
5 Results - shodhganga.inflibnet.ac.inshodhganga.inflibnet.ac.in/bitstream/10603/6303/13/13_chapter...
48
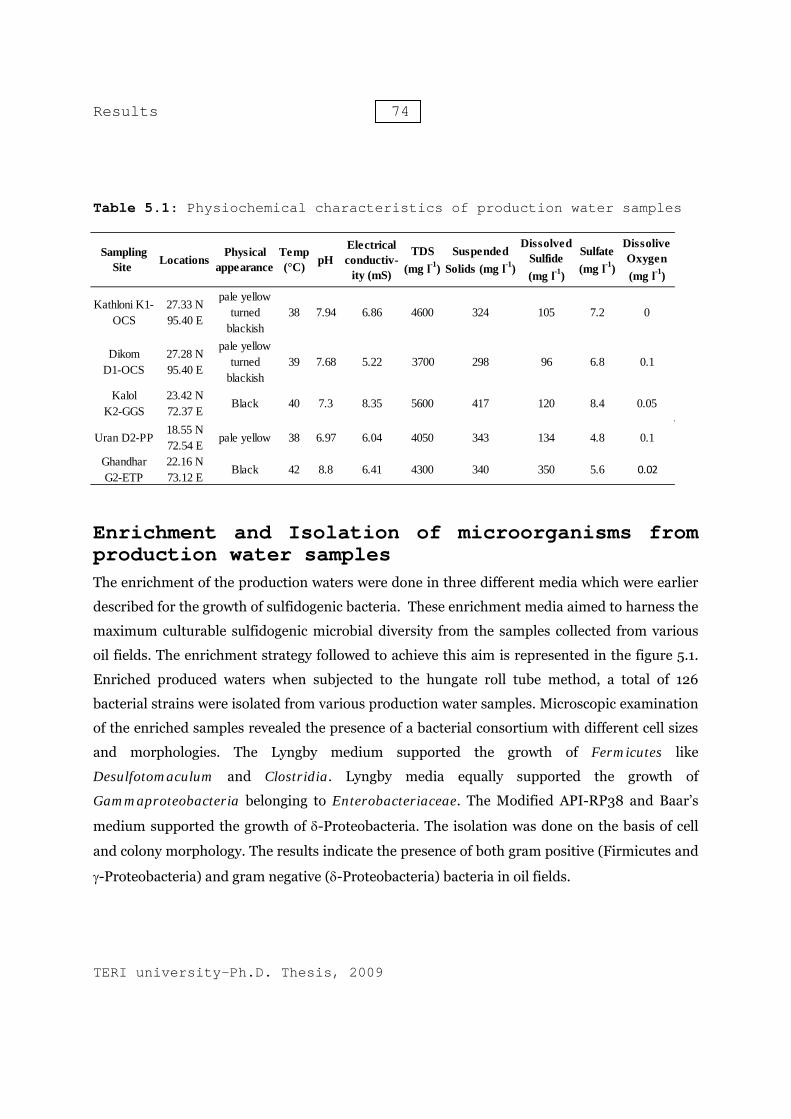
Results Bacterial diversity of production waters collected from various oil field sites Bacteria possess a unique capacity to adapt to diverse and intermittent environments. Bacterial strains existing in production water at phase-separator and storage tanks are capable to adapt to anoxic environment. These bacterial strains make use of short and long chain hydrocarbons, fatty acids and production chemicals as carbon and nitrogen sources. Sulfate, thiosulfate, sulfite and other sulfur compounds can be used by these bacteria as electron acceptors in the process of anaerobic respiration. The capacity of anaerobic respiration provides them ability to survive in anoxic environments. The activity to these microbes leads to production of hydrogen sulfide, which in turn is responsible for souring and corrosion of surface production facilities. Oil-water separators, water injection pump delivery system and storage tanks have higher amount of dissolved hydrogen sulfide and hence, were severely suffering from microbial induced corrosion. There is a need to study microbial diversity in such ecosystem, so to design efficient and specific methods to control their activity. The present study aims at analysing the diversity of the culturable sulfidogenic bacteria in the production water samples collected from various oil-water separation tanks. Culture independent molecular phylogenetic approach using functional bisulfite reductase (dsrB) was used to assess the non-culturable SRB populations. This was followed by development of quantitative real time PCR assay to quantify and monitor SRB population in production water. The surface oil production facilities installed in Dikom and Kathloni oil collection station (OCS), Kalol group gathering station (GGS), Ghandhar emulsion treatment plant (ETP) and Uran process plant (PP) have history of microbial souring and biocorrosion. These surface production facilities, actively treat crude oil, have problems of proper disposal of production water. Further as these facilities are suffering from microbial souring, disposal of production water is more complicated. The physiochemical characteristics of the production water sampled are listed in Table 5.1. 5
Transcript of 5 Results - shodhganga.inflibnet.ac.inshodhganga.inflibnet.ac.in/bitstream/10603/6303/13/13_chapter...

Results
Bacterial diversity of production waters collected from various oil field sites
Bacteria possess a unique capacity to adapt to diverse and intermittent environments. Bacterial strains existing in production water at phase-separator and storage tanks are capable to adapt to anoxic environment. These bacterial strains make use of short and long chain hydrocarbons, fatty acids and production chemicals as carbon and nitrogen sources. Sulfate, thiosulfate, sulfite and other sulfur compounds can be used by these bacteria as electron acceptors in the process of anaerobic respiration. The capacity of anaerobic respiration provides them ability to survive in anoxic environments. The activity to these microbes leads to production of hydrogen sulfide, which in turn is responsible for souring and corrosion of surface production facilities. Oil-water separators, water injection pump delivery system and storage tanks have higher amount of dissolved hydrogen sulfide and hence, were severely suffering from microbial induced corrosion. There is a need to study microbial diversity in such ecosystem, so to design efficient and specific methods to control their activity.
The present study aims at analysing the diversity of the culturable sulfidogenic bacteria in the production water samples collected from various oil-water separation tanks. Culture independent molecular phylogenetic approach using functional bisulfite reductase (dsrB) was used to assess the non-culturable SRB populations. This was followed by development of quantitative real time PCR assay to quantify and monitor SRB population in production water. The surface oil production facilities installed in Dikom and Kathloni oil collection station (OCS), Kalol group gathering station (GGS), Ghandhar emulsion treatment plant (ETP) and Uran process plant (PP) have history of microbial souring and biocorrosion. These surface production facilities, actively treat crude oil, have problems of proper disposal of production water. Further as these facilities are suffering from microbial souring, disposal of production water is more complicated. The physiochemical characteristics of the production water sampled are listed in Table 5.1.
5
Results 74
TERI university-Ph.D. Thesis, 2009
Table 5.1: Physiochemical characteristics of production water samples
Enrichment and Isolation of microorganisms from production water samples The enrichment of the production waters were done in three different media which were earlier
described for the growth of sulfidogenic bacteria. These enrichment media aimed to harness the
maximum culturable sulfidogenic microbial diversity from the samples collected from various
oil fields. The enrichment strategy followed to achieve this aim is represented in the figure 5.1.
Enriched produced waters when subjected to the hungate roll tube method, a total of 126
bacterial strains were isolated from various production water samples. Microscopic examination
of the enriched samples revealed the presence of a bacterial consortium with different cell sizes
and morphologies. The Lyngby medium supported the growth of Fermicutes like
Desulfotomaculum and Clostridia. Lyngby media equally supported the growth of
Gammaproteobacteria belonging to Enterobacteriaceae. The Modified API-RP38 and Baar’s
medium supported the growth of δ-Proteobacteria. The isolation was done on the basis of cell
and colony morphology. The results indicate the presence of both gram positive (Firmicutes and
γ-Proteobacteria) and gram negative (δ-Proteobacteria) bacteria in oil fields.
Kathloni K1-OCS
27.33 N 95.40 E
pale yellow turned
blackish38 7.94 6.86 4600 324 105 7.2 0
Dikom D1-OCS
27.28 N 95.40 E
pale yellow turned
blackish39 7.68 5.22 3700 298 96 6.8 0.1
Kalol K2-GGS
23.42 N 72.37 E
Black 40 7.3 8.35 5600 417 120 8.4 0.05
Uran D2-PP18.55 N 72.54 E
pale yellow 38 6.97 6.04 4050 343 134 4.8 0.1
Ghandhar G2-ETP
22.16 N 73.12 E
Black 42 8.8 6.41 4300 340 350 5.6 0.02
Dissolive Oxygen (mg l-1)
Sulfate (mg l-1)
LocationsTDS
(mg l-1)Suspended
Solids (mg l-1)
Dissolved Sulfide (mg l-1)
Sampling Site
Temp (°C) pH
Physical appearance
Electrical conductiv-
ity (mS)

Results 75
TERI university-Ph.D. Thesis, 2009
Figure 5.1: Enrichment Strategy for production waters
Diversity of culturable sulfidogenic bacteria from oil fields of India
The isolated strains from various production water samples were subjected for microbiological and molecular characterization. Cell morphology studies of these isolates by light microscopy and electron microscopy revealed a cell size of approximately 2-3 µm length. The cell size was linked with cell shapes like rods, cocci or vibrio. All isolates were able to produce hydrogen sulfide in the range of 55 to 250 mg.L-1. It was observed that hydrogen sulfide production was significantly higher in the microbes isolated from Lyngby medium. Most of the isolates have salinity tolerance range from 0 to 6% and optimal salinity between 0.5 to 2%. It was also noted that sulfide production by the isolates was decrease when they were grown at higher salinity. The optimum temperature ranges from 35-40°C and maximum growth temperature varies from 40-45°C.
Phylogenetic identification of the isolates by 16S rRNA gene indicated that Lyngby medium favoured the growth of several Firmicutes related to Clostridium thiosulfatireducens, Clostridium subterminale and Fusibacter paucivorans. Lyngby medium equally supported the growth of Enterobacteriaceae, similar to the genera Enterobacter, and Citrobacter. Modified API RP 38 and Baar’s (sulfate containing) medium favoured the growth of many spore forming

Results 76
TERI university-Ph.D. Thesis, 2009
and non spore forming SRB related to the genera Desulfotomaculum, Desulfomicrobium, Desulfobulbus, and Desulfovibrio.
To study the inter- and intra-species diversity between the bacterial strains originated from one geographical location the PCR-DGGE Profiling of 16S rRNA gene was done. Separation in DGGE is based on differences in sequence composition that affect the melting behaviour of the amplicon, causing a decrease in the electrophoretic mobility of a partially melted DNA molecule in a polyacrylamide gel containing a linearly increasing gradient of DNA denaturants. PCR amplification was carried out employing 63F (with 40bp GC clamp) and 518R universal primers resulted in the 495bp long amplicon. The PCR-DGGE of 16S rRNA amplicon of various isolated strain showed the different DGGE fingerprint in the gel. Position in the DGGE band is based on the GC percentage of the amplicons. The amplicons with the similar GC content were showing the bands at the same position. Therefore, the isolates to which these amplicons showing similar band position in DGGE were designated to one genotypic cluster. Cluster analysis was done by Bionumerics software (Applied mathematics, Belgium). Different genotypic clusters revealed the inter and intraspecies diversity among the isolates from the same geographical location. Previous researchers have also used PCR DGGE Profile to study inter- and intraspecies diversity among the isolated strains (Souza et al., 2004). PCR-DGGE method to analyse inter and intraspecies diversity between the pure isolates is as powerful technique as RAPD and RFLP (Bergsma-Vlami M et al., 2008).
Evaluation of diversity of culturable sulfidogenic bacteria in production water collected from oil-water separator located at Dikom Oil collection station Description of sample collection site and chemical characteristics of the collected sample Dikom oil fields (27.28 N 95.40 E) are owned by a public sector company Oil India Limited (OIL), Assam which is situated in the North eastern parts of India. The oil field is 20 km away from the Dibrugarh (city in Assam state of India). The ambient temperature of oil field ranged from 35- 40 oC. The company owns various oil collecting stations viz. Dikom, Kathloni, Bekulajaan, and Hathiali. The Dikom oil field are actively producing crude oil via number of oil wells located in field. The crude oil from these oil wells is transferred to Dikom Oil collection station where petroleum is separated from gas and production water. The production was pale in color which turned blackish in 2 min. The dissolved sulfide was measured 96 mg.L-1 and

Resul
TERI u
sulfate suspendcontenttanks pmesophproduct
MicroA total from Dnegativ2%. Thbetweengram-nfrom Lyand gra37°C (fmg.L-1. from Di
Figurepictur
ts
universit
content waded solids wt. The ambi
provides favhilic sulfidogtion facilitie
obiologof 22 bacte
Dikom (D1) ve short rodese isolatesn 80-100 m
negative rodyngby mediam negativefigure 5.2 a The morpikom produ
e 5.2: Lire of iso
ty-Ph.D.
as 6.8 mg.Lwere measuient tempervourable congenic bactees installed
gical crial isolates OCS. Bact
ds with salins were able mg.L-1 Singld, optimally ium enrichee rods were and 5.3). Hyphological auction water
ight microlate T18
Thesis,
L-1. Total diured 298 mgrature of oinditions forria lead to m in the field
haractes were isolaterial strainnity toleran to grow ople bacterial growing ated producti present whydrogen su
and physiocr has been l
roscopic 8
77
2009
issolved solg.L-1 revealel-water sepr the growtmicrobial so.
erizatiated from thns isolated nce range frptimally at isolate wat 2% salinityion water sahich were aulfide produchemical chisted in Tab
lids (TDS) ed the preseparator (39°th of mesopouring (incr
ion
he enrichme by usingrom 0.5-1037-40°C ans isolated fy and 35°Cample from
able to growuction in th
haracteristicble 5.2
Figure 5microscopT18.
were measence of high°C) and prophilic sulfidreased H2S
ents of prodg API RP38% and opti
nd H2S prodfrom Baar’s. Seventeen
m Dikom OCw optimally hese isolatecs of the ba
.3: Scannpic pictu
ured 3700 h organic anoduction wa
dogenic bact level) and c
duction wat8 medium imum saliniduction was medium. n isolates wCS. Both gra at salinity es ranges fracterial strai
ning elecure of is
mg.L-1 andnd inorganicater storageteria. Thesecorrosion of
ter collectedwere gramity was 1.5-s measuredIsolate wasere isolatedam positive0.5-2% and
rom 75-240ins isolated
ctron solate
d c e e f
d m
-d s d e d 0 d

Results 78
TERI university-Ph.D. Thesis, 2009
Table 5.2: Morphological physical and biochemical characteristics of bacterial strains isolated from Dikom production water samples
*values of H2S production after 28 days
Phylogenetic identification of the isolated bacterial strains from Dikom production water sample
16S rDNA sequencing of the isolates revealed that out of seventeen isolates from Lyngby medium out of which 12 strains T14, T18, T21, T59, T74, T96, T98, T107, T113, T142, T143 and T144,belongs to genera Clostridiaceae family and genus Clostridium. The clostridium strains were affiliated to 2 different species. The strains T14, T18, T21, T59, T74, T96, T98, T107 and T113 showing maximum similarity with Cloastridium subterminale (95%) and strains T142, T143 and T144 were affiliated to Clostridium thiosulfatireducens (96%). Four other strains T25, T63, T64 and T65 isolated from Lyngby medium were affiliated with Enterobacter cloacae (98%) . An isolate T114 showed 100% similarity with 16S rRNA sequence of Fusibacter paucivorans. The light and scanning electron microscopy photographs of isolate T18 is shown in
Isolate Number
Isolation Medium Cell Morphology Gram Staining
Salinity Tolerence
%
Optimal Salinity
%
Growth Temp °C
H2S production range mg L-1*
T141 Modified API RP 38 Short rod Gram negative 0.5-10 2 40 80
T150 Modified API RP 39 Short rod Gram negative 0.5-10 2 37 100
T151 Modified API RP 40 Short rod Gram negative 0.5-8 2 37 90
T152 Modified API RP 41 Short rod Gram negative 0.5-8 1.5 40 95
T60 Baar's Rod Gram negative 0-5 2 35 80T18 Lyngby Long Rod Gram positive 0-6 0.5 37 160T14 Lyngby Long Rod Gram positive 0-6 0.5 37 170T21 Lyngby Long Rod Gram positive 0-8 0.5 37 200T59 Lyngby Long Rod Gram positive 0-7 0.5 37 240T74 Lyngby Long Rod Gram positive 0-7 1 37 200T96 Lyngby Long Rod Gram positive 0-6 0.5 37 180T98 Lyngby Long Rod Gram positive 0-6 0.5 37 180T107 Lyngby Long Rod Gram positive 0-8 1.5 37 180T113 Lyngby Long Rod Gram positive 0-6 0.5 37 180T114 Lyngby Spindle shaped Rod Gram positive 0-10 0.5 37 75T142 Lyngby Long Rod Gram positive 0-6 0.5 37 220T143 Lyngby Long Rod Gram positive 0-7 0.5 37 240T144 Lyngby Long Rod Gram positive 0-8 0.5 37 240T25 Lyngby Long Rod Gram negative 0-6 0.5 37 150T63 Lyngby Long Rod Gram negative 0-5 0.5 37 180T64 Lyngby Long Rod Gram negative 0-6 0.5 37 120T65 Lyngby Long Rod Gram negative 0.5-5 0.5 37 130

Results 79
TERI university-Ph.D. Thesis, 2009
figure 5.2 and 5.3 respectively.All the 4 isolates T142, T150, T151 and T152, which were isolated by using API RP 38 medium, were affiliated to Desulfovibrio vulgaris (99%). Strain T60 was isolated from Baar’s medium which showed maximum similarity with Desulfomicrobium escumbium (99%). The phylogenetic affiliation of the sulfidogenic bacterial strains isolated from Dikom production water has been listed in Table 5.3
Table 5.3: Phylogenetic characterization of bacterial strains isolated
from Dikom production water samples
The phylogenetic tree of the isolated strains revealed the presence of both low and high G+C content organisms. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) are shown next to the branches. All positions containing gaps and missing data were eliminated from the dataset (Complete deletion option). There were a total of 418 nucleotide positions in the final dataset. The number of isolates from the same species are represented in a bracket next to the isolate name. The Phylogenetic tree of the isolates was distinctly divided in to 3 clades. Clade I consist of members of Clostridiaceae family (Gram positive and low G+C group), Clade II composed of members of
Isolate Number Family Phylogenetic affiliation (%similarity)T141 Desulfovibrionaceae Desulfovibrio vulgaris (99)T150 Desulfovibrionaceae Desulfovibrio vulgaris (99)T151 Desulfovibrionaceae Desulfovibrio vulgaris (99)T152 Desulfovibrionaceae Desulfovibrio vulgaris (99)T60 Desulfomicrobiaceae Desulfomicrobium escambium(99)T18 Clostridiaceae Clostridium subterminale (95)T14 Clostridiaceae Clostridium subterminale (95)T21 Clostridiaceae Clostridium subterminale (95)T59 Clostridiaceae Clostridium subterminale (95)T74 Clostridiaceae Clostridium subterminale (95)T96 Clostridiaceae Clostridium subterminale (96)T98 Clostridiaceae Clostridium subterminale (95)
T107 Clostridiaceae Clostridium subterminale (96)T113 Clostridiaceae Clostridium subterminale (95)T114 Peptostreptococcaceae Fusibacter paucivorans (100)T142 Clostridiaceae Clostridium thiosulfatireducens (96)T143 Clostridiaceae Clostridium thiosulfatireducens (96)T144 Clostridiaceae Clostridium thiosulfatireducens (96)T25 Enterobacteriaceae Enterobacter cloacae (98)T63 Enterobacteriaceae Enterobacter cloacae (98)T64 Enterobacteriaceae Enterobacter cloacae (98)T65 Enterobacteriaceae Enterobacter cloacae (98)

Results 80
TERI university-Ph.D. Thesis, 2009
Enterobacteriaceae. These are Gram negative bacteria with low G+C content. Clade III consist of members of delta-proteobacteria which are gram negative bacteria with high G+C content (figure 5.4).
Figure 5.4: Evolutionary relationships of 14 taxa (linearized). The evolutionary history was inferred using the Neighbor-Joining method. The phylogenetic tree was linearized assuming equal evolutionary rates in all lineages. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Maximum Composite Likelihood method and are in the units of the number of base substitutions per site. Phylogenetic analyses were conducted in MEGA 4
100
100
73
100
100
100
99
100
99
100
0.000.050.100.15
Clostridium subterminale (AF241842.1)
Clostridium botulium (M59087.1)
T143 (3)
Clostridium thiosulfatireducens (AB294141.1)
T60 (9)
T114 (1)
Fusibacter paucivorans (AF050099.1)
T25 (4)
Enterobactoer cloacae (EU048321.1)
Pantoea agglumerans (EF428997.1)
T60 (1)
Desulomicrobium escambium (AJ277886.1)
T141 (4)
Desulfovibrio vulgaris (AB294142.1)

Results 81
TERI university-Ph.D. Thesis, 2009
16S rDNA fingerprinting and cluster analysis of the isolated strains based on Denaturating Gradient Gel Electrophoresis
The DGGE analysis of the 16S amplified product of the pure strains generated a banding pattern based on the G+C content of the amplicon. As the Denaturating Gradient Gel Electrophoresis is efficient in distinction of amplicon differing in one base pair, strains belonging to same species also showed more than one banding patterns. In the cluster analysis the 22 strains isolated from the Dikom production water delineated into 12 genotypic cluster (figure 5.5).
Figure 5.5: Genotypic cluster analysis of the strains isolated from Dikom production water samples by Bray-Curtis clusters analysis. The 22 isolated strains were divided in 13 genotypes.
The 7 Isolates belonging to Clostridium subterminale were divided in to 5 clusters (VII, VIII, IX, X and XI). The largest cluster was genotypic cluster IX which had a total of 3 isolates (T59, T74
% Similarity
100
95908580757065605550454035
151
152
150
141
64
65
63
25
60
114
18
113
107
59
74
96
98
21
14
143
142
144
I
II
III
IV
VVIVII
VIII
IX
X
XI
XII
XIII

Results 82
TERI university-Ph.D. Thesis, 2009
and T96). The genotypic cluster XII and XIII consist of isolates belonging to the Clostridium thiosulfatireducens species. Isolates belonging to the Desulfovibrio vulgaris species delineated in to two genotypic clusters I and II, Both the cluster I and II were consist of 2 isolates T151/T152 and T150/T147. The genotypic cluster III and IV were consisting of isolates affiliated to the species Enterobacter cloacae. The isolates T64 and T65 were clustered in genotypic cluster III and T63 and T25 were clustered in genotypic cluster IV. The genotypic cluster V and VII were comprises of 1 isolate belonging to Fusibacter paucivorans and Desulfomicrobium escumbium respectively.
Evaluation of diversity of culturable sulfidogenic bacteria in production water collected from oil-water separator located at Kathloni Oil collection station
Description of sample collection site and chemical characteristics of the collected sample
Kathloni oil field (figure 5.6 and 5.7) is situated in the north-eastern region (27.33 N 95.40 E) of India in the state of Assam. Kathloni oil fields are also owned by Oil India Limited (OIL), Assam. The distance between Kathloni and Dikom oil fields is nearly 12 kms. These oil fields are actively producing crude oil via number of oil wells located in field. The crude oil from these oil wells is transferred to Kathloni Oil collection station (OCS) where crude petroleum is separated from gas and production water. The production water collected from oil-water separator was yellowish in colour which turned black after few minutes of collection.
Figure 5.6: Kathloni Oil collection station situated in Assam, India
Figure 5.7: Oil Separation tank installed at the Kathloni OCS, Assam, India

Results 83
TERI university-Ph.D. Thesis, 2009
The dissolved sulfide and sulfate in the production water was measured 105 and 7.2 mg.L-1 respectively. It was observed that dissolved sulfide content increased from 105 mg.L-1 at the oil-water separator tank to 210 mg.L-1 in the storage tank. No dissolved oxygen was detected in the sample. Total dissolved solids and suspended solid was measured 4600 and 324 mg.L-1 which was higher than the production water sample collected from the nearby Dikom OCS. The ambient temperature of oil-water separator (38°C) and production water storage tanks provides favourable conditions for the growth of mesophilic sulfidogenic bacteria. Growth of these sulfidogenic bacteria led to microbial souring (increased H2S level), corrosion of production facilities and environmental hazards from production water spills.
Microbiological characterization
A total number of 43 bacterial strains were isolated from Kathloni production waters. Five Bacterial strains isolated u API RP38 medium were gram negative short rods with salinity tolerance range from 0.5-10% and optimum salinity was 2%. These isolates were able to grow optimally at 37-40°C and H2S production was measured between 80-120 mg.L-1. The light and scanning electron micrograph of isolate T147 is represented in figure 5.8 and 5.9. The other two isolates (T145 and T146), isolated from API RP38 medium had small curved rod morphology. These isolates were optimally growing at 2% salinity and 30 °C temperature. Hydrogen sulfide production from T145 and T146 was 55 and 90 mg.L-1.
Isolates from Baar’s medium were gram-positive short curved rods. All these isolates were able to grow at salinity between 0-14%, however optimal salinity was 6%. These isolates were gram-positive rods having optimal growth temperature of 30 and H2S production range from 210-225. Thirty-one isolates were isolated from Lyngby medium enriched production water sample from Kathloni OCS. Both gram positive and gram negative rods were present which were able to
Figure 5.8: Light microscopic pictures of isolate T147
Figure 5.9: Scanning electron microscopic pictures of isolate T147

Results 84
TERI university-Ph.D. Thesis, 2009
grow optimally at salinity 0.5-1% and temperature between 35-40°C. Morphology of these isolates was short, long or think rods. Hydrogen sulfide production in these isolates varies between 55-240 mg.L-1. The morphology and growth characteristics of the isolates are given in table 5.4
Phylogenetic identification of the isolated bacterial strains from Kathloni production water sample
The bacterial strains were isolated from Lyngbay medium which were belonging to 4 phylotypes. These phylotypes were affiliated to Clostridium subterminale, Clostridium thiosulfatireducens, Citrobacter freundii and Enterobacter cloacae. The bacterial strains were isolated from enriched production water in API RP 38 medium which were affiliated to Desulfovibrio vulgaris and Desulfobulbus sp. Bacterial strains isolated from production water enriched in Baar’s medium showed maximum similarity with the 16S rRNA gene of Desulfotomaculum halophilum. Earlier Desulfotomaculum halophilum has only been reported from oil field brine in France. These were gram-positive rods and can tolerate up to 14% NaCl (Tardy-Jacquenod et al., 1998).
Clostridium thiosulfatireducens was also found which is close relative of C. subterminale and shares the similar phenotypic properties including the use of proteins, peptides and amino acids. Micorbiological characterization of the isolated strains and their phylogenetic affiliation with the closest relative are shown in table 5.5
The evolutionary phylogenetic relationship of the bacterial strains isolated from Kathloni production water sample is shown in figure 5.10. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) are shown next to the branches. All positions containing gaps and missing data were eliminated from the dataset (Complete deletion option).
There were a total of 416 nucleotide positions in the final dataset. The representatives of the isolated bacteria are represented in the phylogenetic tree. The number in the bracket represents total number of bacterial isolates affiliated to same species. The phylogenetic tree was divided in 3 groups. Group I represented the bacterial isolates which were affiliated to Gram negative and low G+C ratio. These bacterial isolates were belonging to Enterobacteriaceae family. Group II represented bacterial isolates belonging to members of delta-proteobacteria division. The isolates affiliated to genus Desulfovibrio and Desulfobulbus were belonging to this group. These isolates were gram-negative and high G+C content. Group III was comprised of members

Results 85
TERI university-Ph.D. Thesis, 2009
belonging to Clostridiaceae and Peptococcaceae family. Members of Group III were Gram positive having low G+C content.
Table 5.4: Morphological physical and biochemical characteristics of bacterial strains isolated from Kathloni production water samples
Isolate Number
Isolation Medium Cell Morphology Gram Staining
Salinity Tolerence
%
Optimal Salinity
%
Growth Temp °C
H2S production range mg L-1*
T147 Modified API RP 38 Vibrio Gram negative 0.5-10 2 40 80
T165 Modified API RP 38 Vibrio Gram negative 0.5-10 2 37 120
T25 Modified API RP 38 Vibrio Gram negative 0.5-08 2 37 120
T61 Modified API RP 38 Vibrio Gram negative 0.5-08 2 37 110
T62 Modified API RP 38 Vibrio Gram negative 0.5-08 2 40 80
T29 Baar's Curved rod Gram positive 0-14 6 30 210T31 Baar's Curved rod Gram positive 0-14 6 30 210T83 Baar's Curved rod Gram positive 0-14 6 30 225T88 Baar's Curved rod Gram positive 0-14 6 30 220T93 Baar's Curved rod Gram positive 0-14 6 30 220
T145 Modified API RP 38 Small Curved Rod Gram negative 0.5-3 2 30 55
T146 Modified API RP 38 Small Curved Rod Gram negative 0.5-3 2 30 90
T23 Lyngby Spore forming Rod Gram positive 0-6 0.5 37 250T58 Lyngby Spore forming Rod Gram positive 0-6 0.5 37 250T10 Lyngby Spore forming Rod Gram positive 0-6 0.5 37 230T11 Lyngby Spore forming Rod Gram positive 0-6 0.5 37 250T16 Lyngby Spore forming Rod Gram positive 0-6 0.5 37 200T8 Lyngby Spore forming Rod Gram positive 0-6 0.5 37 150T9 Lyngby Spore forming Rod Gram positive 0-6 0.5 37 170T63 Lyngby Spore forming Rod Gram positive 0-6 0.5 37 230T24 Lyngby Spore forming Rod Gram positive 0-6 0.5 37 230T46 Lyngby Spore forming Rod Gram positive 0-6 0.5 37 240T12 Lyngby Spore forming Rod Gram positive 0-6 0.5 35 150T82 Lyngby Spore forming Rod Gram positive 0-6 0.5 35 180T89 Lyngby Spore forming Rod Gram positive 0-6 0.5 35 180T90 Lyngby Thick Rod Gram negative 0.5-2 0.5 40 55T91 Lyngby Thick Rod Gram negative 0.5-2 0.5 40 120T94 Lyngby Thick Rod Gram negative 0.5-2 0.5 40 130T95 Lyngby Thick Rod Gram negative 0.5-2 0.5 40 130T97 Lyngby Thick Rod Gram negative 0.5-2 1 40 90
T100 Lyngby Long Rod Gram negative 0.5-2.5 0.5 37 150T103 Lyngby Long Rod Gram negative 0.5-2.5 0.5 37 160T104 Lyngby Long Rod Gram negative 0.5-2.5 0.5 37 160T105 Lyngby Long Rod Gram negative 0.5-2.5 0.5 37 190T109 Lyngby Long Rod Gram negative 0.5-2.5 0.5 37 150T111 Lyngby Long Rod Gram negative 0.5-2.5 0.5 37 200T115 Lyngby Spore forming Rods Gram positive 0-6 0.5 37 220T117 Lyngby Spore forming Rods Gram positive 0-5 0.5 37 170T140 Lyngby Spore forming Rods Gram positive 0-5 0.5 35 170T141 Lyngby Spore forming Rods Gram positive 0-5 1 37 210T118 Lyngby Spore forming Rods Gram positive 0-6 1 37 215T119 Lyngby Spore forming Rods Gram positive 0-4 0.5 37-40 170T120 Lyngby Spore forming Rods Gram positive 0-6 0.5 37-40 170

Results 86
TERI university-Ph.D. Thesis, 2009
*values of H2S production after 28 days
Table 5.5: Phylogenetic characterization of the strains isolated from Kathloni production water samples
Isolate Number Family Phylogenetic affiliation (%similarity)T147 Desulfovibrionaceae Desulfovibrio vulgaris (99)T165 Desulfovibrionaceae Desulfovibrio vulgaris (99)T25 Desulfovibrionaceae Desulfovibrio vulgaris (99)T61 Desulfovibrionaceae Desulfovibrio vulgaris (99)T62 Desulfovibrionaceae Desulfovibrio vulgaris (99)T29 Peptococcaceae Desulfotomaculum halophilum (99)T31 Peptococcaceae Desulfotomaculum halophilum (99)T83 Peptococcaceae Desulfotomaculum halophilum (99)T88 Peptococcaceae Desulfotomaculum halophilum (99)T93 Peptococcaceae Desulfotomaculum halophilum (99)
T145 Desulfobulbaceae Desulfobulbus species (99)T146 Desulfobulbaceae Desulfobulbus species (99)T23 Clostridiaceae Clostridium subterminale (95)T58 Clostridiaceae Clostridium subterminale (95)T10 Clostridiaceae Clostridium subterminale (95)T11 Clostridiaceae Clostridium subterminale (95)T16 Clostridiaceae Clostridium subterminale (95)T8 Clostridiaceae Clostridium subterminale (95)T9 Clostridiaceae Clostridium subterminale (95)T63 Clostridiaceae Clostridium subterminale (95)T24 Clostridiaceae Clostridium subterminale (95)T46 Clostridiaceae Clostridium subterminale (95)T12 Clostridiaceae Clostridium subterminale (95)T82 Clostridiaceae Clostridium subterminale (95)T89 Clostridiaceae Clostridium subterminale (95)T90 Enterobacteriaceae Citrobacter freundii (98)T91 Enterobacteriaceae Citrobacter freundii (98)T94 Enterobacteriaceae Citrobacter freundii (98)T95 Enterobacteriaceae Citrobacter freundii (98)T97 Enterobacteriaceae Citrobacter freundii (98)
T100 Enterobacteriaceae Enterobacter cloacae (98)T103 Enterobacteriaceae Enterobacter cloacae (98)T104 Enterobacteriaceae Enterobacter cloacae (98)T105 Enterobacteriaceae Enterobacter cloacae (98)T109 Enterobacteriaceae Enterobacter cloacae (98)T111 Enterobacteriaceae Enterobacter cloacae (98)T115 Clostridiaceae Clostridium thiosulfatireducens (96)T117 Clostridiaceae Clostridium thiosulfatireducens (96)T140 Clostridiaceae Clostridium thiosulfatireducens (96)T141 Clostridiaceae Clostridium thiosulfatireducens (96)T118 Clostridiaceae Clostridium thiosulfatireducens (96)T119 Clostridiaceae Clostridium thiosulfatireducens (96)T120 Clostridiaceae Clostridium thiosulfatireducens (96)

Results 87
TERI university-Ph.D. Thesis, 2009
.
Figure 5.10: Evolutionary relationships of 16 taxa (linearized) The evolutionary history was inferred using the Neighbor-Joining method. The phylogenetic tree was linearized assuming equal evolutionary rates in all lineages. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Maximum Composite Likelihood method and are in the units of the number of base substitutions per site. Phylogenetic analyses were conducted in MEGA4
16S rDNA fingerprinting and cluster analysis of the isolated strains based on Denaturating Gradient Gel Electrophoresis
The 16S rDNA gene fingerprint of the pure strains isolated from the Kathloni production water revealed that isolated strains belonging to different species showed different banding pattern. It was also noticed that isolates belonging to same species also have different pattern, suggesting intraspecies diversity between the isolates of same species (figure 5.11). Cluster analysis of the
100
100
100
99
100
100
51
100
95
100
0.000.050.100.15
Pantoea agglumerans (EF428997.1)
Citrobacter freundii (AB244454.1)
T100 (6)
T90 (5)
Enterobactoer cloacae (EU048321.1)T147 (5)
Desulfovibrio vulgaris (AB294142.1)
T145 (2)Desulfobulbus sp. (AY548775.1)
T29 (5)Desulfotomaculum halophilum (U88891.1)
T115 (7)
Clostridium thiosulfatireducens (AB294141.1)T23 (13)
Clostridium subterminale (AF241842.1)
Clostridium botulium (M59087.1)
Results 88
TERI university-Ph.D. Thesis, 2009
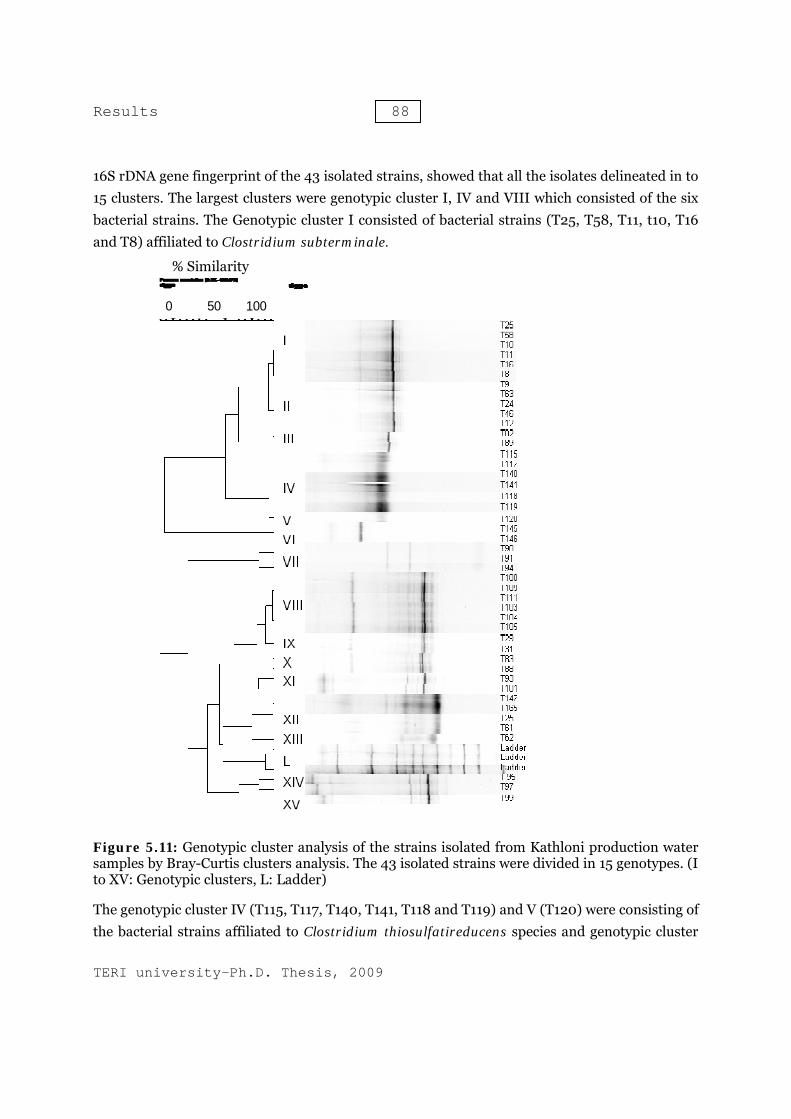
16S rDNA gene fingerprint of the 43 isolated strains, showed that all the isolates delineated in to 15 clusters. The largest clusters were genotypic cluster I, IV and VIII which consisted of the six bacterial strains. The Genotypic cluster I consisted of bacterial strains (T25, T58, T11, t10, T16 and T8) affiliated to Clostridium subterminale.
Figure 5.11: Genotypic cluster analysis of the strains isolated from Kathloni production water samples by Bray-Curtis clusters analysis. The 43 isolated strains were divided in 15 genotypes. (I to XV: Genotypic clusters, L: Ladder)
The genotypic cluster IV (T115, T117, T140, T141, T118 and T119) and V (T120) were consisting of the bacterial strains affiliated to Clostridium thiosulfatireducens species and genotypic cluster
% Similarity
0 50 100

Results 89
TERI university-Ph.D. Thesis, 2009
VIII consisted of the bacterial strains T100, T109, T111, T103, T104 and T105, which showed highest similarity with Enterobacter cloacae species. The genotypic cluster II was made by the bacterial strains T9, T63, T24, T46 and T12, which were also affiliated to Clostridium subterminale. The genotypic cluster VI consisted of both the bacterial strains (T145 and T146) which were showed maximum similarity with Desulfobulbus species. The isolated bacterial strains T90, T91 and T94 set down in genotypic cluster VII. These strains were affiliated to the Enterobacteriaceae family. were composed of two bacterial isolates each. The bacterial strains T29/T31, T83/T88 and T93/T101 were belonging to the genotypic clusters IX, X and XI respectively. The bacterial strains T145, T165, T22 and T61 which were affiliated to the Desulfovibrio vulgaris species, grouped in the cluster XII.
Evaluation of diversity of culturable sulfidogenic bacteria in production water collected from oil-water separator located at Kalol group gathering station (GGS)
Description of sample collection site and chemical characteristics of the collected sample
Kalol oil field is situated in the western region (23.42 N 72.37 E) of India in the state of Gujarat. The Kalol oil fields is managed by Oil and Natural Gas Commission Ltd (ONGC), a public sector undertaking of Government of India. Kalol oil fields are actively producing crude oil via number of oil wells located in field. The crude oil from all these oil wells is transferred to Kalol GGS where petroleum oil is separated from gas and production water. The production water collected from separator tank was translucent in appearance. The dissolved sulfide and sulfate content in the production water was 120 and 8.4 mg.L-1 respectively. Total dissolved solids (5600 mg.L-1) and suspended solids (417 mg.L-1) were highest among the investigated sites. The ambient temperature of oil-water separator (40°C) and production water storage tanks provides favourable conditions for the growth of mesophilic sulfidogenic bacteria. Microbiological characterization Bacterial strains isolated from Kalol production water sample were gram-negative short and gram-positive long and thick rods in morphology. Of the 21 bacterial strains, 7 were isolated from API-RP38 medium. Bacterial isolate T133 was optimally growing at salinity 2.5% while other isolates (T163, T167, T80, T125, T126 and T132) had optimal salinity of 2%. These isolates were able to tolerate salinity range from 0-10 %. Isolate T163 and T167 had optimal growing temperature of 35 °C while other isolates (T80, T125, T126 and T132) were growing optimally at

Results 90
TERI university-Ph.D. Thesis, 2009
40 °C. The H2S production by the isolates was nearly equal and varies between 120-160 mg.L-1. However, H2S production by T80 was 80 mg.L-1. The bacterial strains (T122, T134, T135 and T172) isolated from Baar’s medium were small curved rods which were optimally growing at 2% NaCl concentration and 30 °C temperature. All the isolates were able to tolerate 0.5-3% salinity and 40 °C temperature. The H2S production by the isolates was unequal and varies from 55 to 160 mgL-1.
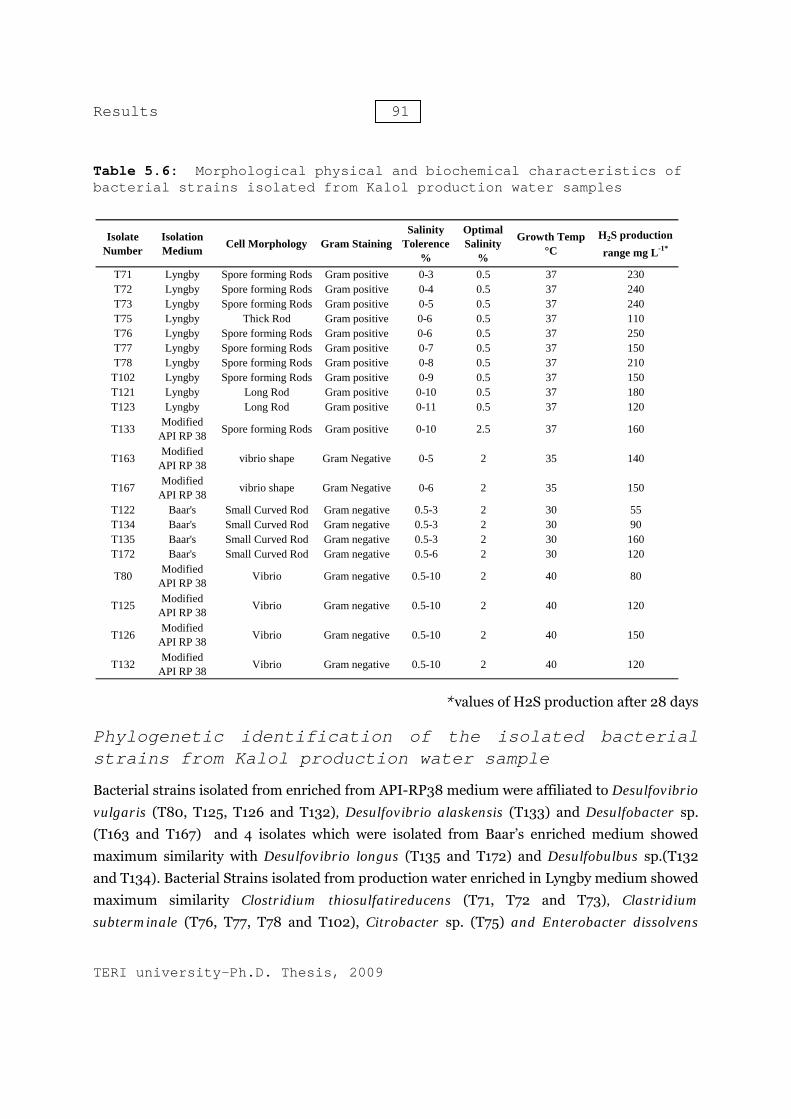
Bacterial strains (T71, T72, T73, T75, T76, T77, T78, T102, T121 and T12) isolated from Kalol production water, enriched in Lyngby medium, revealed the presence of gram positive thick or long Rods. The optimum growth of all the strains was obtained at the salinity 0.5% salinity and 37 °C temperature. The H2S production was estimated between 110 - 240 mg.L-1. All these isolates were able to tolerate salinity between 0-3% and maximum growth temperature was 45 °C except bacterial strains T121 and 123, which did not show any growth beyond 37 °C. However, in addition to normal rod shaped morphology, the cells of strain T76, T77, T78 and T102 showed the presence of long flexuous rods of variable length with some going up to 100μm at 45˚C. The light microscopy photographs and scanning electron micrographs of these cells are shown in Figure 5.12 and 5.13. The morphology and growth characteristics of the bacterial isolates from Kalol oil field’s production water are mentioned in Table 5.6.
Figure:5.13: Scanning electron microscopic pictures of isolate T102
Figure 5.12: Light microscopic pictures of isolate T102
Results 91
TERI university-Ph.D. Thesis, 2009
Table 5.6: Morphological physical and biochemical characteristics of bacterial strains isolated from Kalol production water samples
*values of H2S production after 28 days
Phylogenetic identification of the isolated bacterial strains from Kalol production water sample
Bacterial strains isolated from enriched from API-RP38 medium were affiliated to Desulfovibrio vulgaris (T80, T125, T126 and T132), Desulfovibrio alaskensis (T133) and Desulfobacter sp. (T163 and T167) and 4 isolates which were isolated from Baar’s enriched medium showed maximum similarity with Desulfovibrio longus (T135 and T172) and Desulfobulbus sp.(T132 and T134). Bacterial Strains isolated from production water enriched in Lyngby medium showed maximum similarity Clostridium thiosulfatireducens (T71, T72 and T73), Clastridium subterminale (T76, T77, T78 and T102), Citrobacter sp. (T75) and Enterobacter dissolvens
Isolate Number
Isolation Medium Cell Morphology Gram Staining
Salinity Tolerence
%
Optimal Salinity
%
Growth Temp °C
H2S production range mg L-1*
T71 Lyngby Spore forming Rods Gram positive 0-3 0.5 37 230T72 Lyngby Spore forming Rods Gram positive 0-4 0.5 37 240T73 Lyngby Spore forming Rods Gram positive 0-5 0.5 37 240T75 Lyngby Thick Rod Gram positive 0-6 0.5 37 110T76 Lyngby Spore forming Rods Gram positive 0-6 0.5 37 250T77 Lyngby Spore forming Rods Gram positive 0-7 0.5 37 150T78 Lyngby Spore forming Rods Gram positive 0-8 0.5 37 210T102 Lyngby Spore forming Rods Gram positive 0-9 0.5 37 150T121 Lyngby Long Rod Gram positive 0-10 0.5 37 180T123 Lyngby Long Rod Gram positive 0-11 0.5 37 120
T133 Modified API RP 38 Spore forming Rods Gram positive 0-10 2.5 37 160
T163 Modified API RP 38 vibrio shape Gram Negative 0-5 2 35 140
T167 Modified API RP 38 vibrio shape Gram Negative 0-6 2 35 150
T122 Baar's Small Curved Rod Gram negative 0.5-3 2 30 55T134 Baar's Small Curved Rod Gram negative 0.5-3 2 30 90T135 Baar's Small Curved Rod Gram negative 0.5-3 2 30 160T172 Baar's Small Curved Rod Gram negative 0.5-6 2 30 120
T80 Modified API RP 38 Vibrio Gram negative 0.5-10 2 40 80
T125 Modified API RP 38 Vibrio Gram negative 0.5-10 2 40 120
T126 Modified API RP 38 Vibrio Gram negative 0.5-10 2 40 150
T132 Modified API RP 38 Vibrio Gram negative 0.5-10 2 40 120

Results 92
TERI university-Ph.D. Thesis, 2009
(T121 and T123). The microbiological characterization of the bacterial isolates from Kalol production water and their phylogenetic affiliation is represented in table 5.7.
Table 5.7: Phylogenetic characterization of the strains isolated from Kalol production waters
The phylogenetic analysis was conducted by MEGA4 revealed that all the isolates distributed between the four major phylogenetic clades. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) are shown next to the branches. The phylogenetic tree was linearized assuming equal evolutionary rates in all lineages. All positions containing gaps and missing data were eliminated from the dataset (Complete deletion option). There were a total of 403 nucleotide positions in the final dataset. The clade I consist of members of delta-proteobacteria division. This clade consists of sulfate reducing bacteria belonging to 5 different genera. This revealed the presence of overall high diversity of culturable bacteria belonging to delta-proteobacteria. The Clade 2 composed of members of Clostridiaceae family. Clade III consisted of the isolate belonging to Enterobacter and Clade IV was composed of bacterial isolates belonging to Genera Citrobacter freundii (figure 5.14)
Isolate Number Family Phylogenetic affiliation (%similarity)T71 Clostridiaceae Clostridium thiosulfatireducens (96)T72 Clostridiaceae Clostridium thiosulfatireducens (97)T73 Clostridiaceae Clostridium thiosulfatireducens (97)T75 Enterobacteriaceae Citrobacter sp. (98)T76 Clostridiaceae Clostridium subterminale (95)T77 Clostridiaceae Clostridium subterminale (95)T78 Clostridiaceae Clostridium subterminale (96)
T102 Clostridiaceae Clostridium subterminale (95)T121 Enterobacteriaceae Enterobacter dissolvens (98)T123 Enterobacteriaceae Enterobacter dissolvens (98)T133 Desulfovibrionaceae Desulfovibrio alaskensis (98)T163 Desulfobacteraceae Desulfobacter sp. (98)T167 Desulfobacteraceae Desulfobacter sp. (98)T122 Desulfobulbaceae Desulfobulbus species (99)T134 Desulfobulbaceae Desulfobulbus species (99)T135 Desulfovibrionaceae Desulfovibrio longus (99)T172 Desulfovibrionaceae Desulfovibrio longus (99)T80 Desulfovibrionaceae Desulfovibrio vulgaris (99)
T125 Desulfovibrionaceae Desulfovibrio vulgaris (99)T126 Desulfovibrionaceae Desulfovibrio vulgaris (99)T132 Desulfovibrionaceae Desulfovibrio vulgaris (99)

Results 93
TERI university-Ph.D. Thesis, 2009
Figure 5.14: Evolutionary relationships of 21 taxa (linearized) The evolutionary history was inferred using the Neighbor-Joining method . The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Maximum Composite Likelihood method and are in the units of the number of base substitutions per site. Phylogenetic analyses were conducted in MEGA4
83
100
99
100
99
54
99
100
100
100
100
97
68
100
63
98
73
83
0.00.10.20.30.40.5
Desulfovibrio Longus (AY359867.1)
T135 (2)
T80 (4)Desulfovibrio vulgaris (AB294142.1)
Desulfovibrio alaskensis (DQ867001.1)
T133 (1)T122 (2)Desulfobulbus sp. (AY548775.1)
T163 (2)
Desulfobacter postgatei (AF 418180.1)
Desulfobacter curvatus (AF418175.1)
T71 (3)
Clostridium thiosulfatireducens (AB294141.1)
T76 (4)
Clostridium subterminale (AF241842.1)Clostridium botulium (M59087.1)
Enterobactoer cloacae (EU048321.1)T121 (2)
Pantoea agglumerans (EF428997.1)
Citrobacter freundii (AB244454.1)
T75 (1)

Resul
TERI u
16S isolaElec
The 16Srevealed(figure patternof the 1in to 13clustersT132 we
FigureKalol 21 isGenoty
The geEnterobT102, w
ts
universit
rDNA fated strophor
S rDNA gend that isola5.15). It w, suggesting
16S rDNA g3 clusters. s. The isolaere belongin
e 5.15: producti
solated ypic clus
enotypic clubacter dissowhich were
% Similarity
ty-Ph.D.
fingerpstrains resis
ne fingerprated strains
was also notg intraspeci
gene fingerpStrains affi
ates T125 anng to cluste
Genotypiion watestrains sters and
uster III colvens of fae showing
Thesis,
printinbased
rint of the s belonging ticed that iies diversity
print of the iliated to thnd T126 we
er XI and XI
c cluster samplewere div
d V: ladd
consisted oamily Enter similarity
94
2009
g and d on D
pure strain to differenisolates bely between t isolated str
he Desulfovere groupedII respective
er analyses by Bravided inder, C: T
of bacteriaobacteriace with Clos
clusteDenatur
ns isolated fnt species slonging to the isolates rains, showibrio vulgad in genotyely.
sis of thay-Curtisn 15 gen123 samp
l strain Teae. The bastridium su
er analating
from the Kshowed diffsame speci of same sp
wed that all aris were cluypic cluster
he strais clustenotypes le was ag
121 and Tacterial isolaubterminale
lysis Gradien
Kalol produferent bandies also havpecies. Clusthe isolatesustered in 3 I and strai
ns isolars analy(I-IV angain load
T123 affiliaates T76, T7e grouped
of thent Gel
ction waterding patternve differentter analysis
s delineated3 genotypicin T80 and
ated fromysis. Thend V-XV:ded)
ated to the77, T78 and in to two
e l
r n t s d c d
m e
e d o

Results 95
TERI university-Ph.D. Thesis, 2009
genotypic clusters IV and V. The genotypic cluster IX (T72 and T73) and X (T71) were consisting of the bacterial strains affiliated to Clostridium thiosulfatireducens species. These results revealed the diversity of 16S rDNA gene between the isolates within the species of Desulfovibrio vulgaris, Clostridium subterminale and Clostridium thiosulfatireducens.
Evaluation of diversity of culturable sulfidogenic bacteria in production water collected from oil-water separator located at Uran process plant
Description of sample collection site and chemical characteristics of the collected sample
Uran process plant (18.55N, 72.54E) is located on the coastline of Mumbai (Bombay). The Mumbai High Fields to Uran Trunkline extend starts from Mumbai High Fields and ends at Uran Process plant. Oil & Natural Gas Corporation Ltd has constructed two new pipeline connections from the Mumbai High Fields to Uran over a total length of 204 kilometres. These new pipelines have been constructed in order to replace the existing Bombay High Fields to Uran Trunkline. The old pipelines was ineffective due to exposure to biocorrosion and biofouling for longer duration. The crude petroleum oil produced from offshore oil reservoirs of the Bombay high is transferred to the Uran process plant for further separation of water and gas from the crude oil. Uran process plant and trunkline are suffering from microbial souring and microbial induced corrosion due to activities of sulfidogenic bacteria. The crude petroleum oil produced from offshore oil reservoirs of the Bombay high is transferred to the Uran process plant for further separation of water and gas from the crude oil. Uran process plant and flow line are suffering from microbial souring and microbial induced corrosion due to activities of sulfidogenic bacteria. The production water samples collected from Uran process plant was pale in colour and dissolved sulfide and sulfate was 134 mg.L-1 and 4.8 mg.L-1 respectively. The Uranproduction water had black particles which settled down after a short time. The total dissolved solids and suspended solid were 4050 and 343 mg.L-1. Dissolved oxygen was 0.1 mg.L-
1. The temperature of separation tank located at uran process plant was 38 °C.
Microbiological Characterization
A total number of 28 bacterial strains were isolated from enriched production water collected from Uran process plant. API RP838 and Baar’s medium favoured the growth of short rods and gram negative bacteria. Of the six strains isolated from API RP38 medium, 3 isolates (T136,

Results 96
TERI university-Ph.D. Thesis, 2009
T148, and T149) were growing optimally at salinity 2% and 35 °C while other 3 bacterial isolates (T154, T155 and T156) were growing optimally at salinity 0.5% and 37-40°C. These strains showed growth till 45°C.
Table 5.8: Morphological physical and biochemical characteristics of bacterial strains isolated from Uran production water samples
*values of H2S production after 28 days
The H2S production by these strains was 80-160 mg.L-1. Five bacterial strains (T 159, T160, T161, T162 and T164) were isolated from the production water enriched with Baar’s medium. All these isolates were growing optimally at 2% salinity and 35 °C. The H2S production by these
Isolate Number
Isolation Medium Cell Morphology Gram Staining
Salinity Tolerence
%
Optimal Salinity
%
Growth Temp °C
H2S production range mg L-1*
T106 Lyngby Long Rod Gram positive 0-6 0.5 37 150T110 Lyngby Long Rod Gram positive 0-6 0.5 37 210T112 Lyngby Long Rod Gram positive 0-6 0.5 37 220T116 Lyngby Long Rod Gram positive 0-6 0.5 37 240T124 Lyngby Thick Rod Gram positive 0-6 0.5 37 55T127 Lyngby Thick Rod Gram positive 0-6 0.5 37 200T128 Lyngby Thick Rod Gram positive 0-6 0.5 37 130T129 Lyngby Thick Rod Gram positive 0-6 0.5 37 130T130 Lyngby Thick Rod Gram positive 0-6 0.5 37 150T131 Lyngby Long Rod Gram positive 0-6 0.5 37 180T137 Lyngby Long Rod Gram positive 0-6 0.5 37 150T138 Lyngby Long Rod Gram positive 0-6 0.5 37 120T139 Lyngby Long Rod Gram positive 0-6 0.5 37 140T180 Lyngby Long Rod Gram positive 0-6 0.5 37 140T181 Lyngby Long Rod Gram positive 0-6 0.5 37 150T182 Lyngby Long Rod Gram positive 0-6 0.5 37 250T183 Lyngby Long Rod Gram positive 0-6 0.5 37 210T159 Baar's Rod Gram negative 0-5 2 35 80T160 Baar's Rod Gram negative 0-5 2 35 210T161 Baar's Rod Gram negative 0-5 2 35 150T162 Baar's Rod Gram negative 0-5 2 35 120T164 Baar's Rod Gram negative 0-5 2 35 140
T136 Modified API RP 38 Short rod Gram negative 0-5 2 35 120
T148 Modified API RP 38 Short rod Gram negative 0-5 2 35 160
T149 Modified API RP 38 Short rod Gram negative 0-5 2 35 130
T154 Modified API RP 38 Short rod Gram negative 0.5-10 2 40 80
T155 Modified API RP 38 Short rod Gram negative 0.5-10 0.5 37 120
T156 Modified API RP 38 Short rod Gram negative 0.5-10 0.5 37 130

Results 97
TERI university-Ph.D. Thesis, 2009
strains was notably different and ranges from 80-210 mg.L-1. The detailed morphological and growth characteristics of the isolates is given in table 5.8. The Lyngby medium supported the growth of 17 bacterial strains which were gram-positive rods. These isolates were growing optimally at 0.5% salinity and 37 °C. These isolates were able to tolerate salinity up to 6% and highest temperature of 45°C. The H2S production by these strains was between 120-250 mg.L-1 except isolate T124, which showed lower H2S production (50 mg.L-1). The morphology of isolate T182 was gram-positive long rod, as shown in figure 5.16 and 5.17.
Phylogenetic identification of the isolated bacterial strains from Uran production water sample
Isolates belonging to family Desulfovibrionaceae and Desulfomicrobiaceae were isolated in production water sample enriched in API-RP38 and Baar’s medium respectively. These isolates were showing maximum similarity with the 16S rRNA gene of Desulfovibrio longus (T136, T148 and T149), Desulfovibrio vulgaris (T154, T155 and T156) and Desulfomicrobium escambium (T159, T160, T161, T162 and T164). Lyngby medium supported the growth of members of family Clostrideace and Enterobacteacece , with the strains affiliated to Clostridium thiosulfatireducens (T181, T182 and T183), Clastridium subterminale (T106, T110, T112 and T116), Citrobacter freundii (T124, T127, T128, T129 and T130) and Enterobacter dissolvens (T131, T137, T138, T139 and T180). Microbiological and phylogenetic characterization of the bacterial strains, isolated from Uran production water samples is represented in table 5.9.
Figure 5.16: Light microscopic and pictures of isolate T182
Figure 5.17: Scanning electron microscopic pictures of isolate T182

Results 98
TERI university-Ph.D. Thesis, 2009
Table 5.9: Phylogenetic characterization of the isolated bacterial strains from Uran production water sample
Phylogenetic tree analysis of the bacterial isolates from Kalol production water samples revealed the presence of 3 major phylogenetic clades (figure 5. 18). Clade I consisted of cultures affiliated to the genera Citrobacter and Enterobacter. These cultures were gram negative and with low GC content. Clade II composed of the isolates affiliated to Clostridiaceae family. Clade III consisted of the cultures belonging to gram-negative and high G+C group of delta-proteobacteria.
Isolate Number Family Phylogenetic affiliation (%similarity)T106 Clostridiaceae Clostridium subterminale (95)T110 Clostridiaceae Clostridium subterminale (96)T112 Clostridiaceae Clostridium subterminale (95)T116 Clostridiaceae Clostridium subterminale (95)T124 Enterobacteriaceae Citrobacter freundii (98)T127 Enterobacteriaceae Citrobacter freundii (99)T128 Enterobacteriaceae Citrobacter freundii (98)T129 Enterobacteriaceae Citrobacter freundii (97)T130 Enterobacteriaceae Citrobacter freundii (98)T131 Enterobacteriaceae Enterobacter dissolvens (97)T137 Enterobacteriaceae Enterobacter dissolvens (98)T138 Enterobacteriaceae Enterobacter dissolvens (98)T139 Enterobacteriaceae Enterobacter dissolvens (98)T180 Enterobacteriaceae Enterobacter dissolvens (98)T181 Clostridiaceae Clostridium thiosulfatireducens (96)T182 Clostridiaceae Clostridium thiosulfatireducens (96)T183 Clostridiaceae Clostridium thiosulfatireducens (96)T159 Desulfomicrobiaceae Desulfomicrobium escambium(99)T160 Desulfomicrobiaceae Desulfomicrobium escambium(99)T161 Desulfomicrobiaceae Desulfomicrobium escambium(99)T162 Desulfomicrobiaceae Desulfomicrobium escambium(98)T164 Desulfomicrobiaceae Desulfomicrobium escambium(99)T136 Desulfovibrionaceae Desulfovibrio longus (99)T148 Desulfovibrionaceae Desulfovibrio longus (98)T149 Desulfovibrionaceae Desulfovibrio longus (99)T154 Desulfovibrionaceae Desulfovibrio vulgaris (99)T155 Desulfovibrionaceae Desulfovibrio vulgaris (99)T156 Desulfovibrionaceae Desulfovibrio vulgaris (99)

Results 99
TERI university-Ph.D. Thesis, 2009
Figure.5.18: Evolutionary relationships of 18 taxa (linearized) The evolutionary history was inferred using the Neighbor-Joining method . The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Maximum Composite Likelihood method and are in the units of the number of base substitutions per site. Phylogenetic analyses were conducted in MEGA4
100
99
49
100
100
100
100
70
59
100
89
100
29
0.000.050.100.15
T124 (5)Citrobacter freundii (AB24454.1)
T131 (5)
Pantoea agglumerans (EF428997.1)
Enterobactoer cloacae (EU048321.1)
Enterobacter dissolvens (Z96079.1)
T181 (3)
T106 (4)
T159 (5)
T154 (3)
Clostridium thiosulfatireducens (AB294141.1)
Clostridium subterminale (AF281842.1)
Clostridium botulium (M59087.1)
Desulomicrobium escambium (AJ 277886.1)
Desulfovibrio alaskensis (DQ867001.1)
Desulfovibrio vulgaris (AB294142.1)
Desulfovibrio longus (AY 359867.1)
T136 (3)

Results 100
TERI university-Ph.D. Thesis, 2009
The Clade III was dominating with the cultures affiliated to Desulfovibrio vulgaris, Desulfovibrio longus and Desulfomicrobium escambium. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) was shown next to the branches. The phylogenetic tree was linearized assuming equal evolutionary rates in all lineages. All positions containing gaps and missing data were eliminated from the dataset (Complete deletion option). There were a total of 376 nucleotide positions in the final dataset.
16S rDNA fingerprinting and cluster analysis of the isolated strains based on Denaturating Gradient Gel Electrophoresis profile
The 16S rDNA fingerprints of the strains elucidated different patterns revealing inter and intraspecies diversity between the isolates (figure 5.19). The cluster analysis of the 28 strains isolated from production water sample of Uran process plant showed that all the strains were delineated into 17 genotypic clusters. The largest genotypic cluster was cluster X, with 5 isolates strains (T136, T148, T149, T161 and T162). The strains affiliated to Desulfomicrobium escambium clustered in 3 different genotypic clusters as T159 and T160 clustered with genotypic cluster XI, T161 and T162 clustered with genotypic cluster X and T164 clustered in genotypic cluster XVIII. All the 3 strains (T136, T148 and T149) affiliated to Desulfovibrio longus also clustered in genotypic cluster X, The strains belonging to Clostridium subterminale delineated in to genotypic cluster I (T112, T110 and T106) and II (T116) and strains belonging to Desulfovibrio vulgaris grouped into genotypic cluster III (T154 and T155) and IV (T156). The five isolates belonging to Enterobacter dissolvens grouped in genotypic clusters V (T131), VII (T137 and T138) and VIII (T139 and T180). The three isolates belonging to Clostridium thiosulfatireducens clustered in three different genotypic clusters (V, XVI and XVII). Bacterial isolates phylogenetically affiliated to Citrobacter freundii delineated into 4 genotypic clusters. Bacterial strains T124 and T127 grouped in genotypic cluster XII while T128, T129 and T130 delineated separately in genotypic clusters XIII, XIV and XV.

Resul
TERI u
FigureUran pXVIII:
ts
universit
e 5.19: productio: Genotyp
% Similarity
ty-Ph.D.
Genotypion water pic Clust
Thesis,
c clustesamples
ters, Num
101
2009
er analys by Braymber in t
1
sis of thy-Curtis he right
he straiclustersare isol
ns isolas analyslate numb
ated fromis. (I-ber)
m -

Results 102
TERI university-Ph.D. Thesis, 2009
Evaluation of diversity of culturable sulfidogenic bacteria in production water collected from oil-water separator located at Ghandhar effluent treatment plant (ETP)
Description of sample collection site and chemical characteristics of the collected sample
Ghandhar ETP (22.16 N, 73.12 E) is located in western India in the state of Gujarat. The fields are located near Ankaleshwar, 90 kms from Vadodara city. Ghandhar ETP receives crude oil from various oil reservoirs in the oil field. Production water collected from Ghandhar ETP was translucent with tiny black particles. The black particles settled down after sometime. The production water sample has very high sulfide concentration (350 mg.L-1) and sulfate concentration was 5.6 mg.L-1. The total dissolved soilds (TDS) and Suspended solid were 4300 mg.L-1 and 350 mg.L-1. Dissolved oxygen was almost negligible. The temperature of the production water sample collected was 42 °C and pH was equal to 8.8.
Microbiological characterization
Bacterial strains isolated from production water sample collected from Ghandhar ETP were rod shaped (short and long). Of the 12 bacterial strains isolates, 2 were gram-negative while other stained gram-positive. The gram negative bacteria (T192 and T193) were isolated from API RP 38 medium (figure 5.21). These isolates were short rod in morphology and were growing optimally at 2% NaCl concentration and 40 °C. The H2S production from bacterial isolates T192 and T193 was 80 and 120 mg.L-1. Both the isolates were able to grow from salinity range 0.5-10% and highest temperature at which growth was observed was 45°C. All the Gram positive strains were isolated from Lyngby medium. The bacterial strains T219 and T220 were long rods in shape while other strains (T184, T185, T186, T187, T188, T189, T190 and T221) were thick rods in shape (Figure 5.20). The bacterial strains isolated from Lyngby medium have optimal salinity of 0.5% and optimal growing temperature of 37°C. The sulfide production ranges from 150 to 250 mg.L-1 . It was observed that none of the cultures was isolated from Ghandhar production water samples enriched in Baar’s medium. The morphology and growth characteristics of the bacterial strains isolated from production water collected from Ghandhar ETP are shown in table 5.10.

Results 103
TERI university-Ph.D. Thesis, 2009
of the band represent isolate number)
Table 5.10: Morphological physical and biochemical characteristics of bacterial strains isolated from Ghandhar production water samples
*values of H2S production after 28 days
Phylogenetic identification of the isolated bacterial strains from Ghandhar production water sample
DGGE analysis of isolates from Ghandhar production water samples showed very low culturable diversity with only 3 different phylotypes. A total of 10 and 2 isolates were isolated from production waters enriched in Lyngbay and API RP 38 medium. No bacteria were isolated from Baar’s medium. Bacterial strains which were isolated from Lyngby medium were affiliated to Clastridium subterminale and Enterobacter dissolvens, while all isolated bacterial strains isolated from enriched samples of API RP38 medium were affiliated to Desulfovibrio vulgaris. Microbiological characterization of the isolated strains and their phylogenetic affiliation is represented in the table 5.11.
Figure5.20: Light microscopic pictures of isolate T185
Figure5.21: Light microscopic pictures of isolate T192
Isolate Number
Isolation Medium Cell Morphology Gram Staining
Salinity Tolerence
%
Optimal Salinity
%
Growth Temp °C
H2S production range mg L-1*
T219 Lyngby Long Rod Gram negative 0.5-10 0.5 37 150T220 Lyngby Long Rod Gram negative 0.5-10 0.5 37 180T184 Lyngby Thick Rods Gram positive 0-6 0.5 37 150T185 Lyngby Thick Rods Gram positive 0-6 0.5 37 160T186 Lyngby Thick Rods Gram positive 0-6 0.5 37 250T187 Lyngby Thick Rods Gram positive 0-6 0.5 37 210T188 Lyngby Thick Rods Gram positive 0-6 0.5 37 130T189 Lyngby Thick Rods Gram positive 0-6 0.5 37 220T190 Lyngby Thick Rods Gram positive 0-6 0.5 37 210T221 Lyngby Thick Rods Gram positive 0-6 0.5 37 240
T192 Modified API RP 38 Short rods Gram negative 0.5-10 2 40 80
T193 Modified API RP 38 Short rods Gram negative 0.5-10 2 40 120

Results 104
TERI university-Ph.D. Thesis, 2009
Table 5.11: Phylogenetic characterization of the isolated bacterial strains from Ghandhar production water sample
Figure.5.22: Evolutionary relationships of 12 taxa. The evolutionary history was inferred using the Neighbor-Joining method. The optimal tree with the sum of branch length = 0.68507469 is shown. The tree is drawn to scale, with branch lengths in the same units as those of the evolutionary distances used to infer the phylogenetic tree. The evolutionary distances were computed using the Maximum Composite Likelihood method and are in the units of the number of base substitutions per site. Phylogenetic analyses were conducted in MEGA4
% Similarity
Isolate Number Family Phylogenetic affiliation (%similarity)T219 Enterobacteriaceae Enterobacter dissolvens (98)T220 Enterobacteriaceae Enterobacter dissolvens (98)T184 Clostridiaceae Clostridium subterminale (95)T185 Clostridiaceae Clostridium subterminale (95)T186 Clostridiaceae Clostridium subterminale (95)T187 Clostridiaceae Clostridium subterminale (96)T188 Clostridiaceae Clostridium subterminale (95)T189 Clostridiaceae Clostridium subterminale (96)T190 Clostridiaceae Clostridium subterminale (95)T221 Clostridiaceae Clostridium subterminale (95)T192 Desulfovibrionaceae Desulfovibrio vulgaris (99)T193 Desulfovibrionaceae Desulfovibrio vulgaris (99)
100
99
50
100
85
100
100
57
0.000.020.040.060.080.100.120.14
Enterobactoer cloacae (EU048321.1)
Pantoea agglumerans (EF428997.1)
Enterobactor dissolvens (Z96079.1)
219 (2)
Clostridium thiosulfatireducens (AB294141.1)
T184 (8)
Clostridium subterminale (AF 241842.1)
Clostridium botulium(M59087.1)
Desulfovibrio alaksensis (DQ86700.1)
Desulfovibrio longus (AY 359867.1)
T192 (2)
Desulfovibrio vulgaris (AB 294142.1)

Results 105
TERI university-Ph.D. Thesis, 2009
Phylogenetic tree analysis of the bacterial cultures revealed presence of 3 phylogenetic clades (figure 5.22). Phylogenetic clade I was composed of members of Enterobacteriaceae. The bacterial culture belonging to this clade were afiiliated to the genera Enterobacter. Phylogenetic clade II consist of members of Clostridiaceae family with in genera Clostridium. Cultures belonging to phylogenetic clade III were affiliated with Desulfovibrionaceae family of delta-proteobacteria devision. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 replicates) are shown next to the branches. The phylogenetic tree was linearized assuming equal evolutionary rates in all lineages. All positions containing gaps and missing data were eliminated from the dataset (Complete deletion option). There were a total of 394 positions in the final dataset.
16S rDNA fingerprinting and cluster analysis of the isolated strains based on Denaturating Gradient Gel Electrophoresis profiles
The 16S rDNA fingerprints of the isolated strains revealed the presence of variation in the 16S rDNA amplicon of the strains which were affiliated to the same species. The cluster analysis of the 12 strains isolated from production water sample of Ghandhar ETP showed that all the strains were delineated into 10 genotypic clusters (figure 5.23). The bacterial isolates T186, T187, T188, T219, T220 and T221 l delineated separately in genotypic clusters I, III, V, VI, VIII and IX. The strains (T192 and T193) affiliated to the species Desulfovibrio vulgaris clustered in a genotypic cluster X. Bacterial strains T184 and T185 which were affiliated to Clostridium subterminale clustered in genotypic cluster VII. The bacterial strains T189 and T190 grouped in a genotypic cluster IV.
The molecular techniques like polymerase chain reaction and DGGE are very useful in evaluating the diversity of microbes in any ecosystem. The banding pattern of PCR amplified product of one particular gene when subjected to DGGE, gives a clear cut idea of diversity of the microbial community having that gene. Total microbial diversity of the production water samples was analyzed with an aim to investigate the presence of sulfidogenic bacteria which cannot be cultivated by culturing techniques. To explore the total diversity bisulfite reductase gene was employed. Dissimilatory sulfite reductase gene (dsr) encode for dissimilatory sulfite reductase enzyme. This enzyme catalyses the formation of sulfide from sulfite and hence can be used to study diversity of the bacteria which produce hydrogen sulfide. As the dsr gene is widely studied in different ecosystems, both nucleotide and protein sequences of this gene from various microorganism are well documented. The dsr nucleotide sequence from all the known species

Results 106
TERI university-Ph.D. Thesis, 2009
harbouring this gene is available in the public database of NCBI World Wide Web. Therefore, this gene was chosen for the phylogenetic identification of these microbes.
Figure 5.23: Genotypic cluster analysis of the strains isolated from Ghandhar production water samples by Bray-Curtis clusters analysis
Diversity of non-culturable SRB in production water samples
Analysis of SRB diversity in production water samples by Denaturating Gradient Gel Electrophoresis of dsrB gene
Production water samples which were collected in 10-liter carboys were used for the DNA extraction. The DNA extraction was done by centrifugation of production waters to harness the maximum bacterial population. Pallet derived from centrifugation of the production water was dark brown in colour. It contains precipitated soil particles, mineral salts and black particles corroded and small pieces of bio-films developed in the system. During centrifugation both plank-tonic and sessile bacteria were pallet down. Pallets from different tubes were pooled and consequential pallet was used for DNA extraction. Other methods which were used to collect microbial density of production water were by filtration of production water with 0.22 micron filters and Mobio water sample DNA extraction kit. DNA yields were very low by these methods when compared to centrifugation method. Mobio water DNA extraction kit yielded a pure DNA
100
9590858075706560555045403530252015
220
122
221
B
190
189
187
188
184
185
186
219
193
192
IIIIIIIVVVIVIIVIIIIXXXIXII

Resul
TERI u
but theexperimwater Electropproductpopulatgeograp
PCR a
PCR amand DSapproxiany amforwardband ofDGGE a
FigurenegatiDesulfDesulfDesulfDesulfDesulfSRB12:
ts
universit
e quantity ments were
contains hphoresis oftion water tion of sulfiphical locati
amplifi
mplificationSR4R primeimately 350
mplicon. Semd primer) af 390 bp apanalysis.
e: 5.24: ive confobulbus fosarcinafomicrobifobulbus fomicrobi: Clone o
ty-Ph.D.
was low w done with high densitf the bisulsamples sh
idogenic comions.
ication
n of the dsrBers sets. All 0 bp. The Dmi-nested Pand DSR4Rproximately
PCR ampntrol:
proponia viriabium escrhabdof
ium norvof dsrB g
350 bp
Thesis,
while no D the DNA ety bacterialfite reduct
howed very mmunities
of dsr
B gene of p the pure SRNA of E. co
PCR with thR, using PCR
y. Semi-nes
plificatiSRB1:
icus, SRbilis, Scambienseformis, vegicum, gene used
107
2009
DNA was rextracted fra which ptase (dsr) diverse ba in the prod
rB gene
pure SRB stRB strains g
oli which wahe primers R product osted approa
on of dsDesulfot
RB3: DesSRB5: De, SRB7:SRB9: DSRB11:
d as posi
retrieved urom centrif
produce H2gene amp
anding patteduction wate
e fragme
trains was cgenerated aas used as n gcDSRp20of first PCR
ach was used
srB gene tomaculumsulfobactesulforha: DesulDesulfovib
Desulfotive con
using filtratfugation me2S. Denatu
plified fromern. This reer samples
ent
carried out a single ampnegative con060F (GC cR as templad to make t
of pure m aeronter vibraphlus vlfovibriobrio terotomaculutrol
350 bp
tion methoethod. The urating Gr
m DNA isoevealed hig collected fr
by using Dplified PCR
ntrol did noclamp was ate, generatthe amplico
SRB strnauticum,rioformisvoculatuso gigasrmitidis,um aeron
od. Further productionradient Gelolated fromghly diverserom various
DSRp2060FR product ofot generatedattached toted a singlen useful for
rains and, SRB2:s, SRB4:s, SRB6:, SRB8:, SRB10:nauticum,
r n l
m e s
F f
d o e r
d
:
,

Results 108
TERI university-Ph.D. Thesis, 2009
Semi-nested PCR overcome the drawback of poor amplification while using direct PCR with GC clamp attached primer to amplifying target sequence. Hence semi nested strategy where amplification product of the first PCR can be used as template for consecutive PCR with GC-clamp attached primers was successfully used. This strategy has been used by several authors before who observed difficulty in amplifying the fragment directly with GC-Clamp attached primer set. Earlier reports also mention this type of semi-nested PCR strategy (Nicolaisen & Ramsing, 2002; Hendrickx et al., 2006). Figure 5.24 showed the dsrB amplicon of 350 bp amplified from DNA extracted from pure strains of sulfate reducing bacteria.
Differentiation of dsrB-gene fragment by DGGE
It is utmost important that the dsrB gene product of pure SRB strains should differentiated from each other when subjected to the DGGE. Differentiation of the dsrB gene is based on the migration of the amplified product in the DGGE. Migration of the bands at the different position in the gel attribute to the fact that the amplicon was highly diverse among the member of SRB and therefore the dsrB gene based DGGE can be potentially used for diversity studies.
When semi-nested PCR amplified dsrB gene products were subjected to DGGE, all the amplicons derived from pure SRB strains showed a single bright band that migrated at a particular position. Species belonging to same genera like Desulfobulbus proponicus (SRB8) and Desulfobulbus rhabdoformis (SRB2); Desulfomicrobium norvegicum (SRB10) and Desulfomicrobium escambiense (SRB6); Desulfovibrio termitidis (SRB9) and Desulfovibrio gigas (SRB7) showed distinctly separated and distantly migrated bands (fig. 5.27). Thus dsrB gene based DGGE was efficient in discriminating between the different SRB strains.
It was very important to know that primers indiscriminately binds to the dsr gene of all the strains of SRB when mixture of DNA of all the pure strains was used for two step semi-nested PCR amplification and subsequently loaded to DGGE. The specificity of the primer, PCR conditions and DNA extraction procedures can significantly influence PCR results. The amplicon was separated in form of different bands which migrated distinctly in the DGGE gel. This revealed that dsrB gene primer pairs equally bind to the dsrB gene of all the species without any inclination towards any species.
The simulated melting temperature (as generated during melt-curve analyses in qRT-PCR) of the dsrB gene amplicons of pure strain showed variation from 85.7 ˚C to 92.2 ˚C. The dsrB gene amplicon with lower melting temperature showed lesser migration in comparison to amplicons with higher melting temperature. It was noted that each dsrB amplicon with different position in DGGE were also having different melting temperature.

Results 109
TERI university-Ph.D. Thesis, 2009
DsrB fragment of few pure strains showed an additional faint band in the DGGE gel (SRB1, SRB9, SRB10 and SRB11). The reason for this could be the degeneracy of the dsrB gene primers or slight difference in the targeted sequence of the dsrB gene (Throbäck et al., 2004).
In the figure 5.25, PCR amplified dsrB gene amplicon of ten pure SRB strains was subjected to DGGE. SRB 5 retained top position in the gel, which corresponds to lowest melting temperature of the amplicon. SRB 7 retained the lowest position in the DGGE gel which corresponds to the highest melting temperature when compared to other amplicons.
dsrB gene based DGGE ladder
A dsrB gene based DGGE ladder was constructed by mixing dsrB gene amplicons of the pure SRB strains. The mixture of various bands in the ladder migrated across the full denaturation range. DGGE results were reproducible for both single and mixture DSR amplicons from pure SRB strains.
Figure 5.25: DsrB gene based DGGE for pure SRB strains: SRB1: Desulfotomaculum aeronauticum, SRB2: Desulfobulbus proponicus, SRB3: Desulfobacter vibrioformis, SRB4: Desulfosarcina viriabilis, SRB5: Desulforhaphlus voculatus, SRB6: Desulfomicrobium escambiense, SRB7: Desulfovibrio gigas, SRB8: Desulfobulbus rhabdoformis, SRB9: Desulfovibrio termitidis, SRB10: Desulfomicrobium norvegicum, SRB11: Desulfotomaculum aeronauticum

Results 110
TERI university-Ph.D. Thesis, 2009
Phylogenetic analysis and DGGE profile of dsrB gene sequences in oil field samples, India
The diversity of dsrB genes in SRB-communities from production waters of various oil fields was investigated (figure 5.26 ) and depicted in a phylogenetic tree (figure 5.29 ). The dsrB gene was amplified by using genomic DNA extracted from production water samples, collected from various oil fields. The dsrB gene based DGGE community profile of production water samples from north-eastern oil fields (D1 and K1) displayed similar DGGE banding profile (figure 5.26 ). The community profile of D1 and K1 were having 3 similar bands designated as K1-F2/D1-E3, D1-B5/K1-C4 and D1-F7/K1-F7. The deduced amino acid sequence of D1-F7 and K1-F7 exhibited maximum similarity with the amino acid sequence of Desulfovibrio vulgaris dsrB protein (99%). Similarly the translated amino acid sequence of D1-B5 and K1-C4 showed maximum similarity with the Desulfotomaculum thermoacetoxidans (95%) dsrB protein sequence.
Figure 5.26: DGGE analysis of dsrB gene amplified from production water samples

Results 111
TERI university-Ph.D. Thesis, 2009
Figure 5.27: Phylogenetic tree of translated dsrB gene sequences obtained from various production waters

Results 112
TERI university-Ph.D. Thesis, 2009
These sequences clustered within the dsrB protein lineage of the Desulfotomaculum species that were monophyletic with delta-proteobacterial dsr sequences (Klein et al., 2001). Deduced amino acid sequences of K1-F2 and D1-E3 clustered with Desulfomicrobium macestii (98%). The bands D1-E1 and D1-E8 also appeared in D1 production water samples. The band D1-E1 exhibited maximum similarity (98%) with dsrB amino acid sequence of Desulfotomaculum aeronauticum, forming a divergent cluster with the genera Desulfosporosinus and Desulfitobacterium (figure 5.27 ). The deduced amino acid sequences of D1-E8 clustered with the dsrB protein sequence of sulfite-reducing bacteria Desulfitobacterium hafniense (85%) . Bands K1-B1 also showed maximum similarity with dsrB protein sequence of Desulfotomaculum thermoacetoxidans.K2 (Kalol) production water samples displayed rich dsrB-DGGE profile with 8 intense bands. Deduced amino acid sequences of seven clones showed significant similarity with dsrB protein sequences of various SRB. The deduced amino acid sequence of clones of K2 samples exhibited similarity with dsrB protein sequence of Desulfotomaculum aeronauticum (K2-F7, K2-E5, 96%), Desulfacinum infernum (K2-C6, K2-B2, 97%) and with a member of the Desulfobacteriaceae family (K2-B3, 74%). The deduced protein sequence of K2-C5 affiliated for 89% to that of the protein sequence of Desulfobulbus rhabdoformis, which is a member of the Desulfobulbaceae (figure 5.27 ). K2-B8 (93%) exhibited maximum homology with the dsrB gene sequence of Thermosinus carboxydivorans, a gram negative, thermophilic and chemolithotrophically growing sulfite-reducing bacterium (Sokolova et al., 2004).
The dsrB- based DGGE community profile of D2 and G2 showed little diversity with 2 and 4 dominating bands respectively. DGGE profile of both the samples had one band at similar position in DGGE gel (D2-B4 and G2-B5): deduced amino acid sequence of both the bands showed 98% similarity with Desulfotomaculum aeronauticum. The deduced dsrB based amino acid sequences from G2 oil field were affiliated to Desulfomicrobium macestii (G2-B4, 97%) and Desulfatibacillum alkenivorans (G2-D2, 86%).
Quantification of SRB in production water samples
Quantification of SRB in the production water samples is done by quantitative real-time PCR of the dsr gene. Estimation of the functional metabolic genes by using molecular methods is the most accurate method for quantification of microbes with similar metabolic pathway.
The quantitative real-time method to estimate dsr gene using Sybr-Green chemistry revealed lowest detection limit of 1o2 dsr gene copies. To design the accurate method for quantification it

Results 113
TERI university-Ph.D. Thesis, 2009
is important to use the primers which can indiscriminately amplify the dsr gene of all the known SRB as well as optimum concentration of these primers in the reaction so to minimize primer-dimer formation. Therefore, the same primer pair which was used for DGGE analysis was employed for quantitative real-time PCR assay
Performance, sensitivity and detection limits, of dsrB gene standard curve
PCR with primers DSR1F and DSR4R resulted in amplification of 1.9 kb dsrAB gene fragment of when Desulfomicrobium norvegicum (DSMZ) DNA was used as template. Likewise, 1.5 kb 16S rDNA gene of hydrogenophaga species was amplified with 27F and 1492R PCR primers. Recombinant plasmids harbouring the dsrAB and 16S rDNA gene were extracted from the transformed cells. Sequencing of plasmids with M13 primer confirmed 99% similar match with Desulfomicrobium norvegicum dsrAB and hydrogenophaga species 16S rDNA gene sequence in GenBank database. Triplicate serial dilution series of extracted plasmids were used for preparation of standard curve. Linearity and reproducibility of the standard curve was tested. The optimized primer ratio was 400nM for each primer when used in qRT-PCR. The dsrB standard curve (figure 5.28 ) was linear from 4.5 × 107 to 4.5 × 102 copies per reaction with the slope of -3.59 (R2 value =0.994). The 16S rDNA standard curve (figure 5.29 ) was linear from 9 × 108 to 90 copies per reaction with a slope of -3.025 (R2 value =0.997).
Figure 5.28: Standard curve for dsrB gene copies calculation

Results 114
TERI university-Ph.D. Thesis, 2009
Figure 5.29: Standard curve for 16S rDNA gene copies calculation
The efficiency of standard curves to determine absolute copies in the chromosomal DNA
samples, was estimated by qRT-PCR against the dilution series of Desulfovibrio vulgaris
chromosomal DNA (table 5.12 ). Copy number of Desulfovibrio vulgaris genome was calculated,
based on DNA concentration. Graph plotted between theoretical copy number of Desulfovibrio
vulgaris versus Ct value was linear from 2.5 × 105 to 25 copies per reaction (r2 = 0.99, slope = -
3.26, Effciency = 1.03). To imitate the effect of foreign DNA in the samples equal amount of
E.coli chromosomal DNA was mixed with the calculated amount of Desulfovibrio vulgaris DNA
(table 5.12 ).
Table 5.12: Effect of chromosomal DNA, foreign DNA and inhibitors on the efficiency of qRT-PCR assay
A dilution series of this mixture was made according to Desulfovibrio vulgaris DNA copy
number (figure 5.30). Standard curve so obtained, was linear from 2.5 × 105 to 25 copies per
Samples
Theoretical copy
number
Calculated copy number via
standard curve
Calculated copy number via standard
curve with E. coli DNA
Calculated copy number via standard
curve with Environmental Sample DNA
D.vulgaris 1 2,50,000 1,14,103 2,40,192 2,23,194 D.vulgaris 2 25,000 26,006 25,542 31,370 D.vulgaris 3 2,500 2,657 2,252 1,992 D.vulgaris 4 250 816 292 250 D.vulgaris 5 25 23 42 44
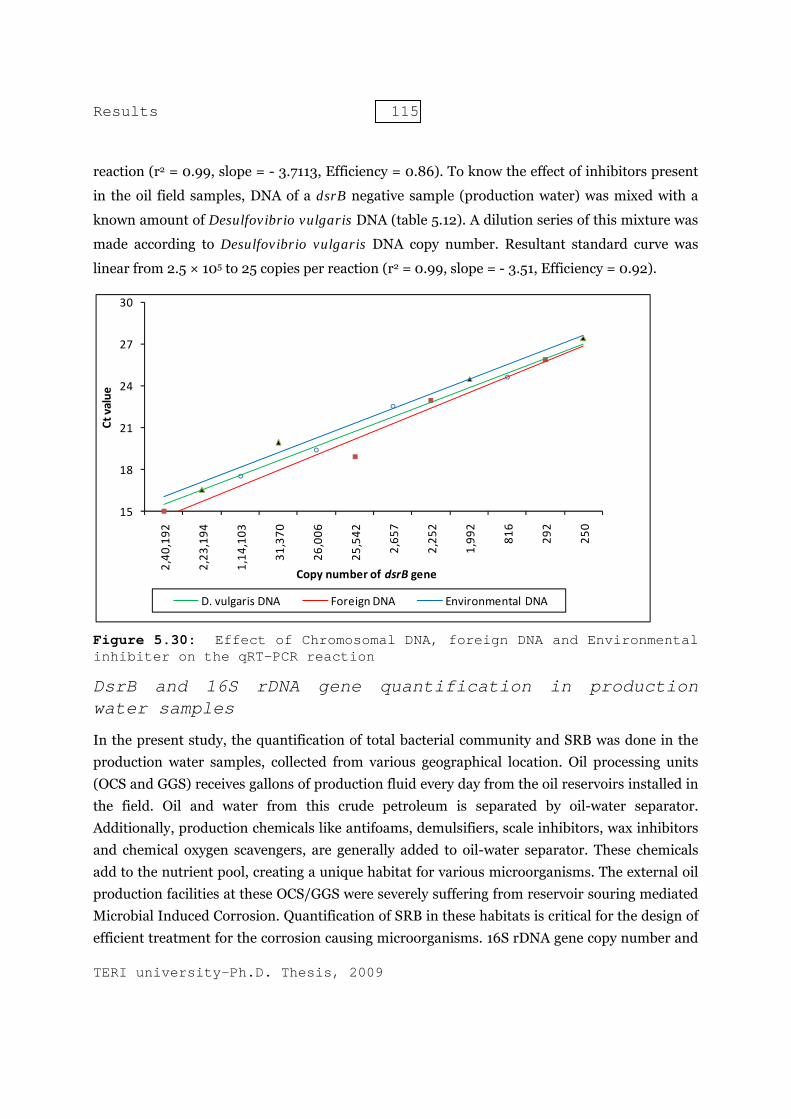
Results 115
TERI university-Ph.D. Thesis, 2009
reaction (r2 = 0.99, slope = - 3.7113, Efficiency = 0.86). To know the effect of inhibitors present
in the oil field samples, DNA of a dsrB negative sample (production water) was mixed with a
known amount of Desulfovibrio vulgaris DNA (table 5.12). A dilution series of this mixture was
made according to Desulfovibrio vulgaris DNA copy number. Resultant standard curve was
linear from 2.5 × 105 to 25 copies per reaction (r2 = 0.99, slope = - 3.51, Efficiency = 0.92).
Figure 5.30: Effect of Chromosomal DNA, foreign DNA and Environmental inhibiter on the qRT-PCR reaction
DsrB and 16S rDNA gene quantification in production water samples
In the present study, the quantification of total bacterial community and SRB was done in the production water samples, collected from various geographical location. Oil processing units (OCS and GGS) receives gallons of production fluid every day from the oil reservoirs installed in the field. Oil and water from this crude petroleum is separated by oil-water separator. Additionally, production chemicals like antifoams, demulsifiers, scale inhibitors, wax inhibitors and chemical oxygen scavengers, are generally added to oil-water separator. These chemicals add to the nutrient pool, creating a unique habitat for various microorganisms. The external oil production facilities at these OCS/GGS were severely suffering from reservoir souring mediated Microbial Induced Corrosion. Quantification of SRB in these habitats is critical for the design of efficient treatment for the corrosion causing microorganisms. 16S rDNA gene copy number and
15
18
21
24
27
30
2,40
,192
2,23
,194
1,14
,103
31,370
26,006
25,542
2,65
7
2,25
2
1,99
2
816
292
250
Ct value
Copy number of dsrB gene
D. vulgaris DNA Foreign DNA Environmental DNA

Results 116
TERI university-Ph.D. Thesis, 2009
dsrB gene copy number was calculated in the DNA, extracted from various production water samples (figure 5.31). All the samples were quantified in triplicate and corresponding average value were converted to gene copies per ml of produced water (table 5.13 ).
Table 5.13: Average dsr and 16S rDNA gene copies per ml of production water and percentage of dsr copies out of 16S rDNA copies
Figure 5.31: The 16S rDNA and dsrB gene copies in various production water samples
Standard deviation in the samples was also calculated. Production water samples from north-
eastern and western part had approximately equal number of sulfate reducers in the range of 105
copies per ml of production water except G2 production water with approximately 106 dsrB gene
copies per ml of production water. Indeed, highest dissolved sulfide was recorded in G2
production waters. The 16S rRNA gene copies were in the magnitude range of 106 -107 copies per
ml of production water with highest copies in D2 production waters. D2 sample also showed
Oil Collection Stations
16S rDNA gene copies/ml of
production water
Average 16S rDNA gene copies per Genome
DSR gene copies/ml of
production water
% DSR gene copies/ml of 16S
rDNA gene copies
% SRB/ml of total
bacterial counts
D1 2824534 807009 3,36,387 11.9 41.6 K1 2589300 739800 3,36,429 12.99 45.47 K2 8046906 2299116 6,58,122 8.1 28.62 D2 15274574 4364164 2,49,651 1.63 5.72 G2 11243134 3212324 7,55,426 6.71 23.51

Results 117
TERI university-Ph.D. Thesis, 2009
lowest dsrB gene copies. This could be attributed to the fact that SRB have to compete with
other bacteria present in the production water and hence, absolute number and proportion of
SRB remain low in D2 production waters (figure 5.31). Earlier it was demonstrated that nitrate
reducing-sulfide oxidizing bacteria (NR-SOB) can outcompete the SRB for energy source and
hence mitigate oil reservoir souring (Haveman et al., 2005). Klappenbach et al. (2001) reported
that 16S rDNA gene copies varies in species and average 3.6 16S rDNA copies per genome is
present. Accordingly percentage of dsrB gene copies and percentage of SRB in total bacterial
community was calculated (table 5.13)
Evaluating biocide efficiency in the production water samples collected from South Kadi, Gujarat using quantitative PCR assay
The most important risk management tool to control to control microbial activity in offshore oil production systems is biocide dosing treatment. To obtain a cost effective biocide treatment it is important to determine biocide efficiency using quantitative (micro) biological assays. Therefore, it was necessary to check the reliability of the quantitative real-time PCR assay to quantify SRB in the production water samples dosed with different biocidal concentration. To achieve this aim fresh production water samples were collected from South Kadi, Gujarat. The effectiveness of the biocide depends on the nature of microorganism to be eliminated and the physical and chemical properties of the system to be operated.
The strategy followed to evaluate efficiency of six different biocides to control sulfidogenic bacteria was initial screen of the biocides by estimation of accumulated hydrogen sulfide at different concentration of biocides (25, 50, 100, 150 and 200 ppm) after 1 and 7 days. The most effective biocide was tested for its efficiency to work at lower concentration. This efficiency was estimated in terms of decrease in bisulfite reductase gene copy number by quantitative PCR assay which were developed in the earlier objectives.
Primary screening of commercial biocides in south Kadi production water enriched samples
Primary screening of the biocides was focused on selection of the suitable biocide against SRB in the production water sample collected from oil-water separator. All the six screened biocides were earlier reported to be effective in different systems. Growth was monitored by visually observing the cultures for turbidity and confirmed by H2S accumulation after 1 and 7 days. Six
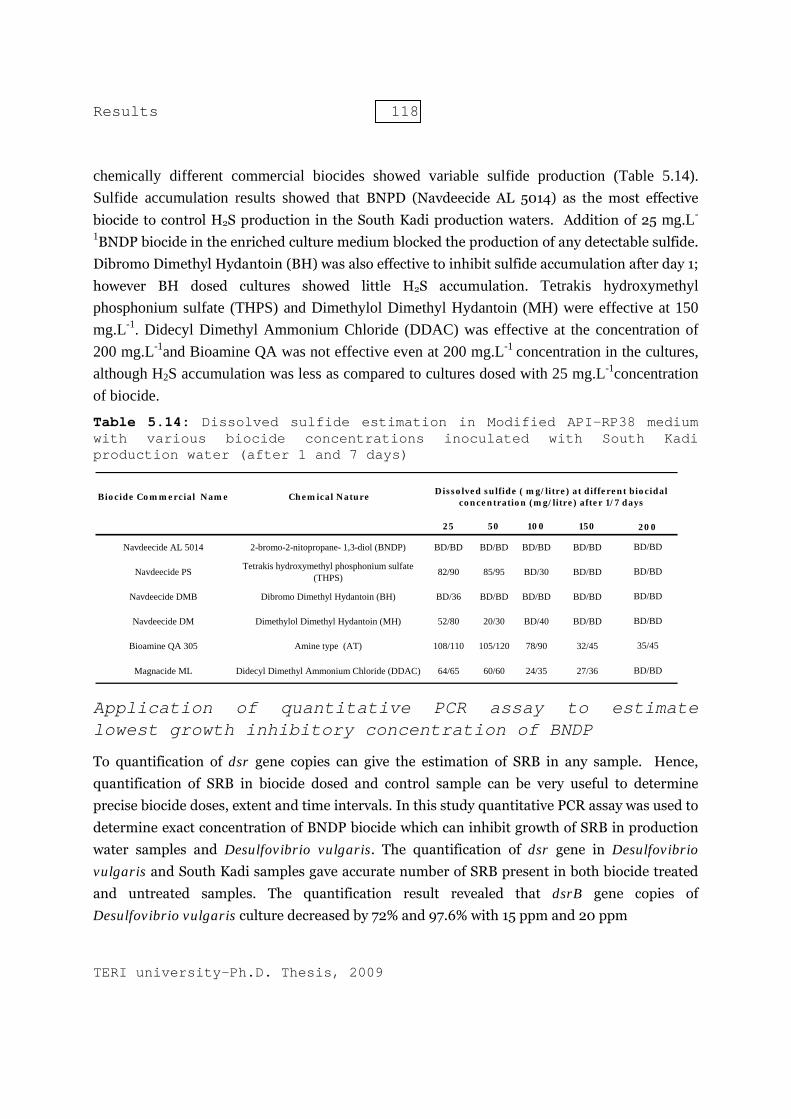
Results 118
TERI university-Ph.D. Thesis, 2009
chemically different commercial biocides showed variable sulfide production (Table 5.14). Sulfide accumulation results showed that BNPD (Navdeecide AL 5014) as the most effective biocide to control H2S production in the South Kadi production waters. Addition of 25 mg.L-
1BNDP biocide in the enriched culture medium blocked the production of any detectable sulfide. Dibromo Dimethyl Hydantoin (BH) was also effective to inhibit sulfide accumulation after day 1; however BH dosed cultures showed little H2S accumulation. Tetrakis hydroxymethyl phosphonium sulfate (THPS) and Dimethylol Dimethyl Hydantoin (MH) were effective at 150 mg.L-1. Didecyl Dimethyl Ammonium Chloride (DDAC) was effective at the concentration of 200 mg.L-1and Bioamine QA was not effective even at 200 mg.L-1 concentration in the cultures, although H2S accumulation was less as compared to cultures dosed with 25 mg.L-1concentration of biocide.
Table 5.14: Dissolved sulfide estimation in Modified API-RP38 medium with various biocide concentrations inoculated with South Kadi production water (after 1 and 7 days)
Application of quantitative PCR assay to estimate lowest growth inhibitory concentration of BNDP
To quantification of dsr gene copies can give the estimation of SRB in any sample. Hence, quantification of SRB in biocide dosed and control sample can be very useful to determine precise biocide doses, extent and time intervals. In this study quantitative PCR assay was used to determine exact concentration of BNDP biocide which can inhibit growth of SRB in production water samples and Desulfovibrio vulgaris. The quantification of dsr gene in Desulfovibrio vulgaris and South Kadi samples gave accurate number of SRB present in both biocide treated and untreated samples. The quantification result revealed that dsrB gene copies of Desulfovibrio vulgaris culture decreased by 72% and 97.6% with 15 ppm and 20 ppm
25 50 100 150
Navdeecide AL 5014 2-bromo-2-nitopropane- 1,3-diol (BNDP) BD/BD BD/BD BD/BD BD/BD
Navdeecide PS Tetrakis hydroxymethyl phosphonium sulfate (THPS) 82/90 85/95 BD/30 BD/BD
Navdeecide DMB Dibromo Dimethyl Hydantoin (BH) BD/36 BD/BD BD/BD BD/BD
Navdeecide DM Dimethylol Dimethyl Hydantoin (MH) 52/80 20/30 BD/40 BD/BD
Bioamine QA 305 Amine type (AT) 108/110 105/120 78/90 32/45
Magnacide ML Didecyl Dimethyl Ammonium Chloride (DDAC) 64/65 60/60 24/35 27/36
BD/BD
35/45
BD/BD
Biocide Commercial Name
200
BD/BD
BD/BD
BD/BD
Chemical Nature Dissolved sulfide ( mg/litre) at different biocidal concentration (mg/litre) after 1/7 days

Results 119
TERI university-Ph.D. Thesis, 2009
Figure 5.32: DsrB gene copies reduction and dissolved sulfide per ml of Desulfovibrio vulgaris at different Biocidal concentration
Figure 5.33: DsrB gene copies reduction and dissolved sulfide per ml in South Kadi Production water at different Biocidal concentration
0
10
20
30
40
50
60
70
80
90
100
0
20
40
60
80
100
120
140
160
180
0 5 10 15 20 25 % r
educ
tion
in d
srB
gen
e co
pies
ml-1
Dis
solv
ed su
lfide
in m
g m
l-1
Biocidal concentration mg ml-1
Dissolved sulfide in mg ml-1 % reduction in dsrB gene copies/ml
0102030405060708090100
020406080
100120140160180200
0 5 10 15 20 25 % re
duct
ion
in d
srB
gen
e co
pies
ml-1
Dis
solv
ed su
lfide
in m
g m
l-1
Biocidal concentration mg ml-1
Dissolved sulfide in mg ml-1 % reduction in dsrB gene copies/ml

Results 120
TERI university-Ph.D. Thesis, 2009
concentration of BNPD in 24 hours. This result showed that 20 ppm concentration of BNPD can control the growth of Desulfovibrio vulgaris effectively (fig. 5.32).
The South Kadi production water enriched cultures showed decrease of 94.17%, 97.92% and 98.07% in dsrB gene copies with biocidal concentration of 10, 15 and 20 mg.L-1 in 24 hours in respect to control. The dsrB gene copy number decreased from 36925 (control, 0 mg.L-1 biocide) to 715 copies ml-1 in biocidal concentration of 20 mg.L-1 (figure 5.33) number (MPN) method. The qrtPCR results showed that sulfide production stopped and there was a 99% reduction in dsrB gene copies at 20 mg.L-1biocidal concentration. No growth was observed, when this sample was re-inoculated in the new API RP38 growth medium. These results indicate that 20 mg.L-1of BNPD is optimum concentration to inhibit SRB in production water samples.
Quantification result showed nearly 10 folds high SRB when compared to most probable number (MPN) method. The qrtPCR results showed that sulfide production stopped and there was a 99% reduction in dsrB gene copies at 20 mg.L-1biocidal concentration. No growth was observed, when this sample was re-inoculated in the new API RP38 growth medium. These results indicate that 20 mg.L-1of BNPD is optimum concentration to inhibit SRB in production water samples.


















