
14-.-3'1' 15:3-3‘F-I3'; we: if. (Lihkl' :H'V
148
M, 55"] ”-5355... 5‘ 0"”. ‘5‘" "03“." (3" -n ‘v "' #355 . ‘5 3;"- ' 5‘" . El ' 5 51:, ‘HI' ' 5' 5"”: 1W -\'n . 15551975.- ‘ 31:33 ”3'45. qu ' '\‘ '36“ ‘ 3:- 33“ fix 3559335 5 (\‘yAL‘J35??? '53:}; ""14"- '1' .-3 15:3- "3" ‘F-I3'"; " we: Egg, :9 «533 V125" "‘ ‘ “35' ‘ ... 2‘ ,- ML"$53 " :55“? :31» 39:5“ '5“..% n}. 'l'_ u' I: f ’ . ' "i ‘ 5 5'” ‘ ‘gfxttg 454 I 5,-5’ L‘. S o . .5 ”3-3 - ' _ :_ Z '3 «‘3 t‘ ‘ “I; (‘55' :3“‘25 ”if , w:“73;? A.§&\§? {- éfitigf‘cufi} 3J1; v‘¢\ 4!“ '6'. ‘ v . ._ ‘ at. Y" ‘Q ‘ .3 . ;_.vf 5. 11‘.' 4 ' l. -y‘ 5:9:- 5 :\‘_J.'. ~. We“ ' J “5,3. - ' . o ‘ I ‘ - — .- J . ‘ . —~.— 4.. “*Wfl—sr- 5: 5. | 555.4 _;l I,“ ‘ 1‘ I." :LVJIE if. (Lihkl' :H'V
Transcript of 14-.-3'1' 15:3-3‘F-I3'; we: if. (Lihkl' :H'V

M, 55"]
”-5355...5‘0"”.‘5‘""03“."(3" -n ‘v
"' #355 . ‘5
3;"- '5‘"
.El
' 551:, ‘HI'
' 5' 5"”:
1W
-\'n .
15551975.-
‘ 31:33
”3'45. qu ' '\‘'36“
‘3:-33“fix35593355 (\‘yAL‘J35???'53:};
""14"-'1'.-315:3-"3"‘F-I3'";
" we:Egg,:9«533
V125" "‘ ‘“35' ‘ ...
2‘,-ML"$53":55“?:31»39:5“ '5“..%n}. 'l'_ u'
I:
f’
.'"i
‘
55'”
‘‘gfxttg
454
I
5,-5’
L‘.
So
..5
”3-3
-
'_:_
Z
'3
«‘3
t‘ ‘ “I;
(‘55'
:3“‘25 ”if ,
w:“73;?A.§&\§? {-éfitigf‘cufi}
3J1;v‘¢\4!“ '6'. ‘ v . ._ ‘at.Y" ‘Q ‘
.3 .
;_.vf
5.
11‘.'
4'l.
-y‘
5:9:
-5
:\‘_J.'.
~.
We“
'
J“5,3.
-'
.o
‘I
‘-
—
.-J
.‘
.—~.—4..“*Wfl—sr-
5:5. | 555.4
_;l I,“ ‘ 1‘ I.":LVJIE
if. (Lihkl' :H'V

WWHIWWII/Will”MM/WT“0064 5451
u ’5 - ‘ ~. £FW 0'4 smart-ve- '1
This is to certify that the
dissertation entitled
Short-term Growth of an Undisturbed
Tropical Moist Forest in the Brazilian
Amazon
presented by
Niro Higuchi
has been accepted towards fulfillment
ofthe requirements for
PhD degreein Forestry
55/
Major professor
Date {/1 31/6?
MSU i: an Alfmmm‘n Action/Equal Opportunity Institution042771

MSU
RETURNING MATERIALS:
Place in book drop to
remove this checkout from
LIBRARIES
All-(gulls.your record. FINES will
be charged if book is
returned after the date
stamped below.
”UN 1 1 19.0.0.
‘ 17:1"'~¢’
, 1'-
}J
r-
I‘l
I. v 1
I f '
l
f‘rfi E-‘k’.
“Anya “
55.
mt! 21 “3“}?
E . 2%55

SHORT-TERM GROWTH OF AN UNDISTURBED TROPICAL MOIST FOREST
IN THE BRAZILIAN AMAZON
BY
Niro Higuchi
A DISSERTATION
Submitted to
Michigan State University
in partial fulfillment of the requirements
for the degree of
DOCTOR OF PHILOSOPHY
Department of Forestry
1987

IHEHRACT
SHORT-TERM GROWTH or AN uuoxsruaaeo TROPICAL uorsr FOREST
IN THE BRAZILIAN amazon.
by
NIRO HIGUCHI
The main objective of this study is to provide basic
information for sustained yield management of the tropical
moist forest in the Brazilian Amazon. This was done by
quantification of diameter distributions, projections of
Idiameter distributions and of tree mortality, and by
development of short-term growth and yield models.
The data for this study were collected from an
undisturbed stand located some 90 kilometers north of
Manaus, the capital of Amazonas State - Brazil. Three
permanent plots were established in 1980 and remeasured in
1985. They are the control plots of a forest management
experiment randomly replicated within an area of about 2,000
hectares of pristine Amazonian forest. In each 4-hectare
plot (200 by 200 meters), all trees with dbh 25 cm or
greater were tagged and their dbhfls were recorded in 1980.
In 1985. all tagged trees were remeasured with special
attention to ingrowth and mortalityu The number of trees,
dbh and basal area of the study area averaged 158 trees/ha.
ii

38 cm, and 20 mZ/ha, respectively - in 1980.
Among three different hypothesized diameter
distribution functions, the Weibull using the percentile
approach best fit the observed data. 3
Since age and successive records from long-term
permanent plots were not available, the first-order Markov
chain approach was used to project diameter distribution and
tree mortality and proved to be a realistic alternative.
The exponential Lotka growth model was adapted to
predict future volume as an alternative for the traditional
growth and yield models, and it behaved satisfactorily. The
volume for 1990 was also predicted by models based upon the
volume estimated in 1985 in relation to the dbh measured in
1980. There was a strong correlation between actual volume
and past dbh, but not between past diameter and diameter
growth.
iii

To
Inezita and Chico, and Maria
my children, my wife - my friends
iv

ACKNOWLEDGEMENTS
I wish to express my gratitudetto Dr. Carl W. Ramm.
Chairman of my dissertation committee, for his insight,
support, and guidance in the preparation of this work. I
also wish to extend my gratitude to Dr. Lee M. James, Dr.
Kurt S. Pregitzer and Dr. Peter G. Murphy for serving on my
guidance committee and assisting throughout my doctoral
program.
I would like to extend my acknowledgements to Dr. Phu
Nguyen, Mr. T. W. W. Wood, Dr. Jurandyr C. Alencar, Dr. Kurt
S. Pregitzer, Dr. Lee M. James, and Dr. Carl W. Ramm. They
provided helpful suggestions on an earlier version of
specific chapters of this manuscript .
I would like to pay special tribute to my wife, my kids
and my "pessoal" from Chavantes and Chapeco for their
encouragement, patience and supportive "rezas".
I am indebted to many people whose friendship was
important during the course of this voluntary exile. Thank-
you to Antonio & Lucia, Josmar & Fernanda, Carlos, Steve
Westin, Robert De Geus, Bill Cole, Don Zak and George Host.
I would like to express my sincere gratitude to Luis
Maurc>& Fatima Magalhaes for being my proxy in Brazil and
for their patient support during this time.

Special thanks are due to the "peaozada" of DST
(Departamento de Silvicultura Tropical)-Aluizhm5Cabore,
Jesus, Barrao, Caroco, Paulista, Armando and other anonymous
helpers - who have been my great masters in the forest and
particularly for their help during field data collection. I
also wish to thank the group of DST's "pica-pan" - Fernando,
Antenor, Jurandyr, Magalhaes, Benedito, Noeli and Joaquim -
who played an important role during the preparation of this
research project. I am also indebted to many people from
other departments of INPA for their support. Thank-you to
"turma" of administration and to Nakamura.
I gratefully acknowledge the support of many staff
members of the Department of Forestry of Michigan State
University.
Finally, my sincere appreciation to my country - Brasil
- through CNPq (Conselho Nacional de Desenvolvimento
Cientifico e Tecnologi'co) for financial and administrative
support, and INPA (Instituto Nacional de Pesquisas da
Amazonia) for inspiration.
THANK YOU GOD !
vi

TITLE . .
ABSTRACT
DEDICATION
ACKNOWLEDG
TABLE OF C
TABLE
EMENTS O O O C
ONTENTS O O O 0
LIST OF TABLES . . . . .
LIST OF FI
CHAPTERS
1. INTRODU
Scepe
GURES O O O O
CTION O O O O O
of the Problem
OF CONTENTS
Statement of the Problem . . . .
2. LITERATURE REVIEW ON THE MANAGEMENT
REGENERATION IN THE TROPICAL MOIST FORESTS
2.1.
2.2.
2.3.
2.4.
2.5.
2.5.
2.7.
Overview . . .
Introduction . .
Tropical America
Tropical Africa
Tropical Asia .
Tropical South Pacific . . .
Conclusion . . .
vii
OF NATURAL
page
ii
iv
vii
xiii
03010101
12
13
16
16

3. THE BRAZILIAN AMAZON . . . . . . . . .
3.1. Introduction . . . . . . . . . . .
3.2. Climate . . . . . . . . . . . . .
3.3. Soils . . . . . . . . . . . . . .
3.4. Vegetation . . . . . . . . . . . .
Tropical moist forest on "terra firme"
Inundated forests . . . . . . . .
"Campina" and "Campinarana" . . .
Tropical semi-evergreen forest . .
"Cerrado" (Savannas) . . . . . . .
4. DESCRIPTION OF THE STUDY AREA . . . . . .
5. MODELLING THE DIAMETER DISTRIBUTION OF AN
UNDISTURBED FOREST STAND IN THE BRAZILIAN
AMAZON TROPICAL MOIST FOREST: WEIBULL VERSUS
EXPONENTIAL DISTRIBUTION . . . . . . . .
5.1. Introduction . . . . . . . . . . .
5.2. Procedures . . . . . . . . . . . .
The data . . . . . . . . . . . . .
The diameter distribution functions
The application of the functions .
5.3. Discussion of Results . . . . . .
5.4. Conclusion . . . . . . . . . . . .
6. A MARKOV CHAIN APPROACH TO PREDICT MORTALITY AND
DIAMETER DISTRIBUTION IN THE BRAZILIAN AMAZON .
6.1. Introduction . . . . . . . . . . .
6. 2. PrOCEdures O O O O O O O O O O O O
20
20
21
23
25
27
30
32
34
35
39
47
47
48
48
49
52
S3
55
68
71

The Data . . . . . . . . . . . . . . . . . 71
The Markov model . . . . . . . . . . . . . 71
The application of the model . . . . . . . 73
6.3. Discussion of Results . . . . . . . . . . 75
6.4. Conclusion . . . . . . . . . . . . . . . . 76
7. SHORT-TERM GROWTH OF UNDISTURBED BRAZILIAN
AMAZON TROPICAL MOIST FOREST OF "TERRA FIRME" . . 92
7.1. Introduction . . . . . . . . . . . . . . . 92
7.2. Procedures . . . . . . . . . . . . . . . . 93
The Data . . . . . . . . . . . . . . . . . 94
Model Development . . . . . . . . . . . . 95
7.3. Discussion of Results . . . . . . . . . . 97
7.4. Conclusion . . . . . . . . . . . . . . . . 102
8 0 CONCLUSIONS 0 O O O O O O O O O O O O O O I O O O 110
APPENDIX 0 O O O O O O O O O O O O O O O O O I O O O 113
L I ST OF REFERENCES 0 O O O C O O O O O O O O O O O 1 2 2
ix

LIST OF TABLES
Page
2.1. 1961 version of TSS - summary of operations . . 18
2.2. Malayan Uniform System (MUS) - summary of
activities . . . . . . . . . . . . . . . . . . 19
4.1. Listed species for the NR management
project . . . . . . . . . . . . . . . . . . . . 44
5.1. Diameter (cm) descriptive statistics for the
study area . . . . . . . . . . . . . . . . . . 56
5.2. Parameter estimates used for diameter
distribution - hectare basis . . . . . . . . . 57
5.3. Diameter distribution for all 3 sample plots
together (Bacia 3) derived from 3 different
methods . . . . . . . . . . . . . . . . . . . . 58
5.4. Diameter distribution for Bloco 1 derived from
3 different methods . . . . . . . . . . . . . . 59
5.5. Diameter distribution for Bloco 2 derived from
3 different methods . . . . . . . . . . . . . . 60
5.6. Diameter distribution for Bloco 4 derived from
3 different methods . . . . . . . . . . . . . . 61
6.1. Bloco 1 - Transition between states during a
5-year period . . . . . . . . . . . . . . . . 78
6.2. Bloco 2 - Transition between states during a
5-year period . . . . . . . .'. . . . . . . . 79
6.3. Bloco 4 - Transition between states during a

5-year period . . . . . . . . . . . . . . . . 80
6.4. Bloco 1 - Transition probability matrix from
one state to another during a 5-year period . . 81
6.5. Bloco 2 - Transition probability matrix from
one state to another during a S-year period . . 82
6.6. Bloco 4 - Transition probability matrix from
one state to another during a 5-year period . . 83
6.7. Bloco 1 Two-step transition probability
matrix . . . . . . . . . . . . . . . . . . . . 84
6.8. Bloco 2 - Two-step transition probability
matrix . . . . . . . . . . . . . . . . . . . . 85
6.9. Bloco 4 - Two-step transition probability
matrix . . . . . . . . . . . . . . . . . . . . 86
6.10. Bloco l - Projection for 1990 . . . . . . . . 87
6.11. Bloco 2 - Projection for 1990 . . . . . . . . 88
6.12. Bloco 4 - Projection for 1990 . . . . . . . . 89
6.13. Summary of one-step transition probability
matrix (1985) . . . . . . . . . . . . . . . . 90
6.14. Summary of two-step transition probability
matrix - projection for 1990 . . . . . . . . . 91
7.1. Basic distributional characteristics of the
data used for individual volume regression
equations . . . . . . . . . . . . . . . . . . . 104
7.2. Regression summary for volume estimation
models . . . . . . . . . . . . . . . . . . . . 105
7.3. Characteristics of the data used as yield
information and yield prediction . . . . . . . 106
xi

7.4. The frequency distribution of the three
dominant families by status in 1980, mortality
and ingrowth, and by periodic increment
classes in cm . . . . . . . . . . . . . . . . . 107
7.5. Regression summary for increment models . . . . 108
7.6. Mean, standard deviation, minimum and maximum
for each (a) dbh classes and (b) increment
Classes 0 O O O O O O O O O O O I O O O O O O O 109
xii

3.2.
4.1.
4.2.
5.1.
5.3.
5.5.
5.6.
LIST OF FIGURES
Index map for the Brazilian Amazon
vegetation map . . . . . . . . . . . .
The vegetation of Brazilian Amazon . .
"Ecological Management" Project area .
Bacia 3 with 4 experimental blocks . .
Bacia 3 - The relationship between the
observed and estimated dbh frequencies,
the Weibull MLE function . . . . . . .
Bacia 3 - The relationship between the
observed and estimated dbh frequencies,
the Weibull PERC function . . . . . . .
Bacia 3 - The relationship between the
observed and estimated dbh frequencies,
Exponential function . . . . . . . . .
Bloco l - The relationship between the observed
and estimated dbh frequencies by Exponential,
Weibull PERC, and Weibull MLE . . . . .
Bloco 2 - The relationship between the observed
and estimated dbh frequencies by Exponential,
Weibull PERC, and Weibull MLE . . . . .
Bloco 4 - The relationship between the observed
and estimated dbh frequencies by Exponential,
Weibull PERC, and Weibull MLE . . . . .
xiii
Page
37
38
45
46
62
63
64
65
66
67

CHAPTER 1
INTRODUCTION
During the past twenty years, the future of tropical
forests has been a matter of international concern.
Comprehensive reviews and evaluations are found in Gomez-
Pompa et a1. (1972), Budowski (1976a), Leslie (1977), Brunig
(1977), Spears (1979), Myers (1982), Myers (1983), Sedjo and
Clawson (1983), and Lanly (1983). The discussion is polemic
and most concerned scientists have been very pessimistic
about the future of tropical forests, especially tropical
moist forest (TMF). Nevertheless, there is one facet which
all the diverse approaches share to some extent: the TMF
ecosystem is very complex and fragile. Therefore, more
studies are required for a full understanding and definition
of its role for the region.
In terms of forest management of TMF, sustained yield
based on natural regeneration (NR) has been recommended by
most scientists. Nevertheless, success with this
silvicultural system is uncommon (Budowski 1976a).
While scientists and technicians are discussing the
problems of managing the tropical forest, about 20 hectares
per minute - an area equivalent to Puerto Rico per month -
of tropical forest are being deforestated, according to

Murphy (pers. comnh). Myers (1982) pointed out that the
principal causes of depauperation and depletion of TM? are
timber harvesting followed by slash-and-burn agriculture in
Southeast Asia, shifting cultivation in Africa, and cattle
ranching in Latin America.
In the Brazilian Amazon TMF, about 8 million hectares
(approximately 2% of the total area) have been deforestated
for the sake of agriculture and cattle ranching programs. By
the end of this century more than 2 million hectares will be
replaced by artificial lakes for energy generation. In
addition, areas open to mineral exploration have also
increased significantly.
In the face of increasing pressure on the definition of
role and vocation of the Brazilian Amazon TMF, in 1979 the
Federal Government made a commitment to develop a forest
policy for the region. All Amazonian research and
educational institutions were engaged to support this
policy. In the State of Amazonas two documents were produced
at the same time, one by the University of Amazonas (EUA
1979) and another by the National Institute for Research in
the Amazon (INPA 1979).
Inspired by the worldwide concern on the use of TM? and
forest policy, INPA initiated a research project. The
project, entitled "Ecological Management of Dry-land (terra-
firme)‘Tropical Moist Forest” was approved in 1979 by the
Brazilian Federal Government. It was financed by INPA, the
Interamerican Development Bank and FINEP (Brazilian

Financial Agency for Research). The main objective of this
project was to evaluate the impact of forest management
practices on the local environment. Basic ecosystem research
began in 1976 and the preparation for forest management
experiment effectively began in 1980.
This dissertation is based on observations of the
forest management area over a 5-year period. Only trees with
diameter at breast height (dbh) of 25 cm or greater were
observed. This study was conceived to provide biological
basis for sustained yield management based upon natural
regeneration development.
ScoEe of the Problem
The Ecological Management project is as important to
the Brazilian Amazon as the Hubbard Brook Ecosystem Study
has been to the mixed-species forest ecosystem of
Northeastern United States.
In the 2,000-hectare project area, two major research
studies have been carried out on ecology and forest
management. The areas for each study are referred to as
"Bacia 1" or "Bacia Modelo" and "Bacia 3", respectively for
ecology studies and forest management experimentation.
The initial results of "Bacia Modelo", including a
collection of basic ecology research results, were
documented by INPA in 1982 (INPA 1982).
"Bacia 3" is the area involved in this dissertation.

The results of this work will be used to help decision
makers in prescribing silvicultural treatments for an
experimental area subjected to a commercial timber
harvesting.
Statement 9; the Problem
The present study will investigate three separate
topics: quantification of diameter (dbh) distributions,
projections of dbh distributions and of tree mortality, and
development of short-term growth and yield models for
natural unmanaged Amazonian forest.
The specific objective of the diameter distribution
study is to find out which distribution function best fits
the observed data. Three hypothesized models were compared:
Weibull by percentile approach, Weibull by maximum
likelihood approach, and the exponential distribution
functions.
The second objective is to test the possibility of
using the first-order Markov chain approach to project
diameter distributions and to estimate tree mortality.
The third objective is to explore alternative ways to
model an undisturbed sample of TMF; Besides classical growth
and yield models, the Lotka's exponential model was tested.

CHAPTER 2
THE MANAGEMENT OF NATURAL REGENERATION IN THE TROPICAL
MOIST FORESTS.
2.1. OVERVIEW
This chapter reviews the management of tropical moist
forests (TMF) using natural regeneration, with or without
classical silvicultural systems. A diagnosis of the recent
situation of the application and research on natural
regeneration management, discussion of methods used in some
countries, and perspectives of sustained yield management
using natural regeneration are presented.
2.2. INTRODUCTION
There is no doubt of the importance of natural
regeneration for the management of TMF's. Very little is
known of the response of these forests when subjected to
intensive timber-oriented management used in temperate
regions (Cheah 1978, Tang 1980, and Rio 1976). Without
exception, all countries which contain TMF are still
considered as "developing" or "less developed" countries
(iJL, a mean GNP/capita about 10% of the North American
GNP). Another common characteristic of these countries is
the complex floristic composition of their predominantly

broadleaf evergreen forests.
Historically, natural regeneration management on a
sustained basis began with the Malayan Uniform System (MUS)
in Malaysia and the Tropical Shelterwood System (TSS) in
Nigeria (Fox 1976L.These two systems, modified and improved
with the passage of time and experience, have been used
extensively'in most tropical countries.lk>be meaningful,
natural regeneration management must be regarded as a
continuous process of silvicultural treatments to favor
economically desirable species. According to Rio (1979), the
objective of most treatments is the perpetuation of the
existing stands by the replacement of exploited forests
without a profound alteration of the characteristic
structure of the forest.
This review divides the tropical world into tropical
America, tropical Africa, tropical Asia, and the tropical
south Pacific. The current situation of natural regeneration
management and its perspectives are presented separately for
each region.‘The term tropical moist forest is based on the
Holdridge classification: biotemperature above 24° C and
annual precipitation between 2,000 and 4,000 mm.
2.3. TROPICAL AMERICA
According to Budowski (1976b), there is no example of
mixed TMF in the American tropics being managed on a
sustained yield basis.
In terms of research, however, Brazil (since 1980) and

Suriname (since 1967) have commenced studies to test the
possibility of TMF management on a sustained yield basis
using natural regeneration. Venezuela started a similar
project in the middle of the 19703, but no progress beyond
initial field establishment of the experiment and the
collection of pre-harvesting data was made. Recently Peru
also entered the natural regeneration management era. With
the assistance of British and Canadian technical aid
programs, the Honduran Forest Service will initiate studies
into sustained yield management of the TMF resources using
natural regeneration (Wood, pers. comnn). In Costa Rica,
sustained yield management has been planned for the Nosara
and Parrita river basins, with the assistance of FAO (Food
and Agriculture Organization) (Wood, pers. comm.). In
Dominica, between 1968 to 1972, an area of approximately 60
hectares was logged and planted with desirable tree species.
After about 3 years it was found that this operation was
very expensive to maintain, mainly due to the vigorous
growth of climbers. Therefore, the option with natural
regeneration management was considered (Bell 1976). In
Puerto Rico, a timber-management plan was completed in 1966
(Wadsworth 1970). This plan consisted of natural
regeneration treatments of 2,700 ha during the next 4
decades.
(a) BRAZIL
The concept of managing the native forests under a

system of sustained yield was introduced by FAO experts in
1958 in Santarem (State of Para) through an agreement with
the Brazilian Government. In Manaus (State of Amazonas),
researchers at INPA in 1964 initiated studies on enrichment
of natural forests, phenology'of tree species, and nursery
and plantation management of native and exotic species
(Higuchi 1981a).
In Santarem natural regeneration research was first
carried out fortuitously in 1960 when, after an area was
burned for species trial site preparation, copious
regeneration of Goupia glabra Aubl. appeared spontaneously.
This area is still under observation by researchers of
EMBRAPA (Brazilian Enterprise for Agricultural and Animal
Husbandry Research) and SUDAM (Superintendency of
Development of the Amazon region). Today, besides Goupia
glabra Aubl., species such as Vochysia maxima Ducke,
Didymopanax morototoni (Aubl.) Decne & P1anch., Manilkara
hubggi (Ducke) Standl., and Simaruba amara Aubl. are
abundant in an adjacent area.
Recent work with natural regeneration management in
Santarem is being carried out over blocks of 100 hectares.
The forest is harvested with diameter limits of 45 and 55 cm
dbh for commercial species after climber cutting and
underbrushing. The objectives of this project are to
determine the effects of different levels of harvest
intensity on the residual stand and regeneration, and to
evaluate the growth and yield under natural regeneration

management.
In Manaus the research with natural regeneration
management effectively started in 1980 under an agreement
among INPA (National Institute for Research in the Amazon),
the Interamerican Development Bank, and FINEP (Brazilian
Financial Agency for Research). The main objective of this
investigation was to test the possibility of managing the
TMF of the region under a system of natural regeneration. A
second objective was to use theidata to determine felling
cycles along with forecasts of yields by species. Within the
experimental blocks (400 by 600 meters), harvesting will be
carried out as the main silvicultural treatment. In
designated sub-blocks (200 by 200 m) felling intensities
will be applied to remove various levels of the basal area
of some 40 listed species, 25 cm dbh and above. This project
is based on multidisciplinary research involving all
departments of INPA (Ecology, Botany, Wood Technology,
Pathology, Agriculture, Chemistry and Zoology), which will
give scientific support to the Department of Tropical
Silviculture. The total area of this project is about 2,000
hectares while the area for silvicultural experimentation is
96 hectares, comprising 4 separate blocks of 24 hectares
each.
The treatments to be randomized in each block are: (1)
control; (2) removal of 25% of exploitable basal area
(b.a.); (3) removal of 50% of exploitable b.a.; (4) removal
of 75% of exploitable b.a.; (5) removal of 100% of

10
exploitable b.a.; and (6) removal of 50% of exploitable b.a.
with enrichment. In each 2 hectare plot a 1 hectare (100 by
100m) permanent sample plot will be established, in which
the following studies will be carried out: growth of the
residual stand of listed species; recruitment and
development of seedlings of the listed species; survival and
growth of listed species; growth and mortality of poles and
saplings; and studies of increment to determine felling
cycles.
(b) SURINAME
Research into the management of TMF resources was
initiated by the Suriname Forest Service in the 19508. The
Malayan Uniform System was used but it was discontinued in
the early 19603 due to the high costs of silvicultural
treatments, the long rotation (70-80 years), and the lack of
species with the silvicultural characteristics of the
dipterocarps of SE Asia.
The need for a management system suited to the
conditions of Suriname was met in 1967 under the auspices of
the CELOS (Center for Agriculture Research in Suriname). Its
objectives were to find an economically and technically
feasible method to stimulate the valuable timber species
increment after a light harvesting, to improve the
regeneration of the valuable species, and to build a forest
with sustained yield. Here, light harvesting meant the

11
removal of some 30 trees from the 25-ha experimental area.
Besides the classic silvicultural treatments, a refinement
was used wherein all non-valuable trees (non-commercial
species) were killed with arboricide (2,4,5-T ester, 5%
solution in diesel oil) using a‘diameter limit of 20 to 40
cm.
In this experiment the liberation treatments were: (1)
elimination of competing lianas and non-valuable trees
around the leading desirable tree selected on an area of 5
by 5 m; (2) elimination of competing species around the
desired species with a diameter criterion (3 to 5 cm dbh),
disregarding the location of the selected trees: and (3)
elimination of competing species around the desired species
in a strip 2 m wide, spaced 12.5 m apart, to provide
accessibility.
In the sampling area (16 ha), over 1,000 valuable trees
larger than 15 cm dbh are being measured yearly. Smaller
valuable trees are recorded in a 17.5% subsample using 40
circular plots of 1,000 sq.m each. As a provisional result,
de Graaf (1981) reported that the annual volume increment is
2.1 cu.m./ha for valuable trees above 15 cm dbh. According
to Johnson (1976), the mean annual growth of the TMF's is
about 1 to 3 cu.m./ha in South East Asia, 2 cu.m./ha in
Nigeria, and 2.9 to 4.3 cu.m./ha in the Philippines
(Dipterocarp forest). Even though there is not too much
detail in terms of tree size, in a general sense the forests
in Suriname are showing almost the same response to the

12
natural regeneration management as reported elsewhere.
2.4. TROPICAL AFRICA
According to Lowe (1978), the tropical shelterwood
system (TSS) was a major management preoccupation in Nigeria
during the 19503. Altogether about 200,000 ha of forest land
were treated under this system. It was intended to obtain
sustained or improved yields. The TSS consists of canopy
opening to promote survival and growth of seedlings of
valuable species. This system has been changed and improved
since its introduction, and the last version of TSS in 1961
is presented in Table 2.1.
However, TSS has been abandoned in Nigeria, primarily
on the economical grounds that it did not make sufficiently
intensive use of the land to compete with other forms of
land use (Lowe 1978). Nevertheless Rio (1976) pointed out
that economically, TSS is more profitable than plantations
if the analysis is correctly applied without bias. He
related that too often the forest management analyst seems
to survey the list of variables and select only those that
will contribute positively to the desired end. It seems
certain that silvicultural arguments did not contribute to
the abandonment of TSS in Nigeria.
In Ghana the TSS was tried on an experimental scale
between 1948 to 1960. It was found to be unsuitable because
of the high maintenance costs and was abandoned (Britwunn
1976). According to this author, the selection system was

13
found to be suitable for Ghana forests although it induced
only moderate regeneration. The treatments for this system
were: (a) stock survey to map all economic trees with dbh >
66 cm; (b) weeding, cutting and poisoning all climbers and
worthless trees which interfere with the development of
young economic trees (10 < dbh < 47 cm); and (c) selection
of trees to be felled from stock maps.
2.5. TROPICAL ASIA
A common characteristic in this region is the
significant presence of species of the Dipterocarpaceae.
This family contains the most important tropical hardwood
timber species. Other important species also occur in this
region, exp, teak (Tectona grandis) in Burma, teak and
Pinus merkusii in Thailand, Pinus kesiya in the Phillipines,
and Pinus merkusii in Indonesia.
According to Tang (1980), natural regeneration is the
basis for the regeneration of TMF in the region. The
silvicultural systems which have been developed for this
region are the Philippine Selective Logging System and the
Indonesian Selection Felling System for advanced growth, and
the Malayan Uniform System (MUS) and Indonesian modified MUS
- for seedling regeneration. Table 2.2 presents the sequence
of activities necessary for the MUS.
The MUS is, in fact, the most popular silvicultural
system in tropical Asia. It is mainly used in lowland

l4
Dipterocarp forests when.adequate reproduction.i3 already
established. There are restrictions in applying it in hill
forests where enrichment planting is often necessary (de
Graaf 1981L.In West Malaysia about 300,000 hectares have
been managed with MUS up to 1976.
Cheah (1978) discussed the differences between the new
selective felling system and the MUS or the modified MUS. He
determined that the first one is more appropriate for
dipterocarp forests in Peninsular Malaysia. The selective
felling system is a modification of the MUS, consisting of
the MUS plus the following operations: pre-felling inventory
which includes all trees below and above 15 cm, climber
cutting, and marking of residual trees for retention.
In Sarawak, the liberation thinning system was
introduced in 1975 by the Forest Department to evaluate the
effects of different intensities of reduction of stand basal
area as an alternative way to manage the natural
regeneration (Higuchi 1981b). This system seeks to eliminate
only trees which restrain the growth of a selected tree
(Hutchinson 1980). Modified MUS and removal of relics
(removal of all trees with dbh > 60 cm regardless of
species) has also been tested in Sarawak (Lee 1982).
In Sabah, the modified MUS was abandoned and replaced
in 1971 by the minimum girth system, which retains the basic
principles of the MUS (Chai and Udarbp 1977). This new
system includes three silvicultural treatments at three
different occasions. The first involves climber cutting two

15
years before felling operation to reduce the risks of
felling damage» The second combines the natural regeneration
inventory by linear sampling of milliacre plot and poison
girdling to eliminate competition. The third silvicultural
treatment involves the natural regeneration inventory by
linear sampling half-chain survey and a liberation
treatment. Chai and Udarbp (1977) concluded that the second
treatment should be modified to suit the present conditions
of logging in Sabah, and they recommended alternative
research to reduce logging damage.
In Indonesia, since 1972, the Indonesian selective
logging system has been used as a means of converting the
virgin forest into an enriched managed stand (Soekotjo and
Dickmann 1978). This system consists of removal of trees
with dbh > 50 cm to favor the growth of residual trees and
seedlings of desirable species. Approximately 25 young and
healthy overstory trees per hectare are usually left. After
4-5 years, the initial results have shown that the
Indonesian system seems to be appropriate for forest
management of Indonesian TMF (Soekotjo and Dickmann 1978).
In the Philippines, the selective logging system has
been used in managing the dipterocarp forests since the
19503” Specifically, this system assures a future crop of
timber and forest cover for the protection and conservation
of soil and water after the removal of the mature,
overmature and defective trees (Virtucio and Torres 1978).
According to these authors, the preliminary evaluation of

16
the selective logging has shown positive results for the
management of dipterocarp forests.
Other countries such as India, Burma and‘Thailand are
using the selective felling system to manage their forests
(James, pers. comm.). Burma contains 75% of world's stands
of natural teak. In India and Thailand, many species of
dipterocarp and teak are very important to the country's
forest economy.
2.6. TROPICAL SOUTH PACIFIC
Natural regeneration management was attempted in Fiji
during the 19603. Five years later this project was
abandoned (Higuchi 1981b) because the first results were not
encouraging. Today the priorities in Fiji are planting Pinus
caribaea var. hondurensis and management of Mahogony
(Swietenia macrophylla) plantations.
In Papua New Guinea, forest plantations seem to be the
only long-term alternative for its forests and for the
supply of its forest industries (Hilton and Johns 1984).
2.7. CONCLUSION
The utilization of natural regeneration as a tool for
forest management on a sustained yield basis in the TMF
mainly for dipterocarp forests is certain in almost all
southeast Asian countries. Although the Tropical Shelterwood
System (TSS) was abandoned in Nigeria, there exists a future

17
for natural regeneration as a way to manage the TMF, mainly
in well-stocked high forests (Kio 1976L.In South America
the first results of research recently established in
Suriname and Brazil have shown that natural regeneration
management is economically feasible and ecologically
acceptable.
The greatest obstacles to success with natural
regeneration management in tropical countries are the lack
of continuity in funding, the inadequacy of the staff, and
sometimes political factors. Tang (1980), for example,
considers that the success of natural regeneration
management depends on the implementation and monitoring
phases which can be carried out only with a well-trained
staff. According to Fox (1976), all mentioned problems are
typical in developing countries, primarily because the
anxiety to show progress is more important than anything
else. Unfortunately, natural regeneration management
requires long periods of time before results are known.
It is very important to maintain a cautious approach in
using the tropical moist forests because, according to Myers
(1983), very little is known about these ecosystems. It will
be better to find that we have been vaguely right than
certainly wrong.

18
Table 2.1: 1961 version of TSS - summary of operations.
YEAR INSTRUCTIONS
-5 Op.I Milliacre sampling
Op.Ia Demarcation
Op.II Climber cutting and cutting uneconomic
saplings if advance growth is inadequate
Op.III Climber cutting only
Op.IV 2nd. milliacre assessment following Op.II
0p.V Poisoning of shade casting trees of lower and
middle storeys
-4 (if Op.II in year -5, then Op.IV followed by 0p.V)
-2 0p.VI Re-demarcation
-1 0p.VII Climber cutting
0 Harvesting
8 Op.Ix Re-demarcation
Op.X Climber cutting
Op.XI Removal of Shelterwood
15 Op.XII Re-demarcation
Op.XIII 1/2 chain linear sampling
Source: partially reproduced from Lowe (1978).

19
Table 2.2: Malayan Uniform System (MUS) - Summary of
activities.
========================================================2=
ACTIVITY DESCRIPTION
Pre-Felling Except in cases where enumeration data
Inventory on trees 39 cm dbh and above is needed
for premium determination only.
Pre-Felling Treatment of bertam in hill forest only.
Treatment
Felling Limit All commercial and utilizable species
with dbh = 45 cm and above.
Tree Marking May or may not be done by forest
officers. Directional felling
incorporated but essentially for
checking completeness of felling only.
No marking of residuals for retention.
Roading Layout Prescribed specifications
and Construction
Post-Felling To determine fines on trees unfelled,
Inventory royalty on short logs and tops, damage
to residuals.
Silvicultural To determine correct treatment.
Sampling
Source: Cheah (1978).

CHAPTER 3
THE BRAZILIAN AMAZON
3.1. INTRODUCTION
The Amazon region includes the following countries in
South America: Brazil with 500 million (mi) hectares (ha),
Bolivia (65 mi.ha.), Colombia (62.5 mi.ha.), Peru (61
mi.ha.), Guyana (21.5 mi.ha.), Venezuela (17.5 mi.ha.),
Suriname (14.5 mi.ha.L, and French Guyana (9 mi.ha.)
(Volatron 1976). The name of this region comes from the
Amazon Basin and its main river, the Amazon, which
originates on Mt. Huagra in Peru at 5,182 meters above sea
level (a.3.l), 195 km from the Pacific shore. According to
Palmer (1977) in the first 965 km from its source, the
Amazon river drops 4,876 m while in the remaining 5,785 km
the fall to sea level is only 306 m.
In the Brazilian territory the area of influence of the
Amazon Basin includes the following regions: Acre (AC),
Rondonia (RO), Amazonas (AM) and Para (PA) states, part of
Mato Grosso (MT), Goias (GO) and Maranhao (MA) states, and
two federal territories, Roraima (RR) and Amapa (AP).
Hereafter, this area will be referred to as the Brazilian
Amazon or simply as the Amazon. This area is under
geographical and political influence of Amazon Basin, even
though it is known that the Amazon forest ecosystem covers
20

21
around 3/5 of this area. The Amazon region corresponds to
about 55% of the Brazilian territory, but its population
represents only 10% of its total. Fig. 3.1 shows the
location of Brazilian Amazon within South America.
3.2. CLIMATE
The Brazilian Amazon region is characterized by
homogeneity in climate conditions. In the interior of the
forest of this region the microclimate is much more equable,
especially on the ground itself where no direct sunlight
falls (Walter 1979). Coastal and in-land temperatures do not
differ greatly. Belem, some 100 km from the sea, has a mean
annual temperature of 25° C. Manaus, nearly 1,000 km up-
river on the Amazon, has an equivalent of 27° C and Taraqua
some 2,000 km in-land has a mean annual temperature of
24.9° C. The maximum temperatures are around 37 to 40° C
with a diurnal variation of some 10 degrees. According to
Salati & Vose’(1984), however, an important phenomenon to be
considered is the "friagem' or cold spells that occur when
air masses from the South Polar region hit the central and
western parts of Amazon, causing the temperature to fall to
about 14° C.‘This phenomenon occurs during the winter in
the states of Acre and Rondonia, and in the southern parts
of Amazonas state.
Rainfall shows greater variability than temperature
across the region. There is approximately 3,000 mm annual
rainfall on the coast, 3,497 mm at Taraqua (less than 100 km

22
from the limit boundary between Brazil and Colombia), 1,504
mm in Boa Vista (the Capital of Roraima), and 1,670 mm in
Conceicao do Araguaia.
According to Ranzani (1979) the dominant climatic types
(Koppen classification) in the region are Af (coolest month
above 18° C and constantly moist) and Aw (coolest month
above 18° C and dry period during the winter).
Including air moisture regime (presence of dry period
with its duration), IBGE (1977) identified five climatic
zones:
(a) Equatorial very moist without dry period: covers
the northwest Amazon (about 30% of Amazonas state) and Belem
(the Capital of Para state).
(b).Equatorial very moist with short dry period.(less
than one month): covers the surrounding areas of type (a)
(about 30% AM and 25% of AC).
(c) Equatorial moist with dry period ( one to two
months): covers the western-center and the southeast of
4Amazon (50% of AC, 30% of AM, 30% of RR, 30% of PAmand 10%
of north of MT).
(d) Equatorial moist with dry period (three months):
covers the southwest and the eastern-center of Amazon (10%
of AM, 100% of RD, 70% of PA, 40% of RR, 70% of AP, 10% of
G0, 40% of MT and 40% of MA).
(e) Tropical semi-moist with dry period (four to five
months): covers part of RR and south and southeast of Amazon
(30% of RR, 50% of MT, 90% of Goland 60% of MA).

23
3.3. SOILS
The soils in the Brazilian Amazon are very old,
reaching back as far as the Paleozoic era. Basically the
region is composed of a sedimentary basin (Amazon Valley)
located between two shields (Guiana and Brazilian).
According to IBGE (1977) these two shields are composed of
igneous Precambrian and metamorphic rocks from Cambrian-
Ordovician, They contain some spots of sediments from the
Paleozoic/Mesozoic (60 to 400 million years ago). There are
two Paleozoic strips of sediments where Devonian shales
predominate, one at the Guiana shield boundaries (east of
the 60 degrees of longitude) and another at the Brazilian
shield boundaries (east of the 57 degrees of longitude) 30
to 50 km wide (Schubart & Salati 1980). The Amazon Valley is
formed by fluvial sediments of coarse texture deposited from
the Cretaceous to the Tertiary periods, originated from the
erosion of the Precambrian shields (Schubart & Salati 1980).
In summary, this is the evolutive process of formation of
”terra firme" (non-flooded ground).
Another important formation in the‘Amazon region is the
"varzea", or temporarily flooded land. According to Schubart
5 Salati (1980) the ”varzeas" are constituted by the
Holocene flood plains of the Solimoes river (Amazon river
above Manaus) and.the Amazon as well as their white water
tributaries. "Varzeas" are the most recent formation in

24
from the deposition of sediments transported by the rivers
(Ranzani 1979). This kind of formation represents only 1.5%
of the region, but its high agriculture productivity is
significant to the Amazon economy. Ranzani (1979) pointed
out that its fertility is not constant, as it depends upon
the materials incorporated annually by flooding.
According to Cochrane & Sanchez (1980) the following
soil orders are found in the Brazilian Amazon: Oxisol
"yellow Latosols" (45.5%), Ultisols "red yellow Podzolics"
(29.4%), Entisols "azonal, alluvial soils" (14.9%), Alfisols
"gray brown Podzolics" (4.1%), Inceptisols "hydromorphics,
humic gley soils" (3.3%), Spodosols "Podsols or giant
tropical Podzols" (2.2%), Mollisols "Chernozem, humic gley
soils" (0.8%), and Vertisols "grumusols" (0.1%).
In general, the soils are extremely poor in nutrients
and very acid. In fact, almost the entire nutrients amounts
required by the forest are contained in the aboveground
biomass (Walter 1979). Cochrane 3 Sanchez (1980) pointed out
that only about 6% of Amazon has well drained soils with
relatively high natural fertility. These soils are found in
Altamira (Para state), Porto Velho (the capital of Rondonia
state) and Rio Branco (the capital of Acre).
Ranzani (1979) stressed that few Amazon soils are
suitable for agriculture, grazing or even for reforestation.

25
3.4. VEGETATION
Using the Holdridge classification and the
climatological observations of IBGE (1977), there are two
major life zones in the Brazilian Amazon. These are the
tropical moist forest (mean annual biotemperature above 24°
C and mean annual precipitation of 2,000 to 4,000 mm) and
the tropical dry forest (mean annual biotemperature above
24° C and mean annual precipitation of 1,000 to 2,000 mm).
According to Schubart & Salati (1980) about 8% of the
Amazon is under secondary vegetation and/or agricultural
activitiesu‘Within the tropical moist forest only limited
areas on the coast, along the major tributaries of the
.Amazon and along the Amazon River have been used for food
production (Tosi 1983). The most significant deforestation
is located in the tropical dry forest, mainly along the
Belem-Brasilia highway, southern portions of MT, and in
Rondonia and Acre states.
It is well known that the main characteristic of the
.Amazon forest is its considerable vegetational diversity,
although at first sight it appears to be rather uniform
(France 1974).
The Amazon region is reported to contain about 6,000
different species of plants, of which one-third are tree
species growing to commercial size. The distribution of
these trees varies tremendously, particularly in relation to
soils and topography.
There are many theories to explain this diversity.

26
According in: Prance (1974) the genetic isolation into
separate populations after a long dry period in the late
Pleistocene and post-Pleistocene was a major factor in the
evolution of the species diversity within the lowland forest
of Amazon. Schubart & Salati (1980) pointed out that the
large number of species and the complexity of their
interrelatioships are a function of evolutionary history
which can be broadly described by three main categories of
factors: proximal (or geographic factors), interactions
within.the communities themselves, andtdynamic instability.
In spite the complexity and diversity of the Amazon
vegetation, a broad classification - based on the Holdridge
system plus part of the classification presented by Prance
(1974) - will be presented for the two major life zones
(Fig. 3.2).
1. Tropical moist forest
1.1. Tropical moist forest on "terra firme"
1.2. Inundated forests: "varzea" (seasonally flooded
forest) and "igapo" (permanently water-logged)
lu3. Forest on white sand soils or spodosols: "Campina"
and "Campina r ana" .
2. Tropical dry forest
2.1. Amazon tropical semi-evergreen forest
2.2. "Cerrado" (Savannas).

27
Tropical moist forest on "terra firme”
The superior stratum of this forest type is composed of
trees whose heights may vary from 30 to 40 meters. Only a
few species can grow above this height. Exceptions are
Cedrelinga catenaeformis and Dinizia excelsa with, on some
sites, 50 and 60 meters height respectively. For trees with
dbh greater than 20 cm, the forest on "terra firme" has a
mean commercial volume of 150 to 300 cu.m./ha and a basal
area of 20 to 40 sq.m./ha.
IBGE (1977), Braga (1979), Silva et a1. (1977), Higuchi
et al. (1983a), and four forest inventories carried out by
Department of Tropical Silviculture of INPA (National
Institute for Research in the Amazon) in different parts of
Amazon are the guide for the description of floristic
composition of this type of forest. Here the emphasis is
only (”1 those species which can characterize specific
regions.
In a broad sense the following phanerophytes can be
considered as typical species of "terra firme": Dinizia
excelsa, Bowdichia nitida and Cedrelinga catenaeformis
(Leguminosae), Anacardiug gigagtggm (Anacardiaceae),
Bertholletia excelsa "Brazilian nut" (Lecythidaceae),
Caryocar villosum (Caryocaraceae), Minquartia guianensis
(Olacaceae), and two species of Palmae, Oenocarpus bacaba
and Astrocaryum mumbaca. The characteristic epiphytes of
”terra firme" are: several species of Phillodendron
(Araceae), Clusia insignis and Clusia grandiflora

28
(Guttiferae), several species of Operculina (Convolvulaceae)
and Bauhinia macrostachya (Leguminosae).
IBGE (1977) divided the ”terra firme" into seven sub-
regions to show the characteristic tree species of these
areas, in contrast to the previous group of species which is
common to all sub-regions.
The sub-regions are:
(a) Delta of Amazon river: In this area the following
species characterize the "terra firme": several species of
Parkia, Vatairea guianensis and several species of Ormosia -
(Leguminosae), Erisma fuscum and Vochysia guianensis -
(Vochysiaceae), several species of Manilkara and Pradosia -
(Sapotaceae), and several species of Virola -
(Myristicaceae).
(b) Northeast Amazon: several species of Micropholis,
Ecclinusa, Chrysophyllum and Manilkara - (Sapotaceae),
several species of Eperuaz Swartzia, Ormosia, Tachigalia and
Inga - (Leguminosae), Goupia glabra (Celastraceae), several
species of Iryanthera - (Myristicaceae), and several species
of Qualea - (Vochysiaceae).
(c) Tocantins & Gurupi rivers: Swietenia macrophylla
"Mahogany", Cedrela odorata and 935323 guianensis -
(Meliaceae), Hevea brasiliensis - (Euphorbiaceae),
Platymiscium duckei, Vouacapoua americana, and several
species of Piptadenia and Peltogyne - (Leguminosae), Cordia

29
goeldiana - (Boraginaceae), Mezilaurus itauba - (Lauraceae),
several species of Astronium - (Anacardiaceae), and
Jacaranda copaia - (Bignoniaceae).
(d) Xingu and Tapajos rivers: The floristic composition
of this sub-region is almost the same as the sub-region (c).
(e) Madeira and Purus rivers: Hymenolobium excelsum,
Peltogyne densiflora, several species of E2353; and
Elizabetha - (Leguminosae) Swietenia macrophylla and Carapa
guianensis - (Meliaceae), Euterpe oleracea - (Palmae),
several species of Theobroma - (Sterculiaceae), Cordia
goeldiana - (Boraginaceae), Manilkara huberi - (Sapotaceae),
Cariniana micrantha - (Lecythidaceae), Hevea brasiliensis.
(f) Occidental "Hileia" - Jurua to Brazilian territory
limits: several species of Theobroma "Cocoa tree" and
numerous palms, and several species of Leguminosae,
Myristicaceae, Bombacaceae, Lauraceae, Vochysiaceae and
Rubiaceae.
(9) Northwestern "Hileia" - Negro to Trombetas river:
Leguminosae is the dominant botanical family in this sub-
region, mainly species of genera Dimorphandra, Peltogyne,
Eperua, Heterostomon and Elizabetha. The genena Dicorynia,
Aldina, Macrolobium and Swartzia are endemic:in this sub-
region. Other characteristic species are: Carapa guianensis,
Cedrela odorata and Cariniana micrantha.

30
(h) Acre: Torresea acreana - (Leguminosae), Hevea
brasiliensis, Swietenia macrophylla and several species of
Cedrela.
Inundated forests
This type of forest represents an area of about 7
million hectares, or 1.5% of the Amazon region (Braga 1979).
Within this type, the best and the biggest portions are the
seasonal "varzea" and tidal "varzea". They are considered
very important for the development of the Amazon region
because of their soil quality and also because they supply
most of the raw material to forest industries.
Prance's (1980) key for the classification of inundated
forest types was used to describe the vegetation. The author
pointed out that the three different types of water (white,
black and clear) of Amazon basin are very important to the
floristic composition. There are peculiar species for
specific water types, mainly due to differences in acidity
and nutrient contents. For example, Victoria amazonica is
found only in white water.
The seven inundated forest types are:
(a) Seasonal ”varzea": this type is characterized by a
relatively high aboveground biomass and represents the most
common type of inundated forests. According to Prance (1980)
its herb layer is rich in species of Heliconia (Musaceae)
and Costus (Zingiberaceae). The following species can

31
characterize this type of forest.cPrance 1980, Braga 1979,
and IBGE 1977): Carapa guianensis, several species of
Cecropia - (Moraceae), Ceiba petandra - (Bombacaceae),
Couroupita subsessilis - (Lecythidaceae), Euterpe oleracea -
(Palmae), Hura crepitans and Piranhea trifoliata -
(Euphorbiaceae).
(b) Seasonal "igapo" - swamp forest: Usually dominated
by sand soils supporting a vegetation much poorer than the
seasonal "varzea". According to Braga (1979), the vegetation
is very specialized with little specific diversity and very
rich in endemism. Characteristic species of this type are:
Aldina latifolia - (Leguminosae), several species of Couepia
- (Lecythidaceae), some species of Licania -
(Chrysobalanaceae), and Macrolobium acaciifolium -
(Leguminosae).
(c) Mangrove: This type is typical in the estuary of
the Amazon. According to Braga (1979) the mangrove type
involves an area of about 100,000 hectares with a low and
uniform aboveground biomass. This type is characterized by
the presence of Avicennia nitida (Verbenaceae), Laguncularia
EEEEEQEE (Combretaceae) and BEEEQREQEE ‘1‘. “91.2.
(Rhizophoraceae).
(d) Tidal "varzea": This type is very similar to the
seasonal "varzea" in both species composition and
aboveground biomass. Prance (1980) stressed that where the

32
tide is daily, the vegetation is similar to the swamp. Where
the spring tide is dominant, is more similar to the seasonal
"varzea". The most common palm species are: Mauritia
flexuosa, Euterpe oleracea, Raphia taedigera and Manicaria
saccifera. Species like Virola surinamensis (Myristicaceae),
Ceiba petandra, Mora paraensis, Pithecolobium huberi, Derris
liEiEQliiL EXEEBEEE 222222 and lflflé BEEEQQBE '
(Leguminosae), and Tabebuia aquatilis (Bignoniaceae) have
also a significant presence in this type of forest.
(e) Flood plain: Species from seasonal "varzea" and
also from "terra firme" can be found in this forest type.
(f) Permanent swamp forest: According to Prance (1980)
there are few permanent swamp forests or permanent ”igapo”
in the Amazon. This type contains very few species,
although trees are usually very big and similar to their
counterparts of seasonal "varzeaF. The canopy is usually
more open than the seasonal "varzea" and the ground is rich
in Cyperaceae.
"Campina" and "Campinarana"
The soil of these two types is almost the same, but
their floristic composition and the stand density are
different. According to Lisboa (1975), the tropical moist
forest on "terra firme" is commonly interrupted by ”islands”
with contrasting tree size, structure and physiognomy. Such
”islands” are oommon.in the Rio Negro river basin and in

33
other areas north of the Amazon river, but almost absent in
the southern parts of this river. "Campina" and
"Campinarana" are evergreen.
(a) "Campina":‘According to Braga (1979), this forest
type presents a low aboveground biomass with sclerotic
vegetation, and covers an area of 3.4 million hectares (0.7%
of Amazon).
Although the "Campina" soils are excessively drained,
acid and poor in nutrients, there is no problem with water
availabilityu Lisboa (1975) pointed out that without this
characteristic the actual vegetation could be replaced by
Gramineae, Cyperaceae and small shrubs.
Thee"Campina" floristic composition is variable, but
the following species could be considered as characteristic
species of this forest type (Braga 1979): Aldina
heteroghylla and Ormosia costulata - (Leguminosae), Clusia
aff. columnaris (Clusiaceae), Glycoxylon inophyllum
(Sapotaceae), Humiria balsamifera (Humiriaceae), Matayba
923 a (Sapindaceae), and Protium heptaphyllum (Burseraceae).
According to Lisboa (1975) the epiphytes are abundant in
"Campina" because the high intensity of light, e.g., many
genera of Orchidaceae (Sauticaria, Octomeria, Rodrfigzia
and Maxillaria) and also many species of Bromeliaceae
(Aechmea and Tillandsia).
(b) "Campinarana" (false "Campina"): In this forest
type the trees are larger and the stands are denser in

34
comparison to the "Campina" type. According to Braga (1979),
"Campinarana" represents an area of approximately 3 million
hectares distributed as small islands in the central Amazon
and as bigger portions north of Amazon river (Negro basin).
"Campinarana" is also very rich in epiphytes, mainly
Hymenophyllaceae and Bryophytae. The following species
characterize this forest type (Braga 1979): Aldina discolor,
Eperua leucantha and Hymenolobium nitidum - (Leguminosae),
Bactris cuspidata (Palmae), Clusia. spathulaefolia
(Clusiaceae), Qggm§_ gatigga£_ (Apocynaceae), ‘ggggg
rigidifolLa (Euphorbiaceae), Sacoglottis heterocarpa
(Humiriaceae) and Scleronema spruceanum (Bombacaceae).
Amazon tropical semi-evergreen forest
This forest is considered as a transition from Savannas
and tropical semi-evergreen to tropical moist forests. It
occurs in part of MA, portions of eastern, southern and
northern PA, northern MT, almost 90% of Rondonia, portions
of AC, small portions at northern and southern AM, a
significant portion of the federal territory of Roraima and
a small portion of Amapa.
In general, according to IBGE (1977), the trees are
relatively tall, with medium diameter and under-developed
crowns. Lianas are abundant, but epiphytes are almost
absent. The species most characteristic of this forest type
is Orbignya martiaga (Palmae). Hevea brasiliensis is
abundant mainly along the southern tributaries of the Amazon

35
river.
In the MA portions and eastern PA the species which
characterize this forest type are: Bertholletia excelsa,
Ceiba petandra, Vouacapoua americana, Castilloa ulei
(Moraceae), Hymenaea courbaril (Leguminosae), Lecythis
paraensis (Lecythidaceae), and several species of Palmae,
e.g., Oenocarpus bacaba, Maximiliana £2933 and Euterpe
oleracea.
According to IBGE (1977) the best known portion of
Amazon tropical semi-evergreen forest is that in the
southern part of PA which partially covers the Brazilian
shield. The characteristic species are: Calophyllum
Riééilififlfifi (Guttiferae), some species of £9222;
Aspidosperma and Moutabea, Apuleia praecoxi Hymenaea
stilbocarpa, Lucuna lasiocarpa, Simaruba amara, etc.
At the eastern of'the Tapajos river, between Santarem
and Belterra, and the northern of the Amazon river, the
northern part of PA, the following species are
characteristic: Qualea grandiflora and Vochysia ferruginea -
(Vochysiaceae), Sclerolobium paniculatum, Dalbergia
spruceana and Centrosema venosum - (Leguminosae).
"Cerrado" (Savannas)
The "Cerrado" trees are relatively short (around 10
meter height) and less abundant than shrubs. Basically there
are two strata: the superior which is composed of trees and

36
shrubs, and the inferior which is composed of grasses. The
tree stratum is characterized by individuals with crooked
stem and branches, thick bark, and thick leaves with rough
grained texture with surfaces of 30 by 20 cm.
According to IBGE (1977), the characteristic species of
"Cerrado" are: Hancornia speciosa (Apocynaceae), Curatella
americana (Dilleniaceae), Qagyggar brasiliensis
(Caryocaraceae), Salvertia convallariaedora, Kielmeyera
coriacea, and Stryphnodendron barbatimao.

37
4
k}?
Ii
'5 vat
a
3
4"s ‘
’~ 1
m {_
al'
wwwvra-wwdarww
w
D! - Federal District: Brasilia

38
EEEI
-
CI]
‘gy/
-
‘TIIIA 'llll”
llNl-IVIRCIIIW VOIISY
SAVAIIA - ‘CIIIAUO"
CIASSLAIDS
'VAIIIA” OI "lfihln"
COASYAL VIC'YATIBI
CD'PLIIII or IBIAIIA. FACIIIIO AID lllflu
'Loollfl GIASSLAIOI
COI'LII ur PAIYAWAL

CHAPTER 4
DESCRIPTION OF THE STUDY AREA.
The data were collected on the control plots of an
experiment on natural regeneration management of an uneven-
aged mixed stand of the Amazonian forest. This experiment is
being carried out by DST (Department of Tropical
Silviculture) of INPA.(Nationa1 Institute for Research in
the Amazon). The experiment is a branch of the project
"Ecological Management of the Dry-land Tropical Moist
Forest". This multidisciplinary research involves all
departments of INPA: Ecology, Botany, Wood Technology, Plant
and Human Patology, Agriculture, Chemistry and Zoology.
These departments will give scientific support to DST in its
future evaluations of the environmental impact of the forest
management.
The study area is located within the domain of the
Tropical Silviculture Experimental Station of INPA, some 90
kilometers north of Manaus, the capital of Amazonas State,
Brazil. The total area of the Station is 23,000 hectares and
the project area is approximately 2,000 hectares. The
geographical coordinates of the project area are 2° 37' to
2° 38' of south Latitude and 50° 09' to 60° 11' of west
longitude. Figure 4.1 shows the location of the study area
39

40
within the Experimental Station.
According to Ranzani (1980) the climate is type Am,
Koppen classification, warm and moist all year long. The
annual rainfall is approximately 2,000 mm without
accentuated dry period, even though the wettest period is
December to May (Ribeiro 1977).
The oxisol soil order "yellow latosols" is predominant
in the area. This research was set up only on non-flooded
ground, iue., on "terra firme". The soils are extremely poor
in nutrients and very acid.
The relief is smoothly undulated and it is formed by
small plateaus which vary from 500 to 1,000 m in diameter.
Most of the experimental treatment areas are located on
those plateaus.
The vegetation is typical of the Amazonian tropical
moist forest on "terra firme". The superior stratum of this
forest is composed of trees whose heights vary from 30 to 40
meters. Basically three botanical families dominate the
floristic composition of the area, Lecythidaceae,
Leguminoseae and Sapotaceae. Individually Micrandropsis
scleroxylon W.Rodr. (Euphorbiaceae) and Scleronema
micranthum Ducke (Bombacaceae) have an impressive presence
in the study area. Several species of Eschweilera ,
Holopyxidium latifolium R. Knuth, Corytophora alta R. Knuth
and Lecythis usitata Miers var. paraensis R. Knuth are the
most frequent species of Lecythidaceae. However,
Bertholletia excelsa Humb. and Bonpl. "Brazilian nut”

41
(Lecythidaceae) is absent from the area. The most frequent
Leguminosae are several species of Inga, Tachigalia,
Swartzia, Parkia and Pithecolobium. Within the Sapotaceae
the most frequent are several species of Chrysophyllum,
Micropholis, Pouteria, Labatia, Ecclinusa, and Manilkara.
The floristic composition of the area is presented in the
Appendix.
The ecological project area is in the Tarumazinho
watershed. The project was divided into three parts,
referred to as bacia l, bacia 2 and bacia 3. Respectively,
these are areas reserved for basic studies, buffer, and
harvesting and forest management.
Bacia 3 is the basis of this study. Figure 4.2 shows
BaciaLB in more detail, Originally this experimental area
covered 96 hectares, consisting of 4 blocks (bloco l, bloco
2, bloco 3, and bloco 4) of 24 ha each. After the commercial
inventory, bloco 3 was reserved for research on artificial
regeneration and, therefore, it was not included in this
study. Within each block (400 by 600 m), harvesting will be
carried out as the main silvicultural treatment. In
designated sub-blocks (200 by 200 m each), different felling
intensities will be applied to reduce basal area of some 40
listed species with dbh ; 25 cm.
The treatments randomly distributed in each block were:
(1) control, (2) removal of 25% of the exploitable basal
area (b.a.), (3) removal of 50% of the exploitable b.a., (4)
removal of 75% of the exploitable b.a., (5) removal of 100%

42
of the exploitable tha., and (6) removal of 50% of the
exploitable txa. with enrichment. In each four-ha sub-block
a one-ha plot (100 by 100 m) was established to evaluate the
growth of the residual stand of listed species, recruitment
and development of seedlings of listed species, survival and
growth of listed species, growth and mortality of poles and
saplings, and increment evaluation for determining the
felling cycles. The listed species for this project are
presented in Table 4.1.
After the randomization of the blocks, the control sub-
blocks were 2, 3 and 5, respectively for blocks 1, 2 and 4.
Those sub-blocks, then, were used in this study. Hereafter
they will be referred to as bloco l, bloco 2, and bloco 4,
and collectively they will be called bacia 3.
In 1980, two different inventories were carried out in
bacia 3: commercial (complete enumeration of trees with dbh
> 25 cm within the experimental blocks), and diagnosis of
natural regeneration by sampling.
From the commercial inventory (Higuchi et al. 1983a)
the following data were obtained: (a) the listed species
represent 1/3 of the population, (b) overall means per ha:
number of trees = 155, b.a. = 19 sq.m., and volume with bark
= 190 qum" (c) block 3 is statistically different from the
others in terms of stand stocking and also in terms of
floristic composition.
From the natural regeneration inventory (Higuchi et a1.
1985) the following summaries were obtained: (a) the

43
stocking index of seedlings averaged 15.6%, (b) the stocking
index of poles and saplings averaged 72.8%, and (c) the
number of trees smaller than 25 cm dbh and greater than 0.30
m height averaged about 40,000 per hectare. The "milliacre”
and "half chain square" methods were used for data
collection of the diagnostic inventory, respectively for
seedlings (tree species with dbh < 5 cm) and for poles and
saplings (5 < dbh < 25 cm).
In 1985, all trees tagged in 1980 from the control
plots were remeasured. This was done to evaluate the growth
of diameter of those trees (increment), to record new trees
that moved to the first merchantable dbh class (ingrowth),
and to record trees which died during the period 1980-1985
(mortality).

44
Table 4.1: Listed species for the NR management project.
Spec1e Family
Virola calophylla Warb. Myristicaceae
Virola multinervia Ducke Myristicaceae
Virola venosa (Bth. ) Warb. Myristicaceae
Ocotea cymbarum H. B. K. Lauraceae
Dialium guianensis (Aubl. ) Sandw.
And1ra micrantha Ducke
D1plotropis purpurea (Rich. ) Amsh.
Manilkara huberi (Ducke) Standl.
Calophyllum angulare A. C. Smith
Nectandra rubra (Mez.) C.K. Allen
Mezilaurus synandra (Miq.) Kostermans
Licaria guianensis Aublet.
Platymiscium duckei Huber
Caryocar villosum (Aubl.) Pers.
Goupia glabra Aubl.
Aniba duckei Kostermans
Naucleopsis caloneura (Hub.) Ducke
Scleronema micrantha Ducke
Minquartia guianensis Aubl.
Copaifera multijuga Hayne
Qualea paraensis Ducke
Diniz1a excelsa Ducke
P1thecolobium racemosum Ducke
Hymenolobium excelsumDucke
Astronium lecointe1 Ducke
Clarisia racemosa R. et P.
Hymenaea courbaril L.
Dipteryx odorata (Aubl.) Willd.
Lecyth1s usitata Miers
S1maruba amara Aubl.
Caryocar pallidum A. C. Smith
Erisma fuscum Ducke
Holopyxidium latifolium R. Knuth
Vouacapoua pallidior Ducke
Eschweilera odora (Poepp) Miers
Eschweilera longipes (Poit) Miers
Anacardium spruceanum Benth. ex Engl.
Aniba canellila (H.B.K. ) Mez.
Park1a pendula Benth. ex Walp.
Corythofora r1mosa Rodr.
Cariniana micrantha Ducke
Cedrelinga catenaeformis Ducke
Peltogyne catingae M. F._da Silva
Bros1mum rubescens Taub.
Leg. Papil.
Leg. Papil.
Leg. Papil.
Sapotaceae
Guttiferae
Lauraceae
Lauraceae
Lauraceae
Leg. Papil.
Caryocaraceae
Calastraceae
Lauraceae
Moraceae
Bombacaceae
Olacaceae
Leg. Caesalp.
Vochysiaceae
Leg. Mimos.
Leg. Mimos.
Leg. Papil.
Anacardiaceae
Moraceae
Leg. Caesalp.
Leg. Papil.
Lecythidaceae
Simarubaceae
Caryocaraceae
Vochysiaceae
Lecythidaceae
Leg. Caesalp.
Lecythidaceae
Lecythidaceae
Anacardiaceae
Lauraceae
Leg. Mimos.
Lecythidaceae
Lecythidaceae
Leg. Mimos.
Leg. Caesalp.
Moraceae

4S
@- 31w. 3 - :xmagfio E MANEJO
[D — A'REA ssnrjo EXPERIMENTAL DE smvncuururu TROPIC
I
AL
@ - AREA nrsznw Badman a: CAMPINA "MI
lB.
‘
/ ‘1.V l, , crpuc 3
~\ . '
5, ___-
, I- 7 a) I
(D __,--J 27- I
I w
-
c“;
aira
a
._ \ (l
N UNIVE
BR-l74
swim on Sat munY Lin»: 5v! do Dunn: we

46
ESTRAOA
c i 4' 2
3-7' If; BLOGOS E SUB-BLOCOS
N CURSO D‘AGJA
—— LIMITES DA AREA
--------- LIMITES cos sue - BLOOOS
acro ACAMPAMENTO
.—..-—~—- ESTRA m. 930 JETA DA

CHAPTER 5
MODELLING THE DIAMETER DISTRIBUTION OF AN UNDISTURBED FOREST
STAND IN THE BRAZILIAN AMAZON TROPICAL MOIST FOREST:
WEIBULL VERSUS EXPONENTIAL DISTRIBUTION
5.1. INTRODUCTION
Since total tree height is very difficult to measure
accurately, diameter is the most powerful simple tree
variable for estimating individual tree volume in the
Brazilian Amazon. Therefore, the quantification of diameter
distributions is fundamental to understanding the structure
of the growing stock and as a baseline for forest management
decisions. In‘addition, regardless of the species of tree,
Amazonian timber commercialization is commonly based only
upon the diameter distribution.
Bailey and Dell (1973) and Clutter et al. (1983) gave a
comprehensive review of diameter distribution models.
According to Clutter et a1. (1983), among various
statistical distributions, the Weibull distribution has been
used the most to model diameter distributions. These results
support Lawrence and Shier (1981), who stated that after the
exponential, the Weibull distribution is possibly the most
widely used distribution for population dynamics
applications.
47

48
The introduction of the Weibull distribution function
to problems related to forestry is attributed to Bailey and
Dell in 1973 (Zarnoch et al. 1982, Little 1983, Clutter et
a1. 1983, and Zarnoch and Dell 1985). Since then, this
distribution function has been used extensively for diameter
distribution of both even-aged and uneven-aged stands in the
USA.
The Weibull distribution has not yet been introduced in
tropical moist forests, especially in the Brazilian Amazon.
There, one of the most common models for diameter
distribution is still the exponential (Barros et a1. 1979
and Hosokawa 1981).
A comparison was made between the Weibull probability
density function and the exponential distribution as
diameter distribution models for Amazonian forests. The
hypothesized distribution functions were tested to see how
well they fit the observed diameters randomly taken from the
study area.
5.2. PROCEDURES
The Data
The data for this study were collected on the research
area of Forest Management Project conducted by the
Department of Tropical Silviculture of the National
Institute for Research in the Amazon (INPA) - described
earlier in Chapter 4.

49
The basic descriptive statistics of the study area are
presented in Table 5.1.
The diameter distribution functions
The Weibull probability density function and the
exponential were chosen for testing as a diameter
distribution model for the experimental area.
In this study, the estimators of Weibull parameters
were computed using the percentile (Zarnoch and Dell 1985)
and maximum likelihood (Cohen 1965) approaches. The
estimation of the exponential model parameters was done
according tx: Einsensmith (1985). These approaches are
described separately.
(1) Weibull Maximum Likelihood (MLE)
The Weibull distribution, which has the probability
density function:
f(x) = (c/b)x°"1 exp(-(x)°/b); x20, c>0, b>0
= 0, otherwise
has the following likelihood function for a sample of n
observations
L(xi, ..., xn; c, b) = n(c/b)xi°"l exp(-xi°/b) (1)
Taking the logarithm of (l)

50
1n L zln [(c/b)xi°"1 exp (-xi°/b)]
{[ln (C/b) + 1n Xic-l ‘ (Xic/b)]
ln 1n (c/b) + 2(c-1) 1n x- - (1/b)‘3xiC
By differentiation with respect to g and b in turn and
equating to zero, the following equations are obtained
d ln L/d c n/c + Zln xi - (l/b) zxic 1n xi
d In L/d b -(n/b) + (l/b2) r xic = 0
Taking b from (3)
b = ( zxi°)/n (4)
and substituting in (2) produces:
n/c + zln xi - [l/( ixic/n)] zxic 1n xi = 0
n [(l/c) - (xxic ln xi)/zxi°] = - zln x-
[inc 1n Xil/[inc] ' (l/C) = (l/n) zln Xi
(2)
(3)
(5)
The coefficient 2 can be estimated by any iterative
procedures or by a simple trial-and-error approach to
equalize both sides of equation (5). The coefficient _b can
be estimated by (4).

51
(2) Weibull Percentile (PERC)
The Weibull function using the percentile»estimators
has the probability density function
f(x) (c/b)[(x-a)/b]°‘1 exp {-[(x-a)/b]°}; x3a30,b>0,c>0
0, otherwise
The parameters a, b, and g are estimated as follows:
3» I- [xlxn - xzzl/[xl + xn - 2x2]
b = “a + X[.63n]
ln {[ln (1 - pk)l/[ln (1 - pi)]}
where: xi (1 = 1, 2, ... n) = the ith ascendent ordered
diameter; pi = 0.16731, and pk = 0.97366.
(3) Exponential
The parameter estimates of the first order exponential
function
Y = a*eb"'x
can be obtained by the linearization method (or Taylor
series). This is an iterative approach using the results of
linear least squares in a succession of stages. According to

52
Draper and Smith (1981), the steepest descent and
Marquardt's compromise can also be used. Here the
linearization method was used to estimate parameters a and
E.
The application pf the models £9 the data
In this context, 5 is the diameter in centimeters
measured at breast height (1.30 m) in 1980. The Weibull
parameters are defined as: a, the location parameter, which
can be the smallest dbh measured, b, the scale parameter,
which shows the relative range of values the dbhfs may
assume; and c, the shape parameter, which determines the
general form of the distribution (Zarnoch et a1. 1982).
For MLE estimators, (x - 24.9999)*was used to compute p
and 9 based on Cohen's two-parameter Weibull distribution,
after assuming a = 25 (the smallest dbh measured). The value
24.9999 was used instead of 25 only to avoid the logarithm
of zero, since no significant differences on the general
computation was detected.
The parameter estimates for all three models, Weibull
MLE, Weibull PERC and exponential, are presented in Table
5L2. Estimates are shown for the combined three sample plots
(bacia 3) and separately for each sample plot.
The Weibull cumulative distribution function was
determined by integrating the probability density function
for both the MLE and the PERC which provided the probability

53
for each dbh class. Then, the absolute frequency for each
dbh class was obtained by the product of the total number of
trees per hectare and its probability. The estimated
frequency using the exponential distribution was obtained by
the simple substitution of each dbh class as independent
variable in the equation.
To see if the three hypothesized distribution functions
fit the data in the sample, the chi-square test was used for
goodness of fit (Conover 1980). The null hypothesis was that
the distribution function of the observed random variable is
the Weibull MLE, or the Weibull PERC, or the exponential;
and the alternative hypothesis, otherwise.
5.3. DISCUSSION OF RESULTS
The diameter distribution for bacia 3 and for each
.sample plot (bloco l, bloco 2 and bloco 4) are presented
respectively in Tables 5.3, 5.4, 5.5 and 5.6.
Except for the Weibull MLE in bloco l and bloco 2, the
remaining computed chi-square's are not significant even for
°= .25, i.e., the null hypothesis cannot be rejected. The
best fit with the Weibull MLE model occurred in bloco 4
where the highest sample variance (32 = 224.86) was
observed. On the other hand, the best fit for the
exponential model was in bloco 2, which had with the lowest
sample variance (32 = 114.76), and the worst fit was in
bloco 4.
In this study, the Weibull PERC model was not seriously

54
affected by variation within the sample plot. It was very
consistent in fitting the observed data to all three sample
plots across a range of diameter Classes. The Weibull MLE
model, in contrast, consistently overestimated the frequency
of the first dbh class and underestimated the next four
classes.‘The Weibull MLE was consistent, after the first dbh
class, only in bloco 4. The exponential model, on the other
hand, was very consistent where the sample variance was low,
and inconsistent with higher variance mainly in estimating
the frequency of higher dbh classes. Bailey and Dell (1973)
pointed out that c < 1 should occur in all-aged stands of
tolerant species. Here the estimate of shape parameter was
greater than one, c = 1.02, only for bloco 2. For the other
sample plots and for the combined plots the 3's are smaller
than one.
When the three sample plots are analyzed together on a
per hectare basis, representing the entire experimental area
(bacia 3), all three hypothesized distribution functions fit
the observed data. This is demonstrated by the non-
significant chi-square values even for a==.25, although
the chi-square of the Weibull MLE model is about ten times
greater than the others.
Graphically the results of each hypothesized model for
the experimental area are presented in Figures 5.1, 5.2 and
5.3, respectively for the Weibull MLE, the Weibull PERC, and
the exponential. Figures 5.4, 5.5 and 5.6 represent the
relationships between the frequencies of observed dbh

55
classes and the frequencies estimated.by the hypothesized
models for each sample plots, respectively for bloco 1,
bloco 2 and bloco 4. As expected, except for the Weibull
PERC in bloco 2, the curves produced by the Weibull
distributions are reversed J because of the c parameter
(c<1). These graphics demonstrated that the Weibull PERC
produced the lowest dispersion of the observed data around
the hypothesized curves for the combined plots and also for
individual plots.
5.4. CONCLUSION
The results indicate that the Weibull PERC model is the
best model of the models tested to quantify the diameter
distribution of natural stands in the Brazilian Amazon.
The simplicity in estimating the Weibull PERC
parameters and its insensibility to the sample variation
demonstrated in this work are valuable attributes to be
considered. This distribution function can be used with a
reliable individual tree volume equation for modelling
forest growth and yield for the study area, or for similar
areas in other portions of the Amazon.
The exponential model also performed adequately in
fitting the observed data. However, it may be inconvenient
to estimate its parameters by the iterative approach. The
ease of this method will depend upon the available computing
capabilities.

Tabla
5.1:
Oiaeeter(ca)
descriptive
statistics
for
the
study
area.
UBLUES
BBOIB
3BLOOD
lBLOOD
2BLOOD
4
aofcases
1891
619
667
605
Hiniaua
25.00
25.00
25.00
25.00
Hauiaua
116.00
116.00
91.00
113.00
arithmeticmean
37.84
38.27
36.58
38.80
Saaplevariance
175.39
190.13
114.76
224.86
Ooef.
of
variation
35.00
36.03
29.29
38.65
56

Table
5.2:
Parameter
estimates
used
For
diameter
distribution
-hectare
basis.
com-F
a
coeF.
b13
14
12
13.5
coeF
c.94419
.9225)
1.01510
.QJIBO
a375.534
359.245
$6.185
$7.169
coeF.
b-.07409
-.0730
-.07250
-.07668
r"2
.99
.%
.99
.99
57

Table
5.3:
Diameterdistribution
For
all
three
sample
plots
together
(bacia
3)
derived
From
three
different
methods.
HUWUJEE
IEEELHI
EMHBWML
EF
a»:
08H(X)m
mm
mnmmmmmmmmmammwmmwmmm".
mm .1
.meeeeemeenzeemym mmmmu_aflaahlalZLL ........ mm
3.xammmepmmewemmmsemmie
i _.
emmm.emsaw.moem...mmxn&&25w753211 .mfi
m_»meemeemmmeemw.e.sfiwm1.52111 “
m.
”.meemmmmeemeemmemmme aE_93W185432111 fi
.nmemmeumsmusn momma m_.maemzeeut ..... ... a
lesseeseeeseeseeeeeeua
asyeaeeeanneeesmmmmmm
mdmfl¥=
5d”
man
IOBH(X)
=center
of
diameter
class
incentime
up»

mo
a mmmmfiymmmwfiammym..mmgmm
m umaammnma7m mmmm
.zaammzaaaun ........ “a
m mm1111mmw1mflmmmmflfifimmm$mm 1 .s
m nammmmu wm3$55a7§amsum
E afimelalzprhh ....... “a
tEIflLL
".5
.mfiwzxzm:smws1Zn5:$_$55m754322111
[Fun
5393177331111
4
154.75
152.3?
methods.
“nammmnwmammmmma omOfi
“5555555555555555555_
_a yaaasaannamuwmmm
TDTFI.
Tfile
5.4:
Diameter
distribution
for
Bloco
1derived
from
three
different
[84003:

Table
5.5:
Diameter
distribution
for
Bloco
2derived
from
threedifferent
”flak.
EflulFflE
IEWELH£
EWUGWML
3'
“It!”
UEH(X)u
mo
mnmfi.mm.fiflflflfiaflwmnmm 1 _4
_mfl..fiw..mmmgwa “a.&%572&54&&L...mfl
”mommmsmm...wfimm.flw. m 1 a
”2.2....mxflmflafi“.$356m74311 . m%
1.73423 m.
‘5
ml
25
(D
'5
00
a)
a!
25
4
25
O
25
25
manilarm
mfll
_wawaaawewnnamn_ I08H<X>
=center
of
diameter
class
incentimeter.
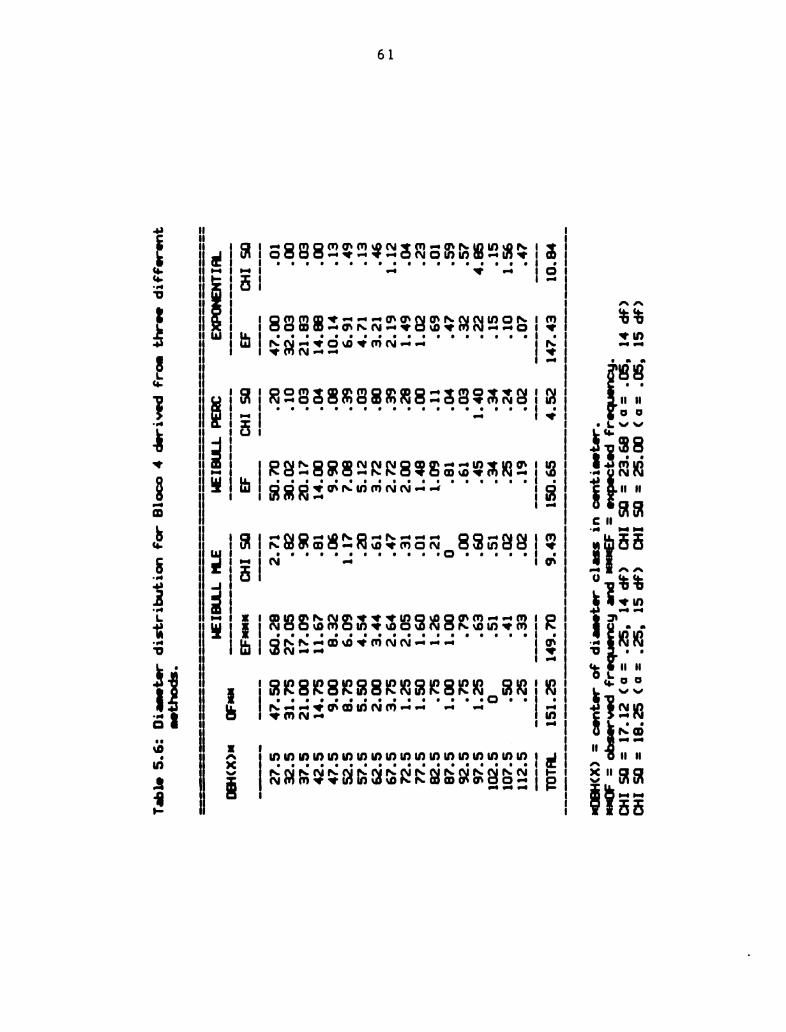
EXPCIENTIR.
Film
llE
metfods.
u-‘mm
UBH<X>I
Table
5.6:
Diameter
distribution
for
Bloco
4derived
from
three
different
my
Elm
PERC
EF
mmmmmmmemmumamaa mannaI. ...... cl" ..... «oh-mm
m“
“mammumnmww mnyawmwne
”maflmm&L&ZLL ....... mm
mmmwmmmfimwfiflmmmmwmmmnflm_ 1 “1
mmmmmmunnmmmmmmaawma.mmmuaz&1&&LL ...... “m
.mmmmmfiflmflflmmmomwflmmflem_2 1 .9
mmflmmflfim..flmm.mnam.a mE_wwwu864322L1L ..... &
.mnwnmnmmmamnwnaoma %
_manmamaaanh.n.n .. a
555£5£5£5fifififififijfifini
_aawenaweannmmuwmmm m

62
70-
{tree/ha
314
1L1
L1
13‘
N O
LL
Il
AL
L
..e
0 L4
.
A I
-..--___._V,,
10 20 30 40 50 50 70 80 90 100 110 120
DBH Gun)
PREDICTION BY WEIBULL MAX. UKEUHOOD
c211.
Figure 3.1: Becia 3 - The relationship between the observed
and estimated dbh frequencies, using the Heibull
HLE function.

Itroo/
r111
63
"5b"'eb"'7b” so so
DBH Own)
PREDICTION B'Y WEIBULL PERCENTILE
TM 110 120
Figure 5.2: Bacia 3 - The relationship between the observed
and estimated dbh frequencies, using the Heibull
PERC function.

ftree/ha
70-1
64
‘1'er VVY'VVV‘f' r'7111lvv'vrf
102030405000703'0901001101
DBH(cm)
PREDICTION BY EXPONENTIAL
Figure 5.3: Bacia 3 - The relationship between the observed
and estimated dbh frequencies, using the
Exponential function.

65
70
e m
a.o—eM
2 SD
\
g .. ."" so
3
ID
D
5......2,0 ..... 4'0.....i.....*mgh".¥b
DDH (cm)
(’0
§
.1“?
3 a can-usa can-us
fl-E Hm Ii!- “q HmI
2 “9 ad\ 3 g 3 .
g “5 2 “H L
‘Vwé ‘Kué
afiafi
0-3 01
40 I 1 20 40 u I 1
DB" (cm) . OBI-I (cm)
(a) (C)
Figure 5.4: Bloco l - The relationship between the observed
and the estieated dbh frequency distributions
by (A) Exponential, (B) Heibull PERC, and (C)
Heibull MLE functions.

66
:
- m
79.5O-e see-Its
”E
2 E\ a;
8 :
b{wj
- s
...;
aé
mi
113 -6IIIIIIgIIIII1%sssss3 iiiiiB'vagwsvvv‘k
DDH (cm)
(A)
‘H:
I “I“
mi
2 i
3 ”‘1of“?
3%
20-5
11.:
.3 '
40 I l
DBH(un)
Figure 5.5: Bloco 2 - The relationship between the observed
and the estimated dbh frequency distributions
by (A) Exponential, (B) Ueibull PERC, and (C)
Ueibull MLE functions.

67
70
e m
.0 v-e “I.“
2 SD
\
g 0
"3
ID
m ,
o .
N 1 1
DDI-I (cm)
(A)
70
a dame -.un—
fl' wwlfllun weaken
3‘” 3:»
\ \
9 ‘° 2in” In
w . 1 .
40 I l 40 u l 1
mfiIknd CBH(ufi)
(B) (C)
Figure 5.6: Bloco 4 - The relationship between the observed
and the estieated dbh frequency distributions
by (A) Exponential, (B) Heibull PERC, and (C)
Heibull "LE functions.

CHAPTER 6
A MARKOV CHAIN APPROACH TO PREDICT MORTALITY AND DIAMETER
DISTRIBUTION IN THE BRAZILIAN AMAZON.
6.1. INTRODUCTION
Little is known about forest structure and stand
dynamics of Amazonian tropical moist forests. Successive
records from representative long-term permanent plots
practically do not exist. The problem of reconstructing
forest history is greatly compounded by the fact that trees
can not be reliably aged, species diversity and spatial
heterogeneity are high, and fallen logs decay rapidly. It is
important to understand and report the natural changes that
occur in representative examples of pristine Amazonian
forests, because their composition and structure can be
altered by man as the demand for tropical timber species
increases.
The main objective of this chapter is to report S-year
changes in the overstory structure of an undisturbed
tropical moist forest. This will be done by the transition
probabilities of the overstory diameter distribution and
mortality of this forest, using a first-order Markov chain.
Diameter distribution and tree mortality will be projected
68

69
ahead to 1990 (t+2), based upon a 5-year period of
observations completed in 1985 (t+l) and its immediate past
in 1980 (t).
A first-order Markov chain is a stochastic process in
which the transition probabilities during the time interval
(t and t+l) depend only upon the state an individual is in
at time tior upon the knowledge of the immediate past at
til, not upon any previous state (Horn 1975, Chiang 1980,
and Bruner and Moser 1973). Shugart (1984) pointed out that
the time-invariant nature of each of the transition
probabilities is an important characteristic of the Markov
approach.
Shugart and West (1981) stressed that the importance of
understanding forest ecosystems is based not on their age,
but on known changes at present. Deterministic models
consisting of a single mathematical function (linear trend,
polynomial, sinusoids, or exponential growth or decay) have
not proven adequate when time series are involved (Morrison
1976).
In tropical moist forests, size may be more important
than age. One reason for this is that size may be more
ecologically informative than age when it is difficult to
make accurate estimates of age (Enright and Ogden 1979).
Division of life-cycles into developmental stages may allow
prediction of future behavior more accurately than division
into true age-classes. Usher (1966) used size attributes
instead of age to develop a model for the management of

70
renewable resources. He stressed that an organism which is
in i-th class at time t can be in the same class at time
£11, or it can be in a next class of that attribute, or it
can have died.
According to Enright and.Ogden (1979), the transition
matrix models in general are suitable for the analysis of
many biological problems, mainly in studies related to the
forest dynamics.
These models have been used intensively in studies of
dynamics of populations of plants or animals in many parts
of the world. Some examples are: the demography of jack-in-
the-pulpit in New York (Bierzychudek 1982); forest dynamics
of a population of Araucaria in a tropical rain forest in
Papua New Guinea, and Nothofagus in temperate montane forest
in New Zealand (Enright and Ogden 1979); termite succession
in Ghana (Usher 1979); forest succession in New Jersey (Horn
1975); the application, although without success. of this
model in secondary succession in coastal British Columbia
(Bellefleur 1981); the discussion of some extensions and
application of Hornfls Markov approach for forest dynamics in
tropical forests (Acevedo 1981); and the application of
Markov model to predict forest stand development (Usher
1966, Usher 1969, Bruner and Moser 1973, Peden et a1. 1973.
and Buogiorno and Michie 1980). Alder (1980) also described
the transition matrix as a possible tool for analysis of
growth and yield data for uneven aged mixed tropical
forests. Most of these works include a reasonable review

71
about the theory behind the Markov approach.<Grossman and
Turner (1974), Chiang (1980), and Anderson and Goodman
(1957) are very useful supplemental readings.
6.2. PROCEDURES
The data
The data for this study were collected on the research
area described in Chapter 4.
The Markov model
According to Bierzychudek (11982), a transition matrix
model is a size-classified model or a form of the Leslie
matrix model. The only requirement of this model is that the
population can be divisible into a set of states, and that
there exist probabilities for movement from one state to
another over time (Enright and Ogden 1979).
Here let the states be i, j = l, 2, .u., m. Let the
times of observation be t = 0, l, ...., T, and let Eli (t+1)
(i, j = 1, 2, ...., m) be the probability of state 1 at time
£11, given state i at time t.
A Markov process {X(t), t E [0,a>]} is said to be
homogenous with respect to time, or time homogenous. if the
transition probability
Pij(tvt+1) = Pr lX(t+1)=j|X(t)=il. i.j = 1, 2, ...., m.

72
depends only on the difference between t and t+1, but not on
t or t+l separately (Chiang 1980).
The computation of this probability can be done as
follows.
where: nij
First, calculate
Pij
given class i at time 3,
individuals in class i at time t.
= "ii/"i
and r1-
= number of individuals in class j.at time t+1,
= total number of
The transition probability matrix of a Markov chain for
a n-state process can be set up as:
j=l j=2 j=3 ..... J=m
r- “_I
1‘1 P11 P12 913 -°-°- Pim
i=2 P21 922 923 -°°-- sz
P = (pij) i=3 P31 P32 P33 °°°°° 93m
1:” Lfml sz pm3 "°°° pmm [ The probabilities Pij are nonnegative and the sum p11 + p12
+ pi3 + see Pim = 1a
The transition probability Pij can be of n-step
transition probability, pijI“). as the probability that the
population goes from state i on one trial to state i 3
trials later.1According to Bruner and Moser (1973), the n-
step transition probabilities matrix may be obtained by the

73
equation
PI“) = P“
where PI“) is the matrix of n-step transition probabilities
and Pn is the initial transition matrix raised to the n-th
power.
In this work, 15 states (i, j = l, 2, 3, u. 15) were
established as follow: state 1 = ingrowth (1), states 2 to
14 were defined as dbh classes, from 25 to the generalized
class next > 80 cm, in 5-cm interval, and state 15 =
mortality (M). Ingrowth is defined as those trees not tagged
in 1980 which in 1985 reached dbh >, 25 cm. The time interval
t and 511 are respectively, 1980 and 1985.
Tables 6.1, 6.2 and 6.3 present the transition of the
absolute frequency of individuals from the state i to state
1 during a 5-year period, respectively for bloco 1, bloco 2,
and bloco 4. The state ingrowth does not appear at time 1980
because it means only the movement to the higher dbh class
from the generalized dbh < 25 cm class.
The probability for transition among states was based
on the frequency of trees which either remained in the same
class, moved to a higher class, or died during a 5-year
period. Tables 6.4, 6.5 and 6.6 present the transitional
matrices for blocos 1, 2 and 4, respectively. These tables
were set up using their counterparts, Tables 6.1, 6.2 and

74
6.3, as bases for the computation of probabilities. For
example, the probabilities for the state 25 cm dbh class for
bloco 1 (Table 6.4) were calculated as follows: pq'z =
155/183 = 0.8470, 122,3 = 16/183 = 0.0874, and 112,15 = 12/183
= 0.0656. From all trees in 25 cm dbh class measured in
1980, 84.7% remained in the same class, 8.74% moved to the
30 cm dbh class, and 6.56% died during the period 1980-
1985. The probabilities for other dbh classes and blocks
were similarly determined with the respective counterpart
tables with absolute frequency distribution.
The tw0wstep transition matrix for each block (Tables
6.7, 6.8 and 6.9) were obtained by squaring their
counterparts (Tables 6.4, 6.5 and 6.6), respectively for
blocos l, 2 and 4. These tables represent the probability
for dbh and mortality distribution after two 5-year periods,
i.e., for t_+3, year 1990. The two-step transition matrix is
the basis for predicting the distribution of diameter and
mortality for the study area in 1990.
The eigenvalues (111-) of the transition matrix of each
sample plot were determined according to Anton (1973). The
dominant eigenvalue (Al = 1 since each matrix is non-
negative and row sums are 1) and the next largest modulus (
12) were determined. These values provide the ratio (Al/12)
which, according to Usher (1979), indicates the speed with
which the system will approach the ”climax" state.

75
6.3. DISCUSSION OF RESULTS
The projections for 1990 of number of survivors from
1980, the frequency distribution of dbh classes, and the
mortality by dbh classes, respectively for blocos 1, 2 and 4
are presented in Tables 6.10, 6.11 and 6.12. These
projections were determined based on the product of the two-
step transition matrix and the initial values of each state.
Volume stocking in 1990 can be estimated by applying a
reliable individual tree volume equation to the projected
diameter distributions. The frequency of individuals per dbh
classes is available for each sample plot.
The plot ratios (Al/A2) were 1.15, 1.00 and 1.00,
respectively for blocos 1, 2 and 4. The mean ratio, 1.05,
suggests that the studied area will approach the "climax"
state slower than the two systems discussed by Usher (1979),
mixed hardwoods in Connecticut (ratio 1.34) and in New
Jersey (ratio 1.57). This result makes sense if compared
with the distribution of changes in dbh classes and
mortality which occurred over a 5-year period. Using Table
6.13, which is the summary'of one-step transition matrix,
the mean estimates of the probability of changes and
mortality per plot are respectively 0.1205 and 0.0918. This
means that 12.05% of the total number of trees in a plot
changed dbh classes, and that 9.18% died during a 5-year
period. In an absolute basis, using the mean number of
trees/plot = 631, 76 trees changed classes, 58 died, and 23
(the mean ingrowth/plot) grew into the measurable dbh

76
classes. Thus, these results suggest that the studied area
is not a static population.
The projection for 1990, based on the summary presented
in Table 6.14, also does not show any trends that this
population is not changing. The average of the probability
of changes of dbh classes increased to 0.1895 and mortality
to 0.1717.
6.4. CONCLUSION
In the study area, the average rates of mortality and
ingrowth, during a 5-year period of observation, are
respectively 9.18% and 3.72% in relation to the total of
initial number of trees recorded in 1980.
There is no evidence that the probability of mortality
increases as dbh increase. The same trend is observed for
changes in dbh classes (the movement from one class to
another), i.e, the changes are occurring independently of
the diameter size.1As this study dealt with only the control
plots, it will be very interesting to compare these plots
with other experimental plots to see how effective were the
silvicultural treatments to change the rates of mortality
and ingrowth.
The ratio (Al/A2) leads to the conclusion that the
population under investigation is not static, that changes
are still taking place, and that the rates of ingrowth and
death are not perfectly balanced. However, it is also

77
necessary to keep in mind that this population is truncated
by size, i.e., only trees with dbh ; 25 cm were involved.
The Markov approach has a lot of potential. It can be
used as a baseline to project the mortality and diameter
distribution, or at least to predict the direction of future
trends, for forest management purposes in natural stands of
the Brazilian Amazon. It provides a general insight into the
nature of the dyamics of a sample of pristine Amazonian
forest which, consequently, will be very helpful to assist
decision makers in exploring and understanding the Amazonian
forest issues.
In 1990 this procedure will be repeated. Then, the
Markov chain approach will be evaluated and, if necessary,
refined based upon a 10-year of observatitun If valid, the
projection ahead to year 2000 will be possible.

Table
6.1:
Bloco
1-Transition
fromone
state
(i)
to
another
(j)
duringa
5-gear
period.
25
30
35
4D
45
50
55
6D
65
7D
75
>90newt
Htotal
0-0
“'3
26
26
155
16
12
193
109
27
17
153
52
22
1
45
4
33
wt
0'1
mvmm NNN d
"(Inn
11
2
“I“8889988882E§§:!
total
191
125
79
67
3B
31
29
13
ll
33
12
252
646
total/1990
=646
-l
(ingrowth)
=620
andtotal/1995
=646
-H
(mortalitg)
=594.
(I)
blank
spacesmean
zeros.
78

Table
6.2:
Bloco2
-Transition
fromonestate
(i)
to
another
(j)
tiringa
53-year
period.
jI
25
33
35
4D
45
50
55
6D
65
7D
75
>83next
Htotal
'd
20
172
20
122
23
112
87
13
7
46
16
8§§§88828**"*
“888998888288§:l
192
142
110
6D
62
25
19
17
65
24
45
i1
33
total/1m
=6m
-I
(ing’outh)
=668
andtotal/1%
=688
-H
(eortalitg)
=643.
(I)
blarkspacesmean
zeros.
79

Tdale
6.3:
Bloco4
-Transition
fromonestate
(i)
toanother
(j)
tiring
a5-gear
period.
Wmmmm
j125m§4045505560657075>mnextfltota1
an
24
24
15)
127
3$IIIIIIRI
NCJNQNWN-‘d
Q
91’
QIDIDWQn F.
N
II.
10an
ID
"namesamssetggglg
total/1m
=629
-I
(ingrowth)
=605
andtotal/1%!)
=629
-H
(mortalitg)
=567.
(it)bldk
spacesmean
zeros.
80

Tdale
6.4:
Dlooo
1-Transitionprobabilitgmatrix
fromone
state
(i)
toanther
(j)(bring
a5?.-
period.
ma—W
W***—M
j125
a)
54D
45
50
55
6D
65
7D
75
mnewt
I!
0‘
1.1111)
.9470
.CB74
(556
.7124
.1765
1111
.6667
.2921
.0129
035
.9333
.0741
.015
.0741
.7513
.11%
.154
0%9
.1667
.7692
.0769
15$
5!!!)
.7500
.25CIJ
.757
.1429
.0714
1
E
"8889988882883.
on
s-Ov-O
(in)blank
spacesmen
zeros.
81

82
.OOLONcoo.
IIOIQI
Jenna
any
aaao._
:
auuo..
axuc
oaaa._
aoA
aaao._
mu
comm.
comm.
an
aamw.
anon.
aamw.
mm
aamw.
cams
om
mo.~.
.mmo_.
wen»
mm
v_~o
man_
cams.
_am
mama.
mmno.
msua.
me
name.
mmmw.
mmnm.
av
vmma.
m.~..
_mdm.
mm
mane.
amen.
wmv_.
mass.
on.
nave
ammo.
m.mu.
mu
aaau..
um
zaxis
amA
ms
as
no
am
am
an
ac
av
an
an
m~
_n
.uomL-uL-cmumamcmgnu
Aw.
Luzaocno»
“my
.gnuu.cosaga
xmggncmum—«nunogu
camamncugp
uucoo—a
"m.uo—nup

83
60.30SeeI:
v1.325
oaaa._
:
noun."
axg:
__~_.
wmmo.
mmmm
aoA
anon.“
mu
acne.“
on
sumo.
same.
«man.
mu
am~..
omsm.
ow
mama.
mama.
~u_a.
mm
m~¢u.
smug.
v.n~.
om
wmma.
44mg.
cums
mv
mam".
mmm~
can“.
av
88.
:3.
~96.
mm
mvoo.
_mma.
tuna.
am
mm_~.
ages.
«mun.
am_a.
nu
ouao._
um
:ax.c
auA
mm
as
mm
um
mm
on
mv
av
mm
on
mw
na
.uowgonLucmrmamcmLau
any
Lasagna
0»
“my
canon
can«cum
xwgancmum—Mn-nogn:o.u.a¢og»
uvcacao
um.mo~nup

84
'30.»:Luau
eased;
)petq
(u)
0000'!
NW
9022'
8211'
8219'
2580'
665'
9295'
“2'
”16'
9260'
4169'
59m
9900'
5921'
5299'
W'
1110'
5512'
2191'
69(1'
6210'
[291'
9059
2692'
0110'
0921
2491'
81m'm'
6211'
9969'
M'
4900'
(580'
2829'
3999'
01.61
W'
590'
9892'
9218'
6121'
9310'
6%1'
9214'
9590'
9M'
0490'
~nameeamsaen§§=
0!.
aan."
oa<
94
oz
99
09
93
as
s»
or
as
as
92
1r
‘d4....-_.
...—.7.-WW+H4—H_.
...—4
'xsawtvfizstsq-qoaduoyzysUOJQ
6018-001CH1
-1
00013
=z'9
OIQIL

6.0:
Bloco
2-
111atin-steptransitionprobdailitg
eatrix.
1¢1e
an
540
45
50
55
60
j1a
.1
115111
oooooooooo
.238
.0260
.0015
.6611
.1010
.02“
.4577
.334
.
.7533
.7251
.1607
.0144
.mas
.53
.515
.09!)
ramsmamswamnmmn 00
blank
spaceseean
zeros.

Table
6.9:
Bloco
4-The
tun-step
transitionprobability
eatrix.
-....--mmm-m“...-“...“...mm
jI
25
30
35
40
45
50
55
60
65
70
75
>00
next
H
1.0150
.0632
.0053
.6655
.1150
.0123
.0004
.7144
.1124
.0097
.5311
.2005
.0264
.5625
.2950
.0167
.5951
.1362
.0070
.6695
.1539
.7656
.7512
.1245
1.0000
1.0000
.6944
.0463
.25
”WW"9:
mameeamaaargfx
l l.
(l)
blank
spaces
eean
zeros.
86

Table
6.10:
Bloco
1-Projection
For
1990.
eortalitg
dias.
distr.
survivors
0trees
m
in
1900
m
mu
13 6705 343 3
95m” .1
_nfiunwseaammmn
.
3463022201.
12
17
* mxnnmsumsaamn
m
.xamsmemsmemnmmu
102.10
543.02
_m
_m
...
z.

Tdale
6.11:
01oco2
-Projection
For
1990.
Otrees
din.
distr.
nrflwn:
nddfig 1
1g)
11'!
mm
”mauxmsnazmmm
”flaw-5461.1
%_ 02763240100011
l
_ .2..........m 0 7 64563L
1_ mmufi6all
% 20 076524
.. muuwgi
mgawmmaafifimmm
m wmxaaaaumzmnml 11
mammmmumamm341.
mmmmgawm441.
m _xamamam5wemnmmn_w

m.uwmgugmsmgmsmg1_mas
GENE
a.
MTU
m9§m0a§fl$fimw
t
.11
mgsn.mmnmummn_a
assaamewnummm
ammaaummuamaa
Issaastmgsuww
a99n63$9umm
mmmwaummwgwam
IaZOOIIZSZB‘Z
9mmwmwemeummu
g
I
,._m
m1
“3
“an.
”mums
wasp
"'19
Envy“
.mst
403mEWFO-‘d
-V00010'2‘?"Q"

90
in
1905.
bloco4
bloco2
bloco
1
1
Table
6.13:
Suseargof
one-step
transitionprobabilityeatrix
-
H
0656
1111
0305
0741
1364
0960
0
1667
1530
5000
0
0714
3
classes
'IGSIBS'QRIBSSRIQQ l '3“:

1&1e
6.14:
Sine-'9of
too-step
transition
probabilityaatrix
-projection
'88399883529 I"
l Ml?
.1339
.1970
.1477
.2497
.1749
.m
.3105
.0179
2.6626
.alg
91

CHAPTER 7
SHORT-TERM GROWTH OF UNDISTURBED BRAZILIAN AMAZON TROPICAL
MOIST FOREST OF "TERRA FIRME"
7 . 1 . INTRODUCTION
Based upon the available literature about growth and
yield studies, the mixed uneven-aged stands of the Amazonian
forest are condemned to stay where they have always been.
These forests are not an attractive forest investment
because little is known of the past growth and future growth
potential. Age and site index, two fundamental variables
used in developing even the simplest growth and yield models
(Sullivan and Clutter 1972, Ferguson and Leech 1978, Alder
1980, Smith 1983, and Clutter et al. 1983) are not
available. In addition, long-term successive measurements on
permanent sample plots are non-existent.
The main objective of this work is to give'a starting
point for growth and yield studies in the Brazilian Amazon
based on a 5-year period of observation. A major constraint
is that age, site index and total height of trees are not
available or even practical to obtain. Therefore, only
diameters measured in 1980 and their 5-year increments will
be used in an attempt to project growth and yield. Another
objective is to avoid the problems that the tropical
92

93
countries of southeast Asia have faced in terms of
divulgation of their experiences with growth and yield
studies. In Peninsular Malaysia, for example, although
growth and yield studies were established back in the
1950's, almost thirty years later few analyses have been
reported (Tang and Mohd 1981). Revilla (1981) also pointed
out that the growth and yield studies reported in Malaysia
and Philippines do not reflect the abundance of growth data
available.
7.2. PROCEDURES
This work focused on two separate aspects of growth and
yield, the current volume estimation for individual trees
and prediction of future volume.
In individual current volume estimation, the emphasis
was on the selection of the best model to estimate the 1980
and 1985 merchantable volumes based on either single-entry
(dbh as independent variable) or double-entry (dbh and
height as independent variables) regression equations. The
variable height (H) from double entry models was estimated
from diameter-height equations. Several regression equation
models were tested. The selection of the best model for
volume estimation was based on the Furnival index (Furnival
1961) - adjusted standard error of estimate (SEE) used to
compare logarithm equations with non-logarithm equations -
residual analysis and the coefficient of determination (R2).
In yield information and prediction, the emphasis was

94
on the development of individual volume growth models based
on diameter or basal area increments, the development of a
model for volume of 1985 as a function of the diameter or
volume measured in 1980, and the use of the exponential
Lotka's growth model for volume prediction on a hectare
basis.
The data
The data used to develop models for current individual
volume estimations came from Higuchi and Ramm (1985). For
this study, trees with dbh < 20 cm were excluded leaving a
total of 654 cases. Table 7.1 presents the basic
distributional characteristics of the data.
For yield information and prediction, the data came
from the three four-hectare permanent plots described in
detail in Chapter 4. I
The quantitative information for all three sample plots
are summarized in Table 7.3. These data refer to 52
botanical families found in the study area, including about
350 different tree species with dbh ; 25 cm. Three families
(Lecythidaceae, Leguminosae and Sapotaceae) contributed 50%
of the total number of trees. Table 7.4 presents the
distribution of frequencies of the three dominant families
in terms of status in 1980, mortality (M), ingrowth (I), and
various classes of periodic increment (PI).

95
Model development
From the available literature, models were selected
which matched the variables available for the study area.
For individual tree volume estimation, the following
models were tested:
(a) Single-entry models (Loetsch et al. 1973)
v = a + b*02 (1)
v = a + b*D + c*02 (2)
log V = a + b*log D (3)
log V = a + b*log D + c*(l/D) (4)
(b) Diameter/Height models
b*D + cm2 + d*D3 (Clutter 1963)+H=a
log H a + b*(l/D) (Loetsch et al. 1973)
l/H II
a:
+ b*(l/D) (Rai 1979) (7)
log H a + b*log D (Schreuder et al. 1979)
(c) Double-entry models (Loetsch et al. 1973)
log V = a + b*log D + c*log H or V = a*Db*H°
v = a + b*DZ*H (10)
For all models:
log denotes logarithm to base 10.
(5)
(5)
(8)
(9)

96
D denotes diameter at breast height (dbh) outside bark
in centimeters (cm). It is measured at 1.3 m above
ground level.
H denotes merchantable height in meters (m), i.e., the
length of stem from the ground to the crown.
V denotes merchantable volume in cubic meters (cu.m.).
For individual yield prediction, the following models
were tested:
(a) Increment
d0 = a + b*D + c*D2 (West 1980) (11)
dBA = a + b*D + c*02 (West 1980) (12)
d0 = a + b*(D - 25)2 (West 1981) (13)
Where: do = periodic diameter increment in cm.
dBA = periodic basal area increment in squared
meters (sq.m.).
These three models were weighted using the inverse of
the estimated sample variance as weight for each diameter
class. The weighted models will be equations (14), (15) and
(16).
(b) Volume in 1985 = f (1980 vol. or 1980 dbh)
V(85) a + b*D(80) + c*(D(80))2 (17)
0(85) a + b*D(80) (Soekotjo 1981) (18)

97
Where: V(85) = volume estimated in 1985 in cu.m.
0(85) dbh measured in 1985 in cm.
dbh measured in 1980 in cm.0(80)
(c) Lotka equation (Pielou 1977)
Adapting this model to predict volume growth produced
V(t) = V(0)*ert (19)
Where: V(t) = volume at time t (for t = l, 2, n 5-year
periods) in cu.m./ha.
V(0) = volume at time 0 (1980) in cu.m./ha.
r = b - d = the intrinsic rate of natural increase.
b= [I + Increment]/V(t) = ingrowth (I) and increment
rate (flow-in quantity).
d=M/V(t)= mortality (M) rate(flow-out quantity).
7.3. DISCUSSION OF RESULTS
Only 1/3 of the total of species belonging to the three
dominant families are considered commercial species by local
markets in Manaus. There exists no occurrence of the two
most valuable species for exportation of the Amazonian
forests, Swietenia macrophylla King (Mahogany) and Cedrela
odorata L"
The individual dbh periodic increment (PI) of the study
area averaged 1.06 cm, equivalent to 0.21 cm/year. This mean
P1 was estimated based on a population from which 31.3% did

98
not have any increment at all (Table 7.4). The mean dbh for
each increment class is also presented in Table 7.4. Note
that the maximum mean increment occurred in trees with 40.9
cm dbh, and that zero-increment occurred in trees with 42.5
cm dbh. More than 80% of trees had a PI less than 2 cm.
The average periodic annual increment (PAI), 0.21
cm/year, can be compared with the long-term PAI of 0.10 to
0.12 cm/yr obtained in Puerto Rico, Maricao and Luquillo
forests (Weaver 1982), and with the PAI of 0.22 to 0.48
cm/yr from the southern Ontario hardwood forest (West 1979).
The PAI for ”pau-rosa” (Aniba duckei Kostermans) at Ducke
Reserve, about 20 km north of Manaus, was 0.38 cm/year
(Alencar and Araujo 1981). In southeast Asia, however, the
dbh PAI's for virgin or managed forests are at least twice
as large as the PAI of the study area (Miller 1981 and Tang
and Mohd 1981).
Although the PAI is positive, the stand stocking
decreased during the period 1980-1985 in terms of number of
trees, basal area and volume (Table 7.3). This is explained
by a mortality rate which was twice the ingrowth rate.
Another explanation for the decreases is the mean dbh's for
mortality and ingrowth, which were 39 cm and 26.3 cm,
respectively (Table 7.3).
The regression models for individual trees volume
estimation were developed using the ordinary least squares
method. The regression summary for these models is presented
in Table 7.2.

99
The diameter/height models did not perform adequately
and, therefore, they were not used. Probably the reason for
failure in fitting the diameter/height curve is because
merchantable height was used instead of total height. All
proposed models used total height. In Rad's (1979) work, for
example, a R2 = 0.956 was obtained, while the highest R2 of
this work was 0.154.
To estimate the current volume of 1980 and 1985, the
following equation was used
log V = -3.4033 + 2.2673*log D (3)
This equation was chosen because it presented an appropriate
residual distribution, had an acceptable R2 and SEE, and
because it was as precise as the equation (4) with three
coefficients.
Before developing the proposed regression equations for
increment and growth' studies, a contingency table was
developed to test the differences in probabilities among
sample plots (Conover 1980).‘This test was carried out to
test the possibility of pooling the sample plots. In this
case let the probability of a randomly selected value from
the i-th bloco being classified in the j-th category be
denoted by Pij' for i = l, 2, 3, and j = l to 5.
The hypotheses were:
HO: All of the probabilities in the same sample plot are
equal to each other (i.e., plj = p2j = p3j for all j).

100
El: At least two of the probabilities in the same sample
plot are not equal to each other.
The chi-square test for differences in probabilities
was carried.out based on the tabulated data from Table 7.3
for number of trees and the mean diameter for each sample
plot. For contingency tables, the rows (j) were constituted
of different categories (status in 1980, status in 1985,
ingrowth, mortality, and periodic increment), and the
columns (i) by the sample plots (bloco l, bloco 2 and bloco
4). For both number of trees and mean diameter, the null
hypotheses could not be rejected. All sample plots were
combined therefore for further development of the growth and
yield models. .
All increment models developed had very poor fits, as
demonstrated by the low R2 values and high SBEs in Table
7.5. Model fit was not improved through the use of weighted
least squares. Based on these results S-year periodic
increments should not be used as a baseline for projection
of growth and yield of the Amazonian forests. A possible
explanation of this result is shown in Table 7.6a, which
contains the mean, standard deviation, minimum and maximum
increment.by dbh classes.‘The same procedure was used for
dbh when increment classes were considered (Table 7.6b). The
contingency table was used to test the differences in
probabilities for mean, standard deviation and maximum
increment and dbh. There were no differences among these
values for both diameter classes and increment classes.

101
Statistically, this means that the mean increment is not
significantly different between.dbh classes, and that the
mean dbh is equal for all increment classes.
In contrast, the two models, equations (17) and (18),
developed for individual volume and diameter growth
performed very well. The explanation for this successful fit
can be found in the explanation for the failure of increment
models. In model (16), for example, the objective was to
study the relationship between the dbh measured in 1985 and
the same dbh measured in 1980. As the increments were very
small, non-negative and non-significant, the dependent and
independent variables were approximately equal. In both
equations the regression coefficients were highly
significant. With these models one may now predict the
individual volume or dbh growth for another period of time,
for 1990, based on the dbh measured in 1985. In 1990, these
models can be validated and refined using, then, a lO-year
period.
Finally, fitting Lotka's model (19) to the data
produced
V(t) = v(o)*e(-0.0347*t)
The intrinsic rate of increase, r, was obtained on a
hectare basis based on the data from Table 7.3 as follows:
0' ll (3.8197 + 12.8010)/ 273.6415 = 0.0617
D: II 26.1019/273.6415 = 0.0954

102
r = 0.0617 - 0.0954 = - 0.0347
Using this equation, the estimated volume in 1985 is
V(1) = v(o)*e('0.0347*1)
V(1) = 274.7256 cu.m./ha
and for 1990
V(2) = V(0):e('0.0347*2)
V(2) = 265.3458 cu.m./ha
Based on the intrinsic rate of natural changes in a 5-
year period, the yield estimation for 1985 is very close to
the observed yield (V(85) = 273.642 cu.m). The projection
for 1990 also looks acceptable. This means that Lotkafis
model seems very promising to predict the future volume
yield. However, a longer period of observation is necessary
to fully validate this model.
7.4. CONCLUSION
In the study area the dbh increment for individual
trees during the period 1980-1985 averaged 1.06 cm. The
mortality rate was twice the ingrowth rate, and the stand
stocking decreased around 4% during this period.
Two models, V(85) = a + b*D(80) + c*(o(80))2 and
Lotka's model, could be used to estimate individual tree
volume for the next period with an acceptable reliability.

103
In 1990, then, these models can be retested, validated, and
refined, if necessary, using a lO-year period of
observation.
There was no indication that dbh could be used to
predict either the merchantable height or short-term
diameter/basal area increment.
Traditional growth and yield models can not be applied
to the Brazilian Amazon forests since age and site index are
not available. The only alternative remaining seems to be
the use of simpler models based on stand structure
monitoring at successive occasions.
The findings of this study suggest that the growth and
yield studies are not possible only on temperate forests,
but they are also feasible on an undisturbed tropical moist
forest in the Amazon. To achieve this goal, however, the
best estimation of the current volume and the dependability
of the permanent sample plots must be pursued.

T‘le
7.1:
Basic
distributional
characteristics
of
thedatamed
For
indivirhalvolmewion
equations
(pooled
data).
“I”
duh
(ca)
1.1951.
(a)
volume
(cu.m.
)
N
a 654
654
w
FEM
STEM.
HIN.
41.602
17.166
20.000
15.502
3.654
5.200
2.373
2.753
0.172
V
max.
120.!!!)
27.CID
27.”
104

Table
7.2:
Regression
summary
For
volume
estimation
mode1s1.
WWWW
600011005
ab
cd
0‘2
SEE
F1
(a)
Single-entry
U=
a+b0‘2
-0.3504
0.0013
0.90
0.0711
0.0711
9=
a+5
0+
c012
0.7353
-0.0451
0.0017
0.90
0.0504
0.0504
159u
=a
+b
1590
-3.4033
2.2573
0.92
0.1094
0.1729
109v
=a
+b
1090
+c
(170)
-2.5209
1.0390
-7.3077
0.92
0.1004
0.1714
(b)
Diameter/HeiQIt
logH
=a
+5
(1/0)
1.3009
-4.4002
0.15
0.0994
1.5152
1/H
=a
+b
(170)
0.0405
0.7133
0.13
0.0175
3.0007
logH
=a
+5
1590
0.7039
0.2492
0.14
0.1002
1.5204
H=
a+b
0+
cm2
400.3
4.7500
0.5015
-0.0057
0.0001
0.15
3.3512
3.3512 1002=
coefficient
of
determination,
SEE
=standard
error
of
estimate,
F1
=Furnival
index,
and
a,
b.
c,
andd
=regression
coefficients.
105

Table 7.3:
106
Characteristics of the data used as yield
information and for yield prediction.
STATUS # CASES MEAN TOTAL TOTAL
N dbh(cm) ba(sq.m.) vol(cu.m.)
BLOOD 1
1900 620 30.4 02.100 1165.974
1905 594 30.7 70.690 1111.470
Ingrowth 26 26.2 1.403 16.910
Mortality 52 41.3 0.507 127.000
P1 560 1.1 3.695 56.475
BLOCO 2
1900 667 36.7 77.060 1073.065
1905 642 37.3 77.300 1070.691
Ingrowth 20 26.6 1.109 13.421
Mortality 45 37.9 5.463 74.971
Pl 622 1.0 3.001 50.375
BLOCO 4
1900 605 30.0 02.210 1173.139
1905 567 39.1 70.163 1101.529
Ingrowth 24 26.1 1.200 15.506
Mortality 62 37.0 7.053 110.372
P1 543 1.0 2.517 30.762
MEANS
1900 631 30.0 00.726 1137.659
1905 601 30.4 70.056 1094.566
Ingrowth 23 26.3 1.267 15.279
Mortality 53 39.0 7.274 104.400
P1 570 1.0 3.330 51.204
MEANS/hectare
1900 150 20.102 204.415
1905 150 19.514 273.642
Ingrouth 6 0.317 3.020
Mortality 13 1.019 26.102
P1 145 0.034 12.001
P1 - Periodic increment a only trees measured in both
occasions, i.e., number of remaining trees in 1905 excluding
ingrowth which was not counted in 1900.

Tdale
7.4:
The
Frequencydistribution
of
the
three
dominantFailies
by
status
in
1%],
mortality
(M)
and
ing-outh
(1),
andbyperiodic
increment
(P1)
classes
in
cm.
WW.
P1=0
W1<1
1<P1<2
2<PI<3
3<Pl<4
Pl>4
116
92
FFHILIES
19m
fl
Lecythidaceae
347
19
75
Leg-inosae
333
Smtaceae
277
TOTE-3
Families
£7
76
61
203
212
00
91
59
70
41
13
19
77
_e—gne
.laezlsg:
227
29
TOTE-shale
area
1092
542
433
440
105
60
57
was)
42.5
33.6
35
5
107

108
[ID'O
9+32'
“(1'0
9+32'
100'0
0&0'11
CID'O
026'90
2(lJ'0
“9'9
9601
'1
9101'0
1100'1
0000'!
HID?
9201'1
(”3'0
9%1'1
066'0
366'0
91100
00000-
20000-
(“13'0-
ZEIII'D-
2%6'0
$20?-
IM'D-
KID'O
9910'0
1CD'0-
7000'0
9610'0
MW
0950
9046'0
19W'0-
0599'0
665'0
6903'0-
4275'0
(001%!
=(g)0
zvamamammw
=(QM)
‘1”
WHO-‘5
(9)
Zv(QZ-0)
9+
v=UP
240
9+
09
+t
=000
240
9+09
+t
=on
909961-H
<9)
zv(sz-a>
9+t
=on
290
9+09
+t
=HEP
2.0
9+09
+t
=0P
009951-n-uou
(r)
901MB
-s[apoeW
(:1)
p1.-
‘stapoe
sue-0401.11
pczubscn
(9)
‘stcpo-
1050.450;
powufisan-uou
(t)
493
9499-409188-4500
=9'201901

109
Table 7.6: Mean, standard deviation, minimum and maximum for
each (a) dbh classes and (b) increment classes.
(0) Periodic increment (PI) by dbh classes.
INCREMENT (cm)
DBH CLASSES(cm)
N MEAN STD.DEV. MIN MAX
25-29.9 531‘ 0.9177_ 0.9854 0.0 8.0
30-34.9 397 1.0091 1.1246 0.0 6.0
35-39.9 252 0.9274 1.1108 0.0 6.0
40-44.9 163 1.1313 1.2349 0.0 8.0
45-49.9 122 0.8852 0.9474 0.0 4.2
50-54.9 84 1.0964 1.3398 0.0 8.0
55-59.9 65 0.9862 1.4453 0.0 8.0
>60 119 0.7483 1.5173 0.0 7.2
overall 1733 0.9573 1.1400 0.0 8.0
(b) 00H by periodic increment (PI) classes.
00H (cm)
PI CLASSEStcm)
N MEAN STD.DEV. MIN MAX
0.0-0.49 672 40.6706 16.3201 25.0 116.0
0.5-0.99 303 33.7709 9.3157 25.0 92.0
1.0-1.49 312 35.5929 10.3603 25.0 94.0
1.5-1.99 136 35.6610 0.9577 25.0 62.0
2.0-2.49 130 30.4615 11.4764 25.0 76.0
2.5-2.99 55 39.0727 11.5002 25.0 73.0
>3.00 125 30.7200 12.7459 25.0 91.0
overall 1733 37.0697 13.5655 25.0 116.0

CHAPTER 8
CONCLUSIONS
The forest resources of the study area are declining
with respect to number of trees per unit area, growing stock
volume and basal area.
The dominant latent root, A.= 0.97, supports the
previous affirmation. According to Enright and Ogden (1979).
when the intrinsic rate of natural increase (r) is equal to
zero, A is equal 1, i.e., the population under investigation
is perfectly balanced or the birth rate is equal to the
mortality rate; when r > 0, A > 1, the population is
increasing and the birth rate is greater than the mortality
rate; and when r < 0, A < l, the population is declining
or the birth rate is smaller than the mortality rate.
Another reason for the decline is that mortality and
ingrowth are not balanced.'These components averaged, during
a 5-year period of observations, 9.18% and 3.72%
respectively, in relation to the total recorded in 1980.
Mortality appears to be independent of diameter size.
However, the survivors are growing at a mean rate of 0.21
cm/year in dbh.
The ratio Al/Az averaged 1.05, suggesting that this
population will approach the ”climax” state slower than at
110

111
least two mixed temperate hardwood systems, one in
Connecticut and another in New Jersey.
In terms of utilizable volume stock, the forest of the
study area,(control plots of NR experiment) is relatively
poor. The total volume including bark averaged 284 cu.m/ha
for all tree species with dbh >, 25 cm. From this total, only
about 40% fulfills the minimum size requirements (dbh > 40
cm) for Amazonian forest industries, and from this only
about 30% qualifies as economically desirable species. Thus,
the remaining utilizable volume is no more than 35 cu.m/ha.
This is much less than half of the average volume reported
for growing stock of North American temperate forests,
Brazilian temperate forests, or even tropical moist forests
in SE Asia.
In contrast, the area is very important in species
richness. About 350 different tree species from 52 different
botanical families were identified. Lecythidaceae,
Leguminosae and Sapotaceae are the dominant families
contributing more than 50% of all trees tallied.
Among the three hypothesized diameter distribution
functions tested in this study, the Weibull PERC
(distribution whose parameters were computed using the
percentile approach) showed the best fit for the observed
data. The Weibull MLE (the maximum likelihood approach) and
the exponential distributions were very sensitive to
variation within sample plots. When the Weibull shape
parameter, 0, equals 1, an exponential distribution results,

112
but even in this case, the Weibull PERC fitted the observed
data as precisely as the exponential function. In addition,
the Weibull PERC function is very simple in estimating its
parameters; it does not require sophisticated computer
capabilities.
The first-order Markov chain analysis allowed
projection of the overstory mortality and frequency
distribution by dbh classes. Since age and successive
records from long-term permanent plots are not available,
the Markov approach is a realistic alternative to predict
the direction of future trends in the study area.
Traditional growth and yield models cannot be applied
to the study area since age and site index are not
available. The 5-year increments did not show any
indications that they can be correlated with dbh. From this
study, Lotka's model appears to be a powerful alternative to
replace the traditional growth and yield models. Another
alternative is the equation, V(85) = a + b* 0(80) +
c*(D(80)f2, which presented a very good fit for the observed
data.
All mathematical models developed in this study have as
output the abstract model for a management strategy to be
implemented in the real world. In 1990, these models should
validated and, if necessary, refined based then on a 10-year
period of observations. If valid, they will be very helpful
to exercise the simulation as a means of determining model
time response for a longer period.

APPENDIX

APPENDIX
Floristic composition of bacia 3 by botanical family
(Developed by Department of Botany of INPA)
1. ANACARDIACEAE
Anacardium spruceanum Benth ex Engl.
Astronium - l sni(*)
Tapira retusa Ducke
2. ANONACEAE
Anaxagorea - l sni
Anona ambota Aubl.
Bocageopsxs multiflora (Mart.) R.E. Fr.
Bocageopsis - l sni
nguetia flagelaris Huber
Duguetia - l sni
Ephe ranthus amazonicus R.E. Fries
Ephedranthus - l sni
Quatteria olivacea R.E. Fries
Guatteria - l sni
gseudoxandra cariaceae R.E. Fries
Bollinia insignia R.E. Fries var. pallida R.E. Fries
Unono sis - l sni
Xylopia benthami R.E. Fries
Xylopia - l sni
3 . APOCYNACEAE
Ambelania acida Aubl.
Anacampta - l sni
Aspidosperma album (Vahl.) R. Ben.
Aspidosperma obscurinervius Azamb.
AspidOsperma carapanauba Pichon
Aspidosperma - l sni
gouma macrocarpa Barb. Rodr.
geissospermum ar enteum R. Rodr.
Himatanthus sucqua (Spruce) Woodson
4. ARALIACEAE
Didymopanax morototoni (Aubl.) Decne. & Planch.
* sni = species not identified for determined genus.
113

9.
114
BOMBACACEAE
Bombacopsis - 2 sni's
gatostemma milanezii Paula Nov.
§cleronema micranthum Ducke
Scleronema - l sni
BIGNONIACEAE
Jacaranda copaia D. Don.
Jacaranda - l sni
Tabebuia serratifolia (D. Don.) Nichols.
BORAGINACEAE
Cordia - 1 sni
BURSERACEAE
gemicrepidospermum rhoifolium (Bth.) Swart.
grotium aracouchili (Aubl.) March.
grotium heptaphyllum (Aubl.) March.
grotium subserratum Engler
Protium - 4 sni's
Tetra astris unifoliata (Engl.) Cuatr.
Tetragastris - 2 snI's
Trattinickia - l sni
CARYOCARACEAE
garygcar pallidum A.C. Smith
Caryocar villosum (Aubl.) Pers.
10L CELASTRACEAE
Goupia glabra Aubl.
11. CHRYSOBALANACEAE
gouepia leptostachya Benth. ex Hook
Coue ia - l sni
Birtella glandulosa Spreng.
Licania a1 a (Ben.) Cuatr.
canescens R. Ben.
Licania
Licania
Licania
Licania
fiicania
Licania
Licania
Eicania
Licania
Licania
gracilipes Taub.
heteromorphg
hypoleuca Benth.
kunthiana Hook f.
latifolia Benth. ex Hook
micrantha Miq.
oblongifalia Standl.
reticulata Prance
- l sni
Parinari montana Aubl.
12. COMBRETACEAE
Buchenavia parvifolia Ducke
Buchenavia - 2 sni s

13.
14.
15.
16.
17.
18.
19.
20.
115
CONNARACEAE
Connarus - l sni
DICBAPETALACEAE
Tapura amazonica
DUCKEODENDRACEAE
Duckeodendron cestroides Ruhlm
EBENACEAE
Diospyros bullata A.C. Smith
ELAEOCARPACEAE
Sloanea - l sni
ERYTHROXYLACEAE
Erythroxylum - lsni
EUPHORBIACEAE
Anomalocalyx - l sni
Qpnceveiba guianensis Aubl.
Qroton lanjouwensis Jablonski
Croton - l sni
erpetes variabilis Vittien
Qavarretia - 1 sni
glycidendron amazonicum Ducke
Hevea guianensis Aubl.
Mabea caudata Pax. ex K. Holhm.
Mabea - l sni
Micrandra rossiana R.E. Schultes
Micrandra siphonioides Bth.
Micrandropsis scleroxylon W. Rodr.
gausandra macropetala Ducke
Pera - 1 sni
ngonophora schomburgkiana Miers. ex Bth.
FLACOURTIACEAE
gasearia combaymensig Tul.
gasearia ulmifolia Vahl. ex Von.
Casearia - l sni
Carpotroche - l sni
Laetia procera (Poepp.) Eichl.____,_
Ryania - l sni
21. GUTTIFERAE
Qalophyllum brasiliense Camb.
Carai a - l sni
Clusia - l sni
Havetiopsis - l sni
Moronobea coccinea Aubl.
Moronobea pulchra Ducke
Rheedia 2 - sni‘s
Symphonia globulifera Linn
V smia uckei Maguire

22.
23.
24.
25.
116
Vismia guianensis (Aubl. ) Choisy
Tovomita - l sni
HIPPOCRATEACEAE
Salacia - l sni
HUMIRIACEAE
Duckesia verrucosa (Ducke) Cuatr.
Endopleura uchi (Huber) Cuatr.
Humiria balsam1fera (Aubl. ) St. Hill
Sacoglottis ceratocarpa Ducke
Sacaglott1s - l sni
Vantanea macrocarpa Ducke
Vantanea parviflora Lam.
Vantanea - l sni
ICACINACEAE
Emmotum - 2 sni's
Poraqueiba - l sni
LAURACEAE
Aniba canelilla (B. B. K. ) Mez.
Aniba duckei Kosterm.
Aniba ferrea Kubitzki
Aniba rosaedora Ducke
Aniba terminalis Ducke
Aniba - l sni
Endlicheria - 4 sni's
Licaria canela (Meissn. ) Kosterm.
Licaria guianensis Aublet
Licaria rigida Kost.
Licaria - 3 sni' s
Mezilaurus decurrens (Ducke) Kost.
Mezilaurus synandra (Mez.) Kosterm.
Mezilaurus - l sni
Nectandra rubra (Mez.) C.K. Allen
Nectandra - l sni
Qcotea canaliculata Mez.
Ocotea neesiana (Miq.) Kosterm.
Ocotea - 9 sni's
26. LECYTHIDACEAE
Cariniana decandra Ducke
Car1niana micrantha Ducke
Corytophora alta Knuth
Corytophora rimosa Rodr.
Couratari - l sni
_schweiIera fracta R. Knuth
§schweilera odora (Poepp.) Miers.
Eschweilera - 8 sni's
Gustavia au usta L.
GustavIa elliptica Mori
Holopyx1dium latifolium (A. c. Smith) R. Knuth.
Lecythis usitata Miers var. paraensis R. Knuth.

117
27. LEGUMINOSAE CAESALPINIODEAE
Aldina hetero h lla Spruce ex Bth.
Bocoa viridirora (Ducke) Cowan
Cassia rubriflora Ducke
Copaifera multijug_ Hayne
Elizabetha bicolor Ducke
Elizabetha princeps Schomb. ex Bth.
Elizabetha - l sni
Eperua bijug_ Mart. ex Benth. var. glabriflora Ducke
§pgrua duckeana Cowan
Eperua schomburgkiana Benth.
Hymenaea pArvifolia Huber
Hymenaea - 4 sni' s
Macrolobium limbatum Spr. ex Benth.
Macrolobium microcalyx Ducke
Peltogyne cat1nggg subsp. labra (W. Rodr. ) M. F.Silva
Peltogyne paniculata subsp. pAniculata Benth.
Swartzia ingifolia Ducke
Swartzia pgnacoco (Aubl. ) Cowan
§wartzia polyphyl1g D.C.
§wartz1a recurva Poepp. & Endl.
Swartzia reticglata Ducke
Swartzia ulei Harms
Swartzia - 3 sni's
§g1erolobium - l sni
Vouacapoua pallidior Ducke
Tachigalia myrmecophilla (Ducke) Ducke
TAch1gAl1a pAn1culata Aubl.
Tach1gA11a - l sni
28. LEGUMINOSAE MIMOSOIDEAE
Dimorphandra parviflora Spr. ex Bth.
Diniz1a excelsa Ducke
Enterolobium schomburgkii Benth.
Hymenolobium - l sni
Inga aff. brevialata Ducke
Inga paraensis Ducke
Inga cayennensis Benth.
Inga - 4 sn1' s
Parkia multiju a Bth.
Parkia opposit olia Spr. ex Bth.
Parkia pendula Benth. ex Walp.
Parkia - 2 sn '3
Piptadenia psilostachya (D. C. ) Bth.
Piptaden1a suaveolens Miq.
g1ptadenia - 1 sni
Eithecolobium racemosum Ducke
Pithecolobium - 2 sni‘s
Sgryphnodendron racemiferum (Ducke) W. Rodr.
Stryphnodendron - l sni
29. LEGUMINOSAE PAPILIONOIDEAE
Andira parviflora Ducke

30.
31.
32.
33.
34.
118
Andira unifoliata Ducke
Andira - l sni
DIpteryx alata Vogel
Dipteryx ma nifica Ducke
Dipteryx odorata (Aubl. ) Willd.
DApteryx oppositifolia (Aubl. ) Willd.
Dipteryx polyphylla (Ducke) Hub.
fiipteryx - 1 sni
gymenolobium sericeum Ducke
gymenolobium cf. pulcherrimum Ducke
gymenolobium - l sni
Ormosia sm1thii Rudd.
Diplotrop1s - l sni
Platymiscium duckei Huber
LINACEAE
Roucheria callophylla Planch
MALPIGHIACEAE
gyrsonima stipulacea Adr. Juss.
B rsonima - l sni
Pterandra arborea Ducke
MELASTOMATACEAE
Bellucia grossularioides (L. ) Triana
Miconia elaeagnoides Cogn.
Micon1a re e111 Cogn.
Mouriria angulicosta Morley
MourirIa - l sn1
MELIACEAE
Guarea - 2 sni's
Trichilia - 2 sni's-
MORACEAE
Brosimum guianensis Aubl.
Brosimum pgtabile Ducke
Brosimum pgr1nar1oides Ducke subsp. parinarioides
Brosimum utile (H. B. K. ) Pittier
Brosimu rubescens Taub.
Cecropia scyadophylla Mart. var. juranyana Snethlage
Claris1a racemosa R.et P.
Cousapoua - 1 sni
Pcus clusiaefolia Schott
F1cus guianensis Desv.
HelicostyAis - l sni
Maggira calophylla (P.A.E.) C.C. Berg.
Magu1ra sclerophylla (Ducke) C. C. Berg.
NaucleOpsis caloneura (Hub. ) Ducke
Naucleopsis glabra Spruce ex Baill
Naucleopsis macrophylla Miq.
Perebea mollis (P. E. ) Huber subsp. mollis
Perebea mollis (P. S. C. ) Huber
Pourouma ovata Trecul.

35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
119
Pseudolmedia - 1 sni
Sorocea - 1 sni
MYRISTICACEAE
gpgpsoneura ulei Warb.
lryanthera - 1 sni
Osteophloeum lat s ermum (A.D.C.) warb.
Virola calophylla Mgf.
girola carinata (Bth.) Warb.
ViroIa elon ata (Bth.) Warb.
Virola c . m1chelii Beckel
girola multinervia Ducke
girola pavonis (A.D.C.) Smith
girola venosa Warb.
Virola venosa (Benth.) Warb.
MYRTACEAE
Eu enia - 3 sni's
M rc1a ma na Legrand
Mxrc1a a ax (Rich.) D.C.
MONIMIACEAE
Siparuna dicipiens (Tu1.) A.D.C.
NYCTAGINACEAE
Neea cf. altissima P. et E.
Neea - 2 sni‘s
OCHNACEAE
Ouratea discophora Ducke
Ouratea - 1 sni
OLACACEAE
Agtandra - 1 ani
Qhaunochiton - 2 sni's
Beisteria acumitetg (B.B.) Engl.
BEISteria barbata Cuatr. .
Beisteria - 2 sni's
ginquartia guianensis Aubl.
Ptychopetalum olacoides Benth.
PROTEACEAE
Rougala - 1 sni
QUIINACEAE
Quiina abovata Tul.
Quiina - 1 sni
Touro ia guianensis Aubl.
RHABDENDRACEAE
Rhabdodendron amazonicum (Spr. ex Bth.) Bub.
RBIZOPHORACEAE
Anisophyllea manausensis Pirea 5 W. Rodr.

120
Sterigmapetalum obovatum Kuhlmann
4S. RUBIACEAE
Amaioua - 1 sni
Duroia fusifera Hook f. ex K. Schum
Duroia - 1 sni
Elaeagia - 1 sni
Faramea - 1 sni
Perainandusa - 2 sni' s
PTlicourea anisoloba M. Arg.
Palicourea cf. leggiflora (Aubl. ) A. Rich.
PTgamea - 1 sni
Psychotria prancei Steyermark
Remijia - 3 sni' s
46. SAPINDACEAE
Mata ba - 1 sni
M1cropholis - 1 sni
Talis1a - 1 sni
Toulicia - 1 ani
47. SAPOTACEAE
Achrouteria Egmifera Eyma
Achrouter1a - 2 sn1 s
Chrysophyllum oppositum (Ducke) Ducke
Chrysophyllum anomalum Piree
Diplocem venezuelana Aubr.
Ecclinusa Bacuri AfiBr. et Pellegr.
Ecclinusa ucugu1rana Aubr. 5 Pellegr.
Ecclinusa - 2 sn1 s
Pranchetella platyphylla (A. C. Sm.) Aubr.
Pranchetella - 1 sni
Glycoxylon pedicellatum (Ducke) Ducke
Lafiétia - 4 sni' s
Manilkara amazonica (Huber) Standley
Manilkara huberi (Ducke) Chev.
Manilkara cavalcantei Pires et Rodr.
Manilkara surinamenEis (Miq.) Dubard
Micropholis truncifiora Ducke
Micropholis guyanensis Pierre
Micropholis venulosa Pierre
Micropholis rosaainha-brava Aubr. et Pellegr.
Microphol1s mensalis (BTehni) Aubr.
Microphol1s - S sni' s
Myrtiluma eu eniifolia (Pierre) Baill
Neoxxthecec uaantha (Sandw.) Aubr.
Pouteria ggyanensis Aubl. L. O.A. Teixeira 82
Pradosia verticillata Ducke
gr1eure11a manaosens1s Aubr.
Pseudolabatié - 1 sni
RafilkoEerelIa - 1 sni
Ra ala spuria (Ducke) Aubr.
R1charde11a manaosensis Aubr. et Pellegr.
Richardella macrophylla (Lum.) Aubr.

48.
49.
SO.
51.
52.
'53.
54.
121
szxgiopsis oppositifolia Ducke
SarcauIis brasiliensis (A.D.C.) Eyma
SIHARUBACEAE
§imaruba amara Aubl.
Simaba guianensis Aubl. subsp. guianensis
SImaEa cuspidata Spruce
STERCULIACEAE
§terculia speciosa K. Schum.
Sterculia - 1 sni
TheoEroma sylvestris Aubl. ex Mart.
STYRACACEAE
Stxrax - 1 sni
TILIACEAE
Apeiba echinata Gaertn.
Apeiba burchelii Sprague
Luehea - 1 ani
VERBENACEAE
Vitex triflora Vahl.
VIOLACEAE
geonia glycicarpa Ruiz et Pav.
ginorea guianensis Aubl. var. subintegrifolia
Rinorea racemosa (Mart. et Zucc.) O. Ktze.
égphirshox sufihamensis Eichl.
VOCHYSIACEAE
Erisma bicolor Ducke
Erisma fuscum Ducke
Qualea clavata Staflen
Qualea paraensis Ducke
QuaIea cassiquiarensig (Spr.) Warm.
Qgglea labourianana Paula
nglea brevipediceilata Staflen
Vochysia obiflénsis (355.) Ducke

LIST 01? REFERENCES

LIST OF REFERENCES
Acevedo, M.F. 1981. On Born's Markovian Model of Forest
Dynamics With Particular Reference to Tropical
Forests. Theoretical Population Biology 19:230-250.
Alder, D. 1979. A Distance-independent Tree Model for
Exotic Conifer Plantations in East Africa. Forest Sci.
25:59-71.
Alder, D. 1980. Forest Volume Estimation and Yield
Prediction. Vt2 - Yield Prediction. FAD Forestry Paper
22/2. 194p.
Alencar, ch. and qu. Araujo. 1981. Incremento Anual do
Pau-rosa (Aniba duckei Kostermans) Lauraceae, em
Floresta Tropical Umida Primaria. Acta Amazonica
11(3):547-552.
Anderson, T.W. and INA. Goodman. 1957. Statistical
Inference About Markov Chains. Annals of Mathematical
Statistics 28:89-110.
Anton, B. 1973. Elementary Linear Algebra. John Wiley and
Sons, Inc. New York. 296p.
Bailey, R.L. and T.R. Dell. 1973. Quantifying Diameter
Distributions With the Weibull Function. Forest Sci.
19:97-104.
Barros, P.L.C., S.A. Machado, D. Burger, and J.D.P.
Siqueira. 1979. Comparacao de Modelos Descritivos da
Distribuicao Diametrica em uma Floresta Tropical.
Floresta lO(2):l9-32.
Bell, 6.8. 1976. Notes on Natural Regeneration in Dominica.
Windward Islands. Commonw. For. Rev. 55(l)27-36.
Bellefleur, P. 1981. Markov Models of Forest-type Secondary
Succession in Coastal British Columbia. Can. J. For.
RES. 11:18-29.
Bierzychudek, P. 1982. The Demography of Jack-in-the-
pulpit, a Forest Perennial That Changes Sex. Ecol.
Monographs 52(4)::335-351.
Braga, IhI.S. 1979. Subdivisao Fitogeografica, Tipos de
Vegetacao, Conservacao e Inventario Floristico da
122

123
Floresta Amazonica. Acta Amazonica 9(4):53-80.
Britwum, S.P.K. 1976. Natural and Artificial Regeneration
Practices in the High Forest of Ghana. Ghana Forestry
Journal 2:45-49.
Bruner, 3.0. and J.W. Moser Jr. 1973. A Markov Chain
Approach to the Prediction of Diameter Distributions
in Uneven-aged Forest Stands. Can. J. For. Res. :409-
417.
Brunig, E.F. 1977. The Tropical Rain Forest - A Wasted
Asset or an Essential Biosphere Resource? Ambio 6(4).
Budowski, G. 1976a. Why Save Tropical Rain Forests? Some
Arguments for Campaigning Conservationists. Amazoniana
5(4):529-538.
Budowski, G. 1976b. Los recursos naturales en el desarrolo
del Tropico Bumedo. Reunion sobre desarrolo regional
del Tropico Americano. IICA-Tropicos. Belem, Para,
Brazil.
Buogiorno, J. and B.C. Michie. 1980. A Matrix Model of
Uneven-Aged Forest Management. Forest Sci. 26:609-625.
Chai, D.N.P. and M.P. Udarbp. 1977. The Effectiveness of
Current Silvicultural Practice in Sabah. The Malaysian
Forester 40(1):27-35.
Cheah, INC. 1978. Forest regeneration and development
options in Peninsular Malaysia today. The Malaysian
Forester 41(2):17l-l75.
Chiang, CAL. 1980. An Introduction to Stochastic Processes
and Their Applications. Robert E. Krieger Publ. Co.
Huntington, New York.
Clutter, £LL. 1963. Compatible Growth and Yield Models for
Loblolly Pine. Forest Sci. 9:354-371.
Clutter, J.L., J.C. Fortson, L.V. Pienaar, G.B. Brister,
and R.L. Bailey. 1983. Timber Management: A
Quantitative Approach. John Wiley and Sons, Inc. New
York. 333p.
Cochrane, T.T. and P.A. Sanchez. 1980. Land Resources,
Soils and Their Management in the Amazon Region: A
State of Knowledge Report. In: Amazonia - Agriculture
and Land Use Research - proceedings of the
International Conference at CIAT in Cali, Colombia.
Cohen, AnC. 1965. Maximum Likelihood Estimation in the

124
Weibull Distribution Based on Complete and on Censored
Samples. Technometrics 7(4):579-588.
Conover, WuJ. 1980. Practical Nonparametric Statistics.
John Wiley and Sons, Inc. New York. 2ed. 493p.
De Graaf, N.R. 1981. Sustained Timber Production in the
South American Tropical Rainforest. Paper prepared for
the workshop on Management of low fertility acid soils
of the Humid American Tropics in Paramaribo. (copy).
Departamento de Silvicultura Tropical. 1982. Inventario
Florestal no Rio Trombetas (INPA/SBELL/ALCOA).
Technical Report. 49p.
Departamento de Silvicultura Tropical. 1983. Inventario
Florestal da UBE de Santa Izabel. Technical Report.
35p.
Departamento de Silvicultura Tropical. 1983. Inventario
Florestal do Projeto Rio Arinos. Technical Report.
64p.
Departamento de Silvicultura Tropical. 1983. Inventario
Florestal da UBE de Balbina. Technical Report. 111p.
Draper, NzR. and B. Smith. 1981. Applied Regression
Analysis. John Wiley and Sons, Inc. New York. 2ed.
709p.
Einsesmith, S.P. 1985. PLOTIT: User's Guide.
Enright, N. and J. Ogden. 1979. Applications of Transition
Matrix Models in Forest Dynamics: Araucaria in Papua
New Guinea and Nothofa us in New ZeTland. Australian
Journal of Ecology 4:3-g3.
Ferguson, I.S. and J3W. Leech. 1978. Generalized Least
Squares Estimation of Yield Functions. Forest Sci. 24:
27-42.
Fox, J.E.D. 1976. Constraints on the natural regeneration
of tropical moist forest. Forest Ecology Management
1:37-65.
FUA - Fundacao Universidade do Amazonas. 1979. Proposta de
Politica Florestal para a Amazonia Brasileira. Metro
Cubico - Livros e Revistas Tecnicas Ltda. Manaus, Am.
72p.
Furnival, G.M. 1961. An Index for Comparing Equations Used
in Constructing Volume Tables. Forest Sci. 7:337-361.
Gomez-Pompa, A., c. Vazquez-Yanes, and S. Guevara. 1972.

125
The Tropical Rain Forest: A Nonrenewable Resource.
Science 177:762-765.
Grossman, 8.1. and J.E. Turner. 1974. Mathematics for the
Biological Sciences. Macmillan Publishing Co., Inc.
New York. 512p.
Higuchi, N. 1981a. A silvicultura no INPA. Supl. Acta
Amazonica 11(1):99-107.
Higuchi, N. 1981b. Relatorio sobre o Encontro da IUFRO nas
Filipinas e ”study-tour” por Filipinas, Sarawak e
Fiji. Report not published.
Higuchi, N., F.C.S. Jardim, J. Santos, A.P. Barbosa, and
T.W.W. Wood. 1983. Inventario Florestal Comercial da
Bacia 3. Acta Amazonica (in press).
Higuchi, N., F.C.S. Jardim, J.Santos, and J.C. Alencar.
1985. Bacia 3 - Inventario Diagnostico da Regeneracao
Natural. Acta Amazonica 15(1-2):199-233.
Higuchi, N. and C.W. Ramm. 1985. Developing Bole Wood
Volume Equations for a Group of Tree Species of
Central Amazon (Brazil). Commonw. For. Rev. 64(1):33-
41.
Hilton, R.G.B. and R.J. Johns. 1984. The future of forestry
in Papua New Guinea. Commonw. For. Rev. 63(2):103-106.
Horn, H.S. 1975. Markovian Properties of Forest Succession.
In: Ecology and Evolution of Communities (M. Cody and
J. Diamond, eds.) pp. 196-211. Harvard University
Press. Cambridge, Mass.
Hosokawa, R.T. 1981. Manejo de Florestas Tropicais Umidas
em Regime de Rendimento Sustentado. UFPr. Technical
Report.
Hutchinson, I.D. 1980. Liberation Thinning: A Tool in the
Management of Mixed Dipterocarp Forest in Sarawak.
UNDP/FAO Field Document 8 5. Kuching.
IBGE. 1977. Geografia do Brasil. Sergraf—IBGE. Vol. 1 and
4.
INPA - Instituto Nacional de Pesquisas na Amazonia. 1979.
Estrategias Para a Politica Florestal na Amazonia
Brasileira. Suplemento Acta Amazonica 9(4). 216p.
INPA - Instituto Nacional de Pesquisas na Amazonia. 1982.
Bacia Modelo: Caracterizacao Ecologica de uma Bacia
Hidrografica Experimental na Amazonia Central.
Suplemento Acta Amazonica 12(3). 114p.

126
James, L.M. (pers. comm.). Notes of FOR 454 - Forestry in
International Development. Winter/1984.
Johnson, N.E. 1976. Biological opportunities and risks
associated with fast-growing plantation in the
Tropics. Journal of Forestry, Apri1:206-211.
Kio, P.R.o. 1976. What future for natural regeneration
of tropical high forest? An appraisal with examples
from Nigeria and Uganda. Commonwealth Forestry Review
55(4):309-318.
Kio, P.R.O. 1979. Management Strategies in the Natural
Tropical High Forest. Forest Ecology and Management
2:207-220.
Lanly, J.P. 1983. Assessment of the Forest Resources of the
Tropics. Forestry Abstracts (Review Article) 44(6):287-
318.
Lawrence, K.D. and D.R. Shier. 1981. A Comparison of Least
Squares and Least Absolute Deviation Regression Models
for Estimating Weibull Parameters. Commun. Statist. -
Simula. Computa. BlO(3):315-326.
Lee, H.S. 1982. Silvicultural Management in Sarawak. The
Malaysian Forester 45(2):485-496.
Leslie, A. 1977. Where Contradictory Theory and Practice
Co-exist. Unasylva 29(15):2-18.
Little, S.N. 1983. Weibull Diameter Distributions for Mixed
Stands of Western Conifers. Can. J. For. Res. 13: 85-
88.
Lisboa, P.L. 1975. Observacoes Sobre a Vegetacao das
Campinas Amazonicas. Acta Amazonica 5(3):211-223.
Loetsch, F., F. Zohrer, and K.E. Haller. 1973. Forest
Inventory. BLV Verlagsgesellschaft mbH, Munchen. v.2.
Lowe, R.G. 1978. Experience with the Tropical Shelterwood
System on regeneration in natural forest in Nigeria.
Forest Ecology Management 1:193-212.
Miller, T.B. 1981. Growth and Yield of Logged-over Mixed
Dipterocarp Forest in East Kalimantan. The Malayasian
For. 44(2):419-424.
Morrison, D.F. 1976. Multivariate Statistical Methods.
McGraw-Hill Inc. 415p.
Murphy. P.G. (pers. comm.). Notes of BOT 441 - Geographical

127
Plant Ecology. Winter/1985.
Myers, N. 1982. Depletion of Tropical Moist Forests: A
Comparative Review of Rates and Causes in Three Main
Regions. Acta Amazonica 12(4):745-758.
Myers, N. 1983. Tropical moist forests: over-exploited
and under-utilized? Forest Ecology Management 6:59-
79.
Palmer, J.R. 1977. Forestry in Brazil - Amazonia. Commonw.
For. Rev. 56(2):115-l29.
Peden, L.M., J.S. Williams, and W.E. Frayer. 1973. A Markov
Model for Stand Projection. Forest Sci. 19:303-314.
Pielou, ELC. 1977. Mathematical Ecology. John Wiley and
Sons. New York.
Prance, G.T. 1974. Phytogeographic Support for the Theory
of Pleistocene Forest Refuges in the Amazon Basin,
Based on Evidence from Distribution Patterns in
Caryocaraceae, Chrysobalanaceae, Dichapetalaceae and
Lecythidaceae. Acta Amazonica 3(3):5-28.
Prance, G.T. 1980. A Terminologia dos Tipos de Florestas
Amazonicas Sujeitas a Inundacao. Acta Amazonica 10(3):
495-504.
Rai, S.N. 1979. Diameter/Height, and Diameter/Girth
Relationship of Some Rain Forest Tree Species of
Karnataka - India. The Malayasian For. 42(1):53-58.
Ranzani, G. 1979. Recursos Pedologicos da Amazonia. Acta
Amazonica 9(4):23-35.
Ranzani, G. 1980. Identificacao e Caracterizacao de Alguns
Solos da EEST. Acta Amazonica 10(1):7-41.
Revilla,.AJL 1981. A Review of Growth and Yield Studies on
the Tropical Mixed Forests in the Asia Region. Paper
for the IUFRO meeting on ”Growth and Yield Studies on
the Tropical Mixed Forests”, in Los Banos, the
Philippines.
Ribeiro, M.N.G. 1976. Aspectos Climatologicos de Manaus.
Acta Amazonica 6(2):229-233.
Salati, E. and P.B. Vose. 1984. Amazon Basin: A System in
Equilibrium. Science 225:129-138.
Schubart, H.O.R. and E. Salati. 1980. Natural Resources for
Land Use in the Amazon: The Natural Systems. In:
Amazonia -- Agriculture and Land Use Research

128
Proceedings. Cali, Colombia. 211-259.
Schreuder, H.T., W.L. Hafley, and P.A. Bennett. 1979. Yield
Predictions for Unthinned Natural Slash Pine Stands.
Forest Sci. 25:25-30.
Sedjo, R.A. and M. Clawson. 1983. How Serious is Tropical
Deforestation. Journal of Forestry Dec.:792-794.
Shugart,‘HJL 1984. A Theory of Forest Dynamics: The
Ecological Forest Succession Models. Springer-Verlag
Inc. New York. 278p.
Shugart, H.H. and D.C. West. 1981. Long-term Dynamics of
Forest Ecosystems. Am. Scientist 69:647-652.
Silva, M.P., P.L. Lisboa, and R.C.L. Lisboa. Nomes Vulgares
de Plantas Amazonicas. INPA. 222p.
Smith, V.G. 1983. Compatible Basal Area Growth and Yield
Models Consistent With Forest Growth Theory. Forest
Sci. 29:279-288
Soekotjo and 0.1. Dickmann. 1978. The Effect of Indonesian
Selective Logging on Natural Regeneration in the East
Kalimantan Rain Forest. Paper for Eighth World
Forestry Congress in Jakarta, Indonesia.
Soekotjo. 1981. Diameter Growth of Residual Stands in
Logged Over Areas in East Kalimantan Tropical Rain
Forest Indonesia. PhD Dissertation Submitted to
Department of Forestry - Michigan State University.
Spears, J.S. 1979. Can the Wet Tropical Forest Survive?
Commonw. For. Rev. 58(3):165-180.
Sullivan, AAD. and JuL. Clutter. 1972. A Simultaneous
Growth and Yield Model for Loblolly Pine. Forest Sci.
18:76-86.
Tang, H.T. 1980. Factors affecting regeneration methods
for Tropical high forests in South East Asia. The
Malaysian Forester 43(4):469-480.
Tang, H.T. and W.R. Mohd. 1981. Growth and Yield Studies in
Inland Mixed Indigenous Forests in Peninsular
Malayasia. Paper for IUFRO meeting "Growth and Yield
Studies on the Tropical Mixed Forests“, in Los Banos,
the Philippines.
Tosi, J. 1983. Provisional Ecological Map of the Republic
of Brazil. Institute of Tropical Forestry; San Jose,
Costa Rica.

129
Usher, M.B. 1966. A Matrix Approach to the Management of
Renewable Resources, With Special Reference to
Selection Forests. Journal of Applied Ecology 3(2):
355-367.
Usher, M.M. 1969. A Matrix Model for Forest Management.
Biometrics June:309-315.
Usher, M.B. 1979. Markovian Approaches to Ecological
Succession. Journal of Animal Ecology 48:413-426.
Virtucio, F.D. and M.G. Torres. 1978. Status of Management
and Utilization of Forest Resources in the
Philippines. The Malaysian Forester 41(2):149-166.
Volatron, B. 1976. La Mise en Valeur des Richesses
Forestieres en Amazon Bresilienne et en Colombie.
Revue Bois et Forets des Tropiques 165(1):59-76.
Wadsworth, F.H. 1970. Review of Past Research in the
Luquillo Mountains of Puerto Rico. In: A Tropical Rain
Forest. Odum, H.T. and R.F. Pigeon (eds). pp. B-33 to
8.46.
Walter, H. 1973. Vegetation of the Earth and Ecological
Systems of the Geo-biosphere. Springer-Verlag New York
Inc. 2ed. 274p.
Weaver, PJL 1982. Tree Diameter Increment in Upper Montane
Forest of Puerto Rico. Turrialba 32(2)119-122.
West, P.W. 1980. Use of Diameter Increment and Basal Area
Increment in Tree Growth Studies. Can. J. For. Res.
10:71-77.
West, P.W. 1981. Simulation of Diameter Growth and
Mortality in Regrowth Eucalypt Forest of Southern
Tasmania. Forest Sci. 27:603-616.
Wood, T.W.W. 1985. (Personal communication). British Aid
Program with Honduran Forest Service. Tegucigalpa,
Honduras.
Zarnoch, S.J., C.W. Ramm, V.J. Rudolph, and M.W. Day. 1982.
The Effects of Red Pine Thinning Regimes on Diameter
Distribution Fitted to Weibull Function. MSU
Agricultural Experiment Station East Lansing. RI-423.
11p.
Zarnoch, S.J. and T.R. Dell. 1985. An Evaluation of
Percentile and Maximum Likelihood Estimators of
Weibull Parameters. Forest Sci. 31:260-268.

"‘WEHIIHEHJHLTMTLMW‘S


















