 155
Physcia leptalea (Ach.) DC. < 2 156
Physcia stellaris (L.) Nyl. 4.3* A 157
Physcia tenella (Scop.) DC. < 2* 158
Physcia tribacioides Nyl. < 2 159
Physcia vitii Nadv. 2.8* C 160
Physconia distorta (With.) J. R. Laundon 2.2* C 161
Physconia grisea (Lam.) Poelt subsp. grisea 2.4* C 162
Physconia servitii (Nadv.) Poelt < 2 163
Pleurosticta acetabulum (Neck.) Elix & Lumbsch < 2 164
Porina borreri (Trevis.) D. Hawksw. & P. James < 2 165
Pseudevernia furfuracea (L.) Zopf var. furfuracea < 2 166
Punctelia borreri (Sm.) Krog 3.1* C 167
Punctelia perreticulata (Rasanen) G. Wilh. & Ladd < 2 168
Punctelia subrudecta (Nyl.) Krog 5.7* B 169
Punctelia ulophylla (Ach.) van Herk & Aptroot < 2 170
Pyrenula chlorospila Arnold < 2 171
Pyrenula nitida (Weigel) Ach. 3.2* C 172
Pyrenula nitidella (Schaer.) Mull. Arg. < 2 173
Pyxine subcinerea Stirt. < 2 174
Ramalina farinacea (L.) Ach. < 2 175
Ramalina fraxinea (L.) Ach. < 2* 176
Rinodina nimisii Giralt & H. Mayrhofer < 2 177
264 Plant Ecol (2008) 195:257–272
123

A (Fig. 3A) show a mainly montane distribution in
the survey area (Eastern Ligurian Apennines and
Maritime Alps). Cluster B (Fig. 3B) includes lichens
that are mainly located in hinterland areas of the
Eastern part of the region. The species of cluster C
are distributed along a narrow coastal belt both in the
Eastern and in the Western Riviera (Fig. 3C).
These three local areas could be intended as
bioclimatic homogeneous units once it is shown that
species grouped on the basis of their local distribution
do have similar ecological response to climatic
variables.
SpOcc results, modelling the ecological response
to macroclimatic factors, are shown in Fig. 4 for case-
species of each group of the cluster analysis. The
NPMR model of Amandinea punctata (Fig. 4A) is
also reported, as an example for species not distrib-
uted in accordance with the pattern of macroclimatic
variables. In this case the probability of occurrence of
the species is relatively high throughout the ecolog-
ical space, so that it is not likely to give relevant
information on the bioclimatic variability in the
survey area.
Model for Buellia griseovirens is representative
for the ecological response of species of cluster A
Fig. 4B). It shows the higher probability of occur-
rence for average yearly temperature ranging
between 108C and 118C and for average yearly
rainfall higher than 1600 mm. These ecological
ranges do correspond to the montane distributional
pattern (Fig. 3A).
Cluster B includes a more ecologically heteroge-
neous subset of species. Some of them (e.g.,
Pertusaria amara—Fig. 4C) show high probability
of occurrence for meso- to warm conditions (from
118C to 158C) and high yearly average rainfall (more
than 1800 mm). Other species (e.g., Flavoparmelia
caperata—Fig. 4D) have a broader ecological range.
This corresponds to a distribution pattern mainly
located in sub-mediterranean areas of the survey area
(Fig. 3B).
All the taxa of cluster C resulted associated with
warm conditions (yearly average temperature ranging
from 13 to 158C), but two sub-cluster may be found.
The former one includes species related to low yearly
average rainfall, e.g., Flavoparmelia soredians,
showing the higher probability of occurrence for
average yearly rainfall lower than 1000 mm
(Fig. 4E), whilst the latter one groups together
thermophilous species related to very humid condi-
tions (e.g., Heterodermia obscurata—Fig. 4F), cor-
responding to average yearly rainfall higher than
Table 1 continued
Taxon logB Cluster Code
Rinodina pyrina (Ach.) Arnold 3.7* A 178
Rinodina sophodes (Ach.) A. Massal. < 2 179
Schismatomma decolorans (Sm.) Clauzade & Vezda 4.2* B 180
Schismatomma graphidioides (Leight.) Zahlbr. < 2 181
Scoliciosporum chlorococcum (Stenh.) Vezda < 2 182
Scoliciosporum umbrinum (Ach.) Arnold 8.5* A 183
Tephromela atra (Huds.) Hafellner var. torulosa (Flot.) Hafellner 4.3* A 184
Thelopsis rubella Nyl. < 2 185
Usnea florida (L.) F. H. Wigg. < 2 186
Usnea hirta (L.) F. H. Wigg. < 2 187
Vulpicida pinastri (Scop.) J. E. Mattsson & M. J. Lai < 2 188
Xanthoria fallax (Hepp) Arnold < 2 189
Xanthoria parietina (L.) Th. Fr. 2.6* C 190
Nomenclature follows Nimis (2003). log B column indicates the evaluation of the NPMR model for species abundance in the
sampling sites. Values for species with distribution strongly related to the pattern of predictive climatic variables (log B > 2, see text)
are in bold
* P (log B) < 0.05. For each taxon the local distributional cluster and an identifying code are also reported (see Fig. 2)
Plant Ecol (2008) 195:257–272 265
123

1600 mm. Irrespectively to their water requirement,
the ecological response of the species of cluster C do
correspond to a mediterranean distributional pattern,
along the coasts of the survey area (Fig. 3C).
Discussion
Though epiphytic lichens distribution depends on a
complex set of environmental and substrate-related
explanatory variables, acting from the tree to the
landscape level (Nimis et al. 2002), many authors
pointed out the role of climatic factors (McCune et al.
1997; Goward and Spribille 2005; Hauck and Spri-
bille 2005; Giordani 2006, 2007).
Interpretation of SpOcc models
About 30% of the epiphytic lichen flora (59 species
out of 190) resulted significantly related to yearly
average temperature and rainfall patterns. The sig-
nificance of climate-dependence for the selected
species is based on two main considerations: (a)
all these taxa showed significantly high log B (> 2),
and (b) log B values resulted significant (P < 0.05)
when evaluated with Monte–carlo test. Under these
conditions, it is assumed that the distribution of all
these taxa does not depend significantly on factors
other than climatic, a part from those covariating:
some nitrophitic species of cluster C (e.g., Phaeo-
physcia hirsuta, Physcia vitii, Physconia distorta,
A005A078A139A070A068A124A157A134A138A178A014A183A081A059A128A048A105A120A184A084A071A141A125B151B088B049B130B102B122B126B135B169B033B142B057B152B148B089B091B113B180C017C021C160C039C146C050C161C190C162C029C140C167C056C132C155C032C115C172
100
3.8e-02
75
4.1e+00
50
8.2e+00
25
1.2e+01
0
1.6e+02
Information Remaining (%)
Distance (ObjectiveFunction)
A
B
C
Fig. 2 Dendrogram of the
species based on their
abundance in the sampling
sites. Species are indicated
with the cluster code (from
A to C) followed by the
identifying codes of Table 1
266 Plant Ecol (2008) 195:257–272
123

Physconia grisea and Xanthoria parietina) are mostly
distributed in the south part of the survey area, where
both temperature and NOx pollution are higher.
A complete analysis of the non climatic factors
affecting epiphytic lichen distribution in the survey
area, such as substrate- or pollution-related ones, is
not an aim of the present work and it is addressed
elsewhere (Giordani 2007).
Selecting guilds of indicator species to assess
bioclimatic features of a territory
Whilst several authors provided results on the
relationship between total epiphytic lichen diversity
and bioclime (Fos 1998; Loppi et al. 2002), in the
present work we focussed the attention on distribu-
tional and ecological features of guilds of species,
statistically selected with reference to their relation to
climatic variables.
The use of guilds of species instead of diversity as
a whole was suggested by Will-Wolf et al. (2002) as a
valid tool for assessing peculiar ecological condi-
tions. A similar approach was recently applied to
various organisms in different regions (e.g., Pyke and
Fischer 2005). Some authors select guilds a priori, on
the basis of well-known ecological requirements or
distribution (Goward and Spribille 2005). We con-
sider that this approach could be biassed by an
incorrect (subjective) use of ‘well-known’ informa-
tion, which should be referred to the specific context
in which it was acquired. The use of an a posteriori
methodology (Karlsen and Elvebakk 2003; Karlsen
et al. 2005), in which a guild is defined as a group of
taxa whose distribution is significantly related to the
pattern of one or more climatic factors, is comple-
mentary to the first approach: in our work, it lays on a
probability-based sampling dataset and, conse-
quently, the level of uncertainty and the statistic
significance of the results are known and the inter-
pretative step is free from the level of floristic
investigation of the area. Nevertheless other con-
straints of the ‘guild approach’ are not completely
solved by our method, including the lack of detailed
information about the complex eco-physiology of the
selected species, which may result in a weak
correlation between ‘indicators’ and response vari-
able (Elzinga et al. 2001).
Ecological and ecophysiological requirements
of the selected species
The guilds derived from SpOcc models includes
some of the most studied species in lichen ecology
and eco-physiology (e.g., Jensen 2002). The out-
comes referred to those species can be used to better
understand the relationship between lichen and
climatic factors distribution.
Honegger (1995) pointed out the remarkable
regenerative capacity of Xanthoria parietina (cluster
C: species of warm areas) vs. Parmelia sulcata
(cluster B: mesothermic and humid), in relation to
drought and temperature stress. Baruffo and Tretiach
(2005) showed that Parmelia saxatilis (cluster A—
100%50%10%
Sum of frequencies
100%50%10%
Sum of frequencies
100%50%10%
Sum of frequencies
A
B
C
Fig. 3 Distributional pattern of the species in the Study Area.
Each map (A–C) shows the joint distribution and the relative
abundance of all the species of the corresponding cluster in Fig.
1. The abundance is measured as the sum of frequencies of all
the species in each sampling site, expressed in percentage on
the maximum observed value
Plant Ecol (2008) 195:257–272 267
123

cold-humid), showed best photosynthetic parameters
in beech forests, whilst Parmotrema chinense (cluster
B—mesothermic-humid) had its optimal performance
in Quercus forests, rapidly decreasing at medium to
high elevations.
According to Caviglia et al. (2001) and Caviglia
and Modenesi (1993) Flavoparmelia soredians and
Parmotrema reticulatum (cluster C) are more reactive
than F. caperata (cluster B) to seasonal fluctuations
of oxidative stresses, showing an increased produc-
tion of antioxidative lichen substances during the
warmest periods, suggesting that these species, in
their bioclimatic optimum, are able to promptly react
to climate-related stresses.
Yearly average rainfall (mm)
Yea
rly
aver
age
tem
per
atu
re (
°C)
800 1000 1200 1400 1600 1800 2000
9
10
11
12
13
14
15
16
Yearly average rainfall (mm)
800 1000 1200 1400 1600 1800 2000
9
10
11
12
13
14
15
16 SpOcc-NPMR for Amandinea punctata SpOcc-NPMR for Buellia griseovirens
Probabilityof SpOcc
0.00
0.25
0.50
0.75
1.00
Probabilityof SpOcc
0.00
0.25
0.50
0.75
1.00
Yearly average rainfall (mm)
Yea
rly
aver
age
tem
per
atu
re (
°C)
800 1000 1200 1400 1600 1800 2000
9
10
11
12
13
14
15
16 SpOcc-NPMR for Heterodermia obscurata
Probabilityof SpOcc
0.00
0.25
0.50
0.75
1.00
Yearly average rainfall (mm)
Yea
rly
aver
age
tem
per
atu
re (
°C)
800 1000 1200 1400 1600 1800 2000
9
10
11
12
13
14
15
16SpOcc-NPMR for Flavoparmelia soredians
Probabilityof SpOcc
0.00
0.25
0.50
0.75
1.00
Yearly average rainfall (mm)
Yea
rly
aver
age
tem
per
atu
re (
°C)
800 1000 1200 1400 1600 1800 2000
9
10
11
12
13
14
15
16 SpOcc-NPMR for Flavoparmelia caperata
Probabilityof SpOcc
0.00
0.25
0.50
0.75
1.00
Yearly average rainfall (mm)
Yea
rly
aver
age
tem
per
atu
re (
°C)
Yea
rly
aver
age
tem
per
atu
re (
°C)
800 1000 1200 1400 1600 1800 2000
9
10
11
12
13
14
15
16 SpOcc-NPMR for Pertusaria amara
Probabilityof SpOcc
0.00
0.25
0.50
0.75
1.00
A
C
E F
D
B
Fig. 4 Non parametric multiregressive models (NPMR) for
species occurrence (SpOcc) in the ecological space defined by
the climatic predictive variables (yearly average temperature
and rainfall) in the Study Area. Models for Amandinea
punctata (A), Buellia griseovirens (B), Pertusaria amara(C), Flavoparmelia caperata (D), Flavoparmelia soredians (E)
and Heterodermia obscurata (F) are shown
268 Plant Ecol (2008) 195:257–272
123

Comparing different methods for delimitating
bioclimatic regions
After the results of this work, we were able to define
three bioclimatic units, on the basis of the joint
distribution of three guilds of species, significantly
sensitive to yearly average temperature and rainfall.
Their distributional patterns identify areas showing a
good accordance when compared with the biocli-
matic units sensu Nimis and Tretiach (1995) and
Nimis (2003), based on precipitations and altitudinal
belts (Fig. 1) and proposed at a national scale. The
following correspondences can be observed:
(1) Species of cluster A (e.g., B. griseovirens) are
distributed in an area corresponding to the
Montane bioclimatic unit (Fig. 1A);
(2) Distributional pattern of Cluster B fits rather
well with the Humid sub-Mediterranean unit
(Fig. 1B) sensu Nimis (2003);
(3) Species of cluster C generally show a good
accordance with the Mediterranean bioclimatic
unit (Fig. 1C). The heterogeneity of these
species in respect to rainfall requirement may
lead to a further localization of a Dry Mediter-
ranean unit, which was already proposed for
other Italian regions, but that was not described
by Nimis (2003) for Liguria.
(4) Our analysis does not evidence a cluster of
species related to the Dry sub-Mediterranean
unit sensu Nimis, being this area characterized
in Liguria by a very poor epiphytic lichen flora.
These results point out also some limits of small-
scale delimitation of bioclimatic units of Italy, which
can generate fictitious regions whose borderlines
cannot be easily defined (Nimis and Tretiach 2004):
the two Tyhrrenian units sensu Nimis do not strictly
correspond to the peninsular West-sided coasts: due
to local geographic, morphologic and climatic fea-
tures of the territory some ‘‘bioclimatic enclaves’’ are
likely to occur.
These limits could be overcome by referring
bioclimatic observations to an higher geographic
scale such as at regional (as in this article) or local
level. We agree with Walther et al. (2002) that
regional changes, being spatially heterogeneous, are
relevant in the context of ecological response to
climatic change. Nevertheless, on the way of dispos-
ing of a much more detailed and precise bioclimatic
subdivision of Italy, a deeper knowledge of lichens
local distribution is needed, as showed by the
increasing number of floristic and vegetational stud-
ies recently carried out throughout the country at
regional scale (Brunialti and Giordani 2003; Nimis
and Tretiach 2004).
Baseline for forthcoming monitoring of the
effects of global change using epiphytic lichens
In a long-term perspective, our results provide a
baseline to investigate the possible effects on lichen
communities caused by global changes. Even if a
broad range of organisms were taken into account as
reliable indicators of climatic changes (Hughes 2000;
Walther et al. 2002), e.g., birds (Crick et al. 1997),
amphibians (Beebee 1995), flowering plants (Menzel
and Estrella 2001; Karlsen et al. 2005) and bryo-
phytes (Gignac 2001), there are few studies dealing
with possible applications with lichens (Insarov et al.
1999; Lucking 2003), thought the peculiar physiol-
ogy of these organisms, directly linked to atmosphere
for water supply (Rundel 1988; Nash 1996), should
increase the scientific interest in this sense. Our
outcomes, in particular, are in accord with those of
Van Herk et al. (2002) which observed fluctuations in
the epiphytic flora of The Netherlands from 1979 to
2001, related to an increasing of average temperature:
Five species of cluster A (Evernia prunastri, Hypo-
gymnia physodes, Lecanora argentata, Parmelia
saxatilis and Pertusaria albescens) and not those of
the other clusters showed in the Netherlands a
decreasing in the last years, whilst four species of
cluster C (Xanthoria parietina, Physconia grisea,
Micarea prasina and Punctelia borreri) and 8 of B
showed a significant (Flavoparmelia caperata, Lec-
idella elaeochroma, Punctelia subrudecta, Parmelia
sulcata, Physcia adscendens and Phaeophyscia or-
bicularis) and in some cases very rapid (Parmotrema
chinense and Hyperphyscia adglutinata) spreading.
As such, we can hypothesize that these species
could respond to climatic changes also at larger
scale than the one referring to Liguria region and in
relative short time framework. Nevertheless, we
should consider that the Netherlands’s original
lichen flora was nearly completely lost due to air
pollution (van Dobben et al. 2001; van Herk 2001),
so that, as Walther et al. (2002) pointed out, in
altered environments colonization of new species
Plant Ecol (2008) 195:257–272 269
123

well adapted to changed climatic conditions was
possible because of a low competitive pressure.
Probably this is not the case of our survey area,
where several ecosystems are still preserved.
Anyway the use of guild of epiphytic lichens as
climatic changes indicators could be tested only
providing a baseline of species occurrence at a given
time, once the species are selected on the basis of a
significant response to climate. In the case of Liguria,
for instance, one could plan to evaluate the future
increasing of warm-humid-related species of cluster
B in low diversity areas, as the urban and suburban
areas near the capital town Genoa or the dry sub-
Mediterranean areas in the western Riviera. Never-
theless, in our opinion, the reliability of epiphytic
lichens as tools for monitoring climatic changes in a
long-term perspective is still to demonstrate: future
studies should address the question if it would be
possible to obtain useful information about climatic
changes at ecosystem level, notwithstanding the
complex poikylohydric nature of these organisms,
which are probably related to micro- rather than
macroclimatic parameters.
Conclusions
The distribution of more than 30% of the releved
epiphytic species has been shown to be explained by
macroclimatic variables. A significant subset of
epiphytic lichens in the study area have been proved
to be efficient bioclimatic indicator for montane,
humid sub-Mediterranean and Mediterranean units,
whereas we have no evidences of a relation between a
dry sub-Mediterranean unit and epiphytic lichen
distribution, being this area characterized only by
the absence of indicator species.
Forthcoming studies will test at national scale this
quantitative approach for evaluating bioclimatic sub-
division of the country. It would be also possible to
model the likelihood of occurrence of indicator
species along the time by means of time series
analysis of macroclimatic variables. Furthermore,
applicative perspectives of this work include (a) a
better interpretation of biomonitoring data, thanks to
an improved knowledge of maximum potential
distribution and frequency of epiphytic lichens
throughout the region, and (b) a plan of long-term
monitoring survey to detect possible effects of global
changes on epiphytic vegetation.
Acknowledgement This work has been supported, in part, by
Ministero dell’Istruzione, dell’Universita e della Ricerca,
project F.I.S.R.—M.I.C.E.N.A., 2006.
References
Asta J, Erhard W, Ferretti M et al (2002) Mapping lichen
diversity as an indicator of environmental quality. In:
Nimis PL, Scheidegger C, Wolseley P (eds) Monitoring
with lichens: monitoring lichens. Kluwer, Dordrecht
Barkman JJ (1958) Phytosociology and Ecology of Cryptoga-
mic Epiphytes. Van Gorcum, Assen
Baruffo L, Tretiach M (2005) Variazioni stagionali dei para-
metri di fluorescenza clorofilliana in licheni lungo un
gradiente altitudinale. Not Soc Lich Ital 18:20–21
Beebee TJC (1995) Amphibian breeding and climate. Nature
374:219–220
Bowman AW, Azzalini A (1997) Applied smoothing tech-
niques for data analysis. Clarendon Press, Oxford
Brunialti G, Giordani P (2003) Variability of lichen diversity in
a climatically heterogeneous area (Liguria, NW Italy).
Lichenologist 35:55–69
Buzzi A, Fantini M, Malguzzi P et al (1994) Validation of
Limited Area Model in cases of Mediterranean cyclo-
genesis: surface fields and precipitation scores. Meteorol
Atmos Phys 53:137–153
Caviglia AM, Modenesi P (1993) Oxidative stress and ascorbic
acid contents in Parmotrema reticulatum and Parmeliasulcata thalli. Lichenologist 31:105–110
Caviglia AM, Nicora P, Giordani P et al (2001) Oxidative
stress and usnic acid content in Parmelia caperata and
Parmelia soredians (Lichenes). Il Farmaco 56:379–380
Coxson DS, Coyle M (2003) Niche partitioning and photo-
synthetic response of alectorioid lichens from subalpine
spruce-forest in north-central British Columbia, Canada:
the role of canopy microclimate gradients. Lichenologist
35:157–175
Crick HQP, Dudley C, Glue DE et al (1997) UK birds are
laying eggs earlier. Nature 388:526
Degelius G (1935) Das ozeanische element der Strauch-und
Laubflechtenflora von Skandinavien. Acta Phytogeogr
Suec 7:1–411
Elzinga CL, Salzer DW, Willoughby JW et al (2001) Moni-
toring plant and animal populations. Blackwell Scientific
Publications, Oxford
Fos S (1998) Lıquenes epıfitos de los alcornocales ibericos:
correlaciones bioclimaticas, anatomicas y densiometricas
con el corcho de reproduccion. Guineana 4:1–507
Fos S, Deltoro VI, Calatayud A et al (1999) Changes in water
economy in relation to anatomical and morphological
characteristics during thallus development in Parmeliaacetabulum. Lichenologist 31:375–387
Gignac LD (2001) Bryophytes as indicators of climate change.
Bryologist 104:410–420
270 Plant Ecol (2008) 195:257–272
123

Giordani P (2006) Variables influencing the distribution of
epiphytic lichens in heterogeneous areas: a case-study for
Liguria (NW-Italy). J Veg Sci 17:195–206
Giordani P (2007) Is the diversity of epiphytic lichens a reli-
able indicator of air pollution? A case study from Italy.
Env Poll 146:317–323
Goward T, Spribille T (2005) Lichenological evidence for the
recognition of inland rain forests in western North
America. J Biogeogr 32:1209–1219
Hauck M, Spribille T (2005) The significance of precipitation
and substrate chemistry for epiphytic lichen diversity in
spruce-fir forests of the Salish Mountains, Montana. Flora
200:547–562
Honegger R (1995) Experimental studies with foliose macr-
olichens: fungal responses to spatial disturbance at the
organismic level and to spatial problems at the cellular
level during drought stress events. Can J Bot 73:569–
578
Hughes L (2000) Biological consequences of global warming:
is the signal already apparent? Trends Ecol Evol 15:56–61
Insarov GE, Semenov SM, Insarova ID (1999) A system to
monitor climate change with epilithic lichens. Environ
Monitoring Assessment 55:279–298
Jensen M (2002) Measurement of chlorophyll fluorescence in
lichens. In: Kranner I, Beckett RP, Varma AK (eds)
Protocols in lichenology. culturing, biochemistry, eco-
physiology and use in biomonitoring. Springer, Berlin
Kappen L (1988) Ecophysiological Relationships in Different
Climatic Regions. In: Galun M (ed) CRC handbook of
Lichenology, vol 2. CRC Press, Boca Raton, pp 37–100
Karlsen SR, Elvebakk A (2003) A method using indicators
plants to map local climatic variation in the Kangerlus-
suaq/Scoresby Sund area, East Greenland. J Biogeogr
30:1469–1491
Karlsen SR, Elvebakk A, Johansen B (2005) A vegetation-
based method to map climatic variation in the arctic-
boreal transition area of Finnmark, north-easternmost
Norway. J Biogeogr 32:1161–1186
Kass RE, Raftery AE (1995) Bayes factors. J Am Stat Assoc
90:773–795
Ligurian Regional Council (1999) Piano di risanamento e tu-
tela della qualita dell’aria. Regione Liguria, Genova
Loppi S, Giordani P, Brunialti G et al (2002) Identifying
deviations from naturality of lichen diversity for bioindi-
cation purposes. In: Nimis PL, Scheidegger C, Wolseley P
(eds) Monitoring with lichens: monitoring lichens. Klu-
wer, Dordrecht
Lucking R (2003) Takhtajan’s floristic regions and follicolous
lichen biogeography: a compatibility analysis. Lichenol-
ogist 35:33–54
McCune B, Mefford MJ (2004) Nonparametric multiplicative
habitat modeling. MjM Software, Gleneden Beach
McCune B, Mefford MJ (1999) Multivariate analysis of eco-
logical data. MjM Software, Gleneden Beach
McCune B, Berryman SD, Cissel JH et al (2003) Use of a
smoother to forecast occurrence of epiphytic lichens
under alternative forest management plans. Ecol Appl
13:1110–1123
McCune B, Dey JD, Peck JE et al (1997) Repeatability of
community data: species richness versus gradient scores
in large-scale lichen studies. Bryologist 100:40–46
McCune B, Grace JB, Urban DL (2002) Analysis of ecological
communities. MjM Software, Gleneden Beach
McCune B (1984) Lichens with oceanic affinities in the Bit-
terroot Mountains of Montana and Idaho. Bryologist
87:44–50
Menzel A, Estrella N, Fabian P (2001) Spatial and temporal
variability of the phenological seasons in Germany from
1951 to 1996. Glob Change Biol 7:657–666
Nash III TH (1996) Lichen biology. Cambridge University
Press, Cambridge
Nimis PL (2003) Checklist of the Lichens of Italy 3.0. Uni-
versity of Trieste, Dept of Biology. http://
dbiodbs.univ.trieste.it. Cited 1 Jun 2006
Nimis PL, Losi L (1984) Lichens as phytoclimatic indicators in
the Trieste Karst. Gortania 5:63–80
Nimis P.L., Martellos S. (2002) ITALIC – The information
system on Italian lichens. Bibl Lichenol 82: 271–283
Nimis PL, Tretiach M (1995) The lichens of Italy. A phyto-
climatic outline. Cryptog Bot 5:199–208
Nimis PL Tretiach M (2004) Delimiting Tyrrhenian Italy: A
lichen foray in the SW part of the peninsula. In: Dobbeler
P, Rambold G (eds) Festschrift hannes hartel: contribu-
tions to lichenology, pp 453–466
Nimis PL, Scheidegger C, Wolseley P (eds) (2002) Monitoring
with lichens – Monitoring lichens. Kluwer, Dordrecht
Pyke CR, Fischer DT (2005) Selection of bioclimatically rep-
resentative biological reserve systems under climate. Biol
Cons 121:429–441
Rose F (1976) Lichenological indicators of age and environ-
mental continuity in woodlands. In: Brown DH, Hawks-
worth DL, Bailey RH (eds) Lichenology: progress and
problems. Academic Press, London, pp 278–307
Rundel PW (1988) Water relation. In: Galun M (ed) CRC
handbook of Lichenology, vol 2. CRC Press, Boca Raton,
pp 17–36
Sancho LG, Schroeter B, Valladares F (1997) Photosynthetic
performance of two closely related Umbilicaria species in
central Spain: temperature as a key factor. Lichenologist
29:67–82
Shirazi AM, Muir PM, McCune B (1996) Environmental fac-
tors influencing the distribution of the lichens Lobariaoregana and L. pulmonaria. Bryologist 99:12–18
van Dobben HF, Wolterbeek HT, Wamelink GWW et al (2001)
Relationship between epiphytic lichens, trace elements and
gaseous atmospheric pollutants. Env Poll 112:163–169
van Herk CM (2001) Bark pH and susceptibility to toxic air
pollutants as independent causes of changes in epiphytic
lichen composition in space and time. Lichenologist
33:419–441
van Herk CM, Aptroot A, van Dobben HF (2002) Long-termmonitoring in the Netherlands suggests that lichens re-
spond to global warming. Lichenologist 34:141–154
Walther GR, Post E, Convey P et al (2002) Ecological re-
sponses to recent climate change. Nature 416:389–395
Plant Ecol (2008) 195:257–272 271
123

Will-Wolf S, Scheidegger C, McCune B (2002) Methods for
monitoring biodiversity and ecosystem function. Moni-
toring scenarios, sampling strategies and data quality.
In: Nimis PL, Scheidegger C, Wolseley P (eds) Moni-
toring with lichens: monitoring lichens. Kluwer, Dordr-
echt
Wirth V (1991) Zeigerwerte von Flechten. In: Ellenberg H,
Weber HE, Dull R et al (eds) Zeigerwerte von P?anzen in
Mitteleuropa. Scripta Geobotanica 18 215–237
272 Plant Ecol (2008) 195:257–272
123