
1 stranded RNA bacteriophages, C-1 and Hgal1: rethi - American
34
1 Genomic sequences and characterizations of two R-plasmid-dependent single- 1 stranded RNA bacteriophages, C-1 and Hgal1: rethinking the evolution of ssRNA 2 phages 3 4 Running title: ssRNA phages C-1 and Hgal1 genomes 5 6 Sherin Kannoly, Yongping Shao*, and Ing-Nang Wang # 7 8 Department of Biological Sciences, University at Albany 9 *Current address: Department of Cancer Biology and Kimmel Cancer Center, Thomas 10 Jefferson University, 233 South 10th Street, Philadelphia, PA 19107 11 # Corresponding author: [email protected] 12 13 14 15 Copyright © 2012, American Society for Microbiology. All Rights Reserved. J. Bacteriol. doi:10.1128/JB.00929-12 JB Accepts, published online ahead of print on 20 July 2012 on January 1, 2019 by guest http://jb.asm.org/ Downloaded from
Transcript of 1 stranded RNA bacteriophages, C-1 and Hgal1: rethi - American

1
Genomic sequences and characterizations of two R-plasmid-dependent single-1 stranded RNA bacteriophages, C-1 and Hgal1: rethinking the evolution of ssRNA 2
phages 3 4 Running title: ssRNA phages C-1 and Hgal1 genomes 5 6 Sherin Kannoly, Yongping Shao*, and Ing-Nang Wang# 7 8 Department of Biological Sciences, University at Albany 9 *Current address: Department of Cancer Biology and Kimmel Cancer Center, Thomas 10
Jefferson University, 233 South 10th Street, Philadelphia, PA 19107 11 # Corresponding author: [email protected] 12 13 14 15
Copyright © 2012, American Society for Microbiology. All Rights Reserved.J. Bacteriol. doi:10.1128/JB.00929-12 JB Accepts, published online ahead of print on 20 July 2012
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

2
Abstract 16 We have sequenced and characterized two R-plasmid-dependent single-stranded 17 RNA bacteriophages (RPD ssRNA phages), C-1 and Hagl1. Phage C-1 requires a 18 conjugative plasmid of the IncC group, while Hgal1 requires the IncH group. Both the 19 adsorption rate constants and one-step growth curves are determined for both phages. 20 We also empirically confirmed the lysis function of the predicted lysis genes. Genomic 21 sequencing and phylogenetic analyses showed that both phages belong to the Levivirus 22 group and are most closely related to another IncP-plasmid-dependent ssRNA phage, 23 PRR1. Furthermore, our result strongly suggests that the stereotypical bauplans of 24 genome organization found in the Levivirus and Allolevivirus predate phage 25 specialization on conjugative plasmids, suggesting that the utilization of conjugative 26 plasmids for cell attachment and entry are independent evolutionary events for these two 27 main clades of ssRNA phages. Our result is also consistent with a previous study, 28 making the Levivirus-like genome organization ancestral and the Allolevivirus-like 29 genome derived. To obtain a deeper insight into the evolution of ssRNA phages, more 30 phages specializing on various conjugative plasmids and infecting different bacterial 31 species would be needed. 32 33 34
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

3
Introduction 35 The single-stranded RNA bacteriophage (ssRNA phage) is the simplest and 36 smallest phage, containing only three or four genes (58, 63). While these phages have 37 traditionally been classified into four subgroups, I, II, III, and IV, based on their 38 serological relationship (53, 60), genomic data have collapsed these subgroups into two 39 genera: Levivirus, which is composed of subgroups I and II, and Allolevivirus, which 40 includes subgroups III and IV (41). A recent phylogenetic study with 30 coliphages 41 further supports such a classification scheme (25). 42 The bulk of our knowledge about the ssRNA phages is mainly derived from F-43 plasmid-dependent (FPD) phages infecting mainly Escherichia coli. This is true for the 44 numbers of genomes sequenced, of molecular studies (58, and reference therein, 63), and 45 of surveys of ecological distribution (12, 27-29, 32, 47, 48, 54). However, many 46 unassigned phages listed in the sixth report of the International Committee on Taxonomy 47 of Viruses (41) are not F-plasmid dependent. With few exceptions, such as PP7 (10, 46), 48 PRR1 (45, 51), AP205 (38), and φCb5 (37), not much is known about these reported, but 49 not-yet sequenced phages. 50 Many of these unassigned phages are isolated by their dependence on pili 51 produced by conjugative R (resistance) plasmids, which are classified based on their 52 memberships in various incompatibility (Inc) groups (13, 19, 20, 61). In fact, many are 53 named after the Inc groups of these plasmids from which they were first isolated with 54 (11, 14-18, 43, 45, 55). When compared to the more familiar FPD ssRNA phages, these 55 R-plasmid-dependent (RPD) ssRNA phages have a much wider host range, 56 encompassing many species in Enterobacteriaceae, presumably due to the promiscuity of 57
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

4
the corresponding R plasmids. Interestingly, while all can propagate (multiply) in the 58 presence of the bacterial hosts with the required R plasmids, not all can plaque on the 59 same propagation hosts. Morphologically speaking, these reported RPD ssRNA phages 60 are all similar in size to the FPD phages, presumably as an icosahedron virion. While all 61 phages, with electron microscopy, are shown to attach to the shaft part of the R pili, not 62 all distributions are the same, with some (e.g., phages D, F0 lac, and t) tending to attach to 63 the shaft parts that are close to the tip. Furthermore, some phages (e.g., phages D, M, 64 pilHα, and SR) are somewhat sensitive to the presence of chloroform, a conventional test 65 for differentiating between plasmid-dependent ssRNA phages (usually chloroform-66 insensitive) and ssDNA filamentous phages (usually chloroform-sensitive). So far, the 67 one common characteristic uniting all these ssRNA phages together is their sensitivity to 68 RNase during infection. All information can be found in the publications cited above. 69 Apparently, there is a certain level of variation among these ssRNA phages that 70 knowledge gleamed from studying the FPD ssRNAs alone is incomplete in capturing the 71 diversity of this very simple group of viruses. 72 In this study, we have obtained phages C-1 and Hgal1 from the National 73 Collection of Type Cultures (NCTC, London, UK), presumably deposited by the 74 researchers who first isolated these phages. Phage C-1 was originally isolated from 75 sewage samples in Pretoria, South Africa (55), and phage Hgal1, also isolated from raw 76 sewage samples collected from the sea outfall in Galway docks, Ireland (43). Since their 77 initial isolation and characterization, no further work has been performed. Our study 78 represents a first attempt after many decades of obscurity for these phages. 79 80
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

5
Materials and Methods 81 Phage and bacterial strains, primers, and growth medium. All phage and 82 bacterial strains are listed in Table 1. Primers for PCR amplification, cloning, and DNA 83 sequencing are listed in Supplemental Material. Both phages C-1 and Hgal1, strain 84 numbers NC50385-01 and NC50402-01, respectively, were obtained from the National 85 Collection of Type Cultures (NCTC), London, UK. The host strains used to propagate 86 phages C-1 and Hgal1 were obtained from the Félix d´Hérelle Reference Center for 87 Bacterial Viruses (Laval University, Québec City, Canada), strain numbers HER1222 and 88 HER1240, respectively. HER1222 is originally E. coli K12 JE-1 and carries an IncC 89 group plasmid, RA1 with Tn5 insertion (a more detailed information can be found at 90 http://www.phage.ulaval.ca/en/search_catalog/). HER1240 is originally E. coli K12 J62 91 and carries an IncHI1 group plasmid, R27 with Tn7 insertion. 92 Throughout this study, LB medium and double-agar overlay plating using LB-93 agar plates were used. Antibiotics, when appropriate, were added to the final 94 concentrations of 100 μg/mL for ampicillin, 40 μg/mL for kanamycin, and 10 μg/mL for 95 tetracycline. 96 Purification of phages. The host bacteria (1 L) were grown to exponential phase 97 of OD550 ≈ 0.1 - 0.2, then infected with phages at an m.o.i. of 0.1 for 16 h, all at 30 ºC. 98 To the lysate, 20 units of bovine pancreatic DNase I (Roche) was added and incubated at 99 room temperature for 30 min, followed by addition of 0.5 M NaCl and incubated on ice 100 for 1 h. After centrifugation at 11,000 g for 10 min at 4 ºC, the phage in the cleared 101 lysate was precipitated by adding polyethylene glycol (mol. wt. 8000, Sigma) at a final 102 concentration of 11%. After centrifugation at 11,000 g, the precipitate was resuspended 103
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

6
with 7.5 mL of SM buffer (5.8 g NaCl, 2 g MgSO4⋅7H2O, 50 mL 1 M Tris-HCl (pH 7.5), 104 5 mL 2% gelatin solution in 1 L water) and mixed with equal volume of chloroform, then 105 centrifuged at 4,000 g for 15 min at 4 ºC. The top layer was collected and added with 106 CsCl to a final concentration of 0.75 g/mL for ultracentrifugation at 36,000 rpm (SW-40 107 rotor, Beckman) for 24 h at 4 ºC. For phage C-1, the titer prior to purification was ∼1011 108 PFU/mL and we recovered ∼1013 total PFU (10% recovery). For phage Hgal1, the 109 numbers are ∼1011 PFU/mL and ∼3×1013 total for the before and after figures, 110 respectively (∼30% recovery). 111 Phage genome cDNA synthesis, cloning, and sequencing. Approximately 1013 112 PFUs were used for RNA genome isolation. RNA was isolated by phenol-chloroform 113 extraction and ethanol precipitation. The phage RNA was tailed with ATP at the 3' end 114 using a poly(A) tailing kit (Ambion), following the manufacturer’s recommendation. 115 The tailed RNA was reverse transcribed using an oligo(dT) primer and AMV reverse 116 transcriptase (Fermentas) as per manufacturer’s instruction. The resulting cDNA was 117 extracted with phenol-chloroform and precipitated with ethanol. The cDNA was tailed 118 with dATP at the 3'-end using terminal transferase (New England Biolabs). The 50-μL 119 reaction was prepared with 5 μL of 10× NEB buffer 4, 20 μL cDNA, 0.5 μL of 10 mM 120 dATP, 0.5 μL terminal transferase, 5 μL of 2.5 mM CoCl2, and 19 μL nuclease-free 121 water. The mixture was then incubated at 37 ºC for 30 min, followed by heat inactivation 122 at 70 ºC for 10 min. The resulting cDNA, tagged with stretches of poly(dT) at the 5'-end 123 and poly(dA) at the 3'-end, was used as template for the second strand synthesis using the 124 oligo(dT) primer. The resulting product was purified (QIAquick PCR purification kit, 125 QIAGEN), end-repaired (with T4 DNA polymerase, Fermentas), and then ligated (with 126
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

7
T4 DNA ligase, Fermentas) to pUC19 cut with SmaI (Fermentas). The ligated product 127 was then transformed into XL1-Blue cells (Stratagene) and screened for insertion using 128 the blue-white screening method by plating cells on LB agar plates containing IPTG and 129 X-gal. White colonies were checked for the presence of inserts using PCR. 130 Recombinants that showed the presence of inserts were selected for plasmid purification. 131 Purified plasmids, containing the cDNA inserts, were used as templates for DNA 132 sequencing, using the strategy of primer walking. DNA sequencing was performed at the 133 University at Albany Life Sciences Molecular Biology core facility, using the BigDye 134 Terminator v3.1 (Applied Biosystems) chemistry and ABI 3730 DNA sequencher 135 (Applied Biosystems). 136 Sequence analysis. Putative open reading frames (ORFs) were identified with the 137 ORF Finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html) tool from NCBI (National 138 Center for Biotechnology Information). The protein BLAST search tool was then used to 139 assign tentative gene products. The annotated genomes are available at GenBank with 140 accession numbers of JX045649 and JX045650 for phages C-1 and Hgal1, respectively. 141 The ORF Finder did not locate phage C-1’s lysis gene, which was subsequently 142 identified by using Gene Construction Kit (Textco BioSoftwares, v. 3.5 for MacOS) 143 analyzing only sequences spanning between the 3’-end of the coat and the 5’-end of the 144 replicase genes. Both lysis protein sequences revealed expected transmembrane domains 145 (7) when analyzed using the TMHMM (http://www.cbs.dtu.dk/services/TMHMM/) tool. 146 Cloning and expression of putative lysis genes. The putative lysis genes for C-147 1 (nt. 1749 to 1946) and Hgal1 (nt. 1794 to 1991) were PCR-amplified using the 148 PfuUltra high-fidelity DNA polymerase (Stratagene) from respective cDNA with primer 149
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

8
pairs of C-1_lys_For2/C-1_lys_Rev2 and Hgal_lys_For/Hgal_lys_Rev, respectively (see 150 Supplemental Material for sequences). The start codon of the putative C-1 lysis gene was 151 changed from GUG to AUG. For C-1’s lysis gene, the resulting amplicon was double-152 digested with ClaI and EcoRV, blunt-ended with T4 DNA polymerase (Fermentas), then 153 ligated with T4 DNA ligase (Fermentas) to the expression vector pZE24 (39) that was 154 first double-digested with KpnI and EcoRV, then followed by blunt-end treatment with 155 T4 DNA polymerase. For Hgal1’s lysis gene, the amplicon was double-digested with 156 KpnI and XbaI, then ligated to the pZE24 double-digested with the same enzymes. The 157 ligated products were then transformed into XL-1 Blue cells. The identity of cloned lysis 158 genes, called pLysis_C-1 and pLysis_Hgal1, were confirmed with DNA sequencing. 159 The lysis curves were determined by induction of the cloned lysis genes. Briefly, 160 cells with the recombinant plasmids were grown in a 24-well plate containing LB broth at 161 37 ºC until an OD550 of 0.17 - 0.2 was reached. The lysis gene expression was then 162 induced by adding IPTG and arabinose to final concentrations of 1 mM and 0.5% (w/v), 163 respectively. OD550 was recorded at 2-min intervals for two hours using a 24-well plate 164 reader (Synergy HT, Biotek) maintained at 37 ºC. 165 Adsorption rate constant determination. The overnight cell cultures (~109 166 CFU/mL) were diluted 10-fold in LB broth and grown for 30 min at 30 ºC with constant 167 shaking at 250 rpm. Phages (~106 PFU/mL) were then mixed with this excess bacterial 168 cells and samples were withdrawn and filtered every 5 min for 20 min. The filtrate is 169 then plated for plaque assay to estimate the number of free phages. The adsorption rates 170 were estimated by fitting the data with the following model of ln(Pt/P0) = -αBt, where Pt 171 and P0 are phage concentrations at time 0 and time t, α, the adsorption rate constant to be 172
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

9
estimated, B the initial cell concentration. Our preliminary result showed that there is not 173 much difference in adsorption rate constant estimates when the cells are in exponential 174 growth or stationary phase, possibly due to the de-repressed state of R pili expression. 175 Also, there is not much growth in cell number during the 20 min period of the assay. 176 Therefore, the cell concentration is assumed to remain constant for the calculation of the 177 adsorption rate constant. 178 One-step growth curves. Phages at a titer of ~107 PFU/mL were mixed with 179 their respective host cells (~108 cells/mL grown in LB) and allowed to adsorb for 20 min 180 at 30 ºC. The mixture was then diluted 104-fold with pre-warmed LB broth to minimize 181 subsequent adsorption and incubated with constant shaking (250 rpm) at 30 ºC. Since 182 there is minimum adsorption after dilution, it can be assumed that the phages released 183 initially are from a single infectious cycle. Aliquots were sampled and filtered at regular 184 intervals to estimate phage numbers using plaque assay. 185 Phylogenetic analysis. A total of 14 ssRNA phage genomes: MS2 186 (NC_001417.2 (24)); fr (X15031.1 (1)); GA (X03869.1 (35)); KU1 (AF227250.1, 187 reference therein); Qβ (NC_001890.1 (4, 5)), M11 (AF052431.1 (4, 5)); SP (X07489 188 (35)); NL95 (AF059243.1 (4, 5)); AP205 (AF334111 (38)); PP7 (NC_001628.1 (46)); 189 PRR1 (NC_008294.1 (51)); φCb5 (HM066936), (37)), including C-1 and Hgal1, were 190 used for phylogenetic reconstruction. The predicted amino acid sequences of the 191 maturation/attachment protein and replicase were aligned separately using both 192 MUSCLE ((23); http://www.ebi.ac.uk/Tools/msa/muscle/) and T-Coffee (specifically, the 193 three variants: M-Coffee (59), Expresso (2), and PSI-Coffee (22); http://tcoffee.crg.cat/) 194 web servers, using the default settings. The phylogenetic reconstruction was conducted 195
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

10
using all four common algorithms: (a) the distance method (specifically, the neighbor 196 joining method (52) implemented in MEGA v. 5.05 (57), running on MacOS 10.7), (b) 197 the maximum parsimony (specifically, TNT v. 1.1 (30, 42) available at 198 http://www.phylogeny.fr/version2_cgi/one_task.cgi?task_type=tnt (21)), (c) the 199 maximum likelihood (specifically, PhyML 3.0 (31) available at http://www.atgc-200 montpellier.fr/phyml/), and (d) the Bayesian inference (specifically, MrBayes v. 3.2 (34, 201 50), running on MacOS 10.7). Most of the settings used for the analyses are defaults for 202 the programs. Specifically, for the neighbor joining method, JJT (36) was used as the 203 amino acid substitution model and one-parameter gamma distribution in rate. Five 204 hundred bootstrap replicates were used to establish branch support. For the maximum 205 parsimony, mPAM250 was used for cost and 500 bootstrap replicates were used. For the 206 maximum likelihood, the WAG substitution model (62) was used, with 100 bootstrap 207 replicates. For the Bayesian inference, a total of 100,000 generations were generated for 208 each run, with the WAG substitution model and the default burnin of 25%. FigTree v. 209 1.3.1 (49), running on MacOS 10.7, was used for tree visualization. 210 While the placement of the root for a phylogenetic tree does not change the 211 branching orders or branch lengths, it does imply ancestry of the taxa included in the 212 study. Since φCb5 is a phage infecting Caulobacter crescentus, a bacterium that belongs 213 to the α-proteobacteria class, while all the other phages used in this study infect γ-214 proteobacteria, we assumed that φCb5 is more distantly related to all the other phages, 215 therefore we rooted the tree between φCb5 and the other lineages. This decision is 216 further supported by the existence of deep homology among various RNA-dependent 217 RNA polymerases (RdRps), which included sources from RNA viruses infecting animals, 218
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

11
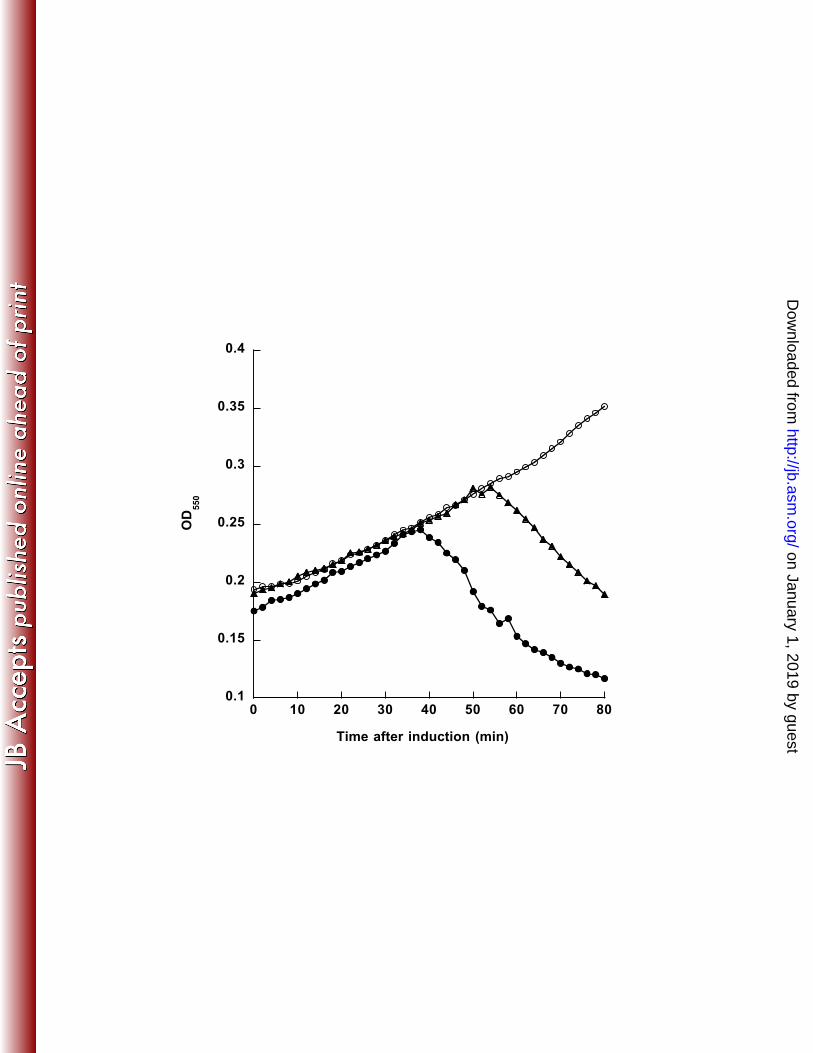
plants, fungi, and replicases from Leviviridae phages (33). For example, when including 219 the RdRp sequence from Ophiostoma mitovirus 3a (GenBank accession number 220 NC_004049), which infects the fungal host Ophiostoma novo-ulmi (33), as the outgroup 221 for phylogenetic reconstruction, the resulting trees always showed φCb5 as the first taxon 222 splitting off from the rest of the ssRNA phages, suggesting that φCb5’s more distant 223 relationship with the remaining ssRNA phages (results now shown). All trees generated 224 in this study that are used for summary consensus trees are listed in the Supplemental 225 Material. 226 227 Results 228 Genomic sequences. The genome sizes for phages C-1 and Hgal1 are, 229 respectively, 3523 and 3562 nucleotides in length. Both phages show the typical 230 Levivirus genome organization, starting with a 5' end un-translated region (UTR), 231 followed by four hypothetical open reading frames (ORFs), encoding, in order, the 232 maturation/attachment protein, coat protein, lysis protein, and replicase, and a 3' UTR 233 (Figure 1). 234 Lysis genes. A typical lysis gene in the Levivirus group spans the 3’ end of the 235 upstream coat gene and the 5’ end of the downstream replicase gene (Figure 1, (3, 6)). 236 Similar stretches of sequence are also found in C-1 and Hgal1 genomes (Figure 1), which 237 we tentatively assigned as the lysis genes. This assignment is further supported by the 238 presence of a single predicted transmembrane domain in both predicted amino acid 239 sequences. To empirically verify whether these two ORFs encode lysis proteins, we 240 cloned and expressed these putative lysis genes in the context of a medium copy-number 241
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

12
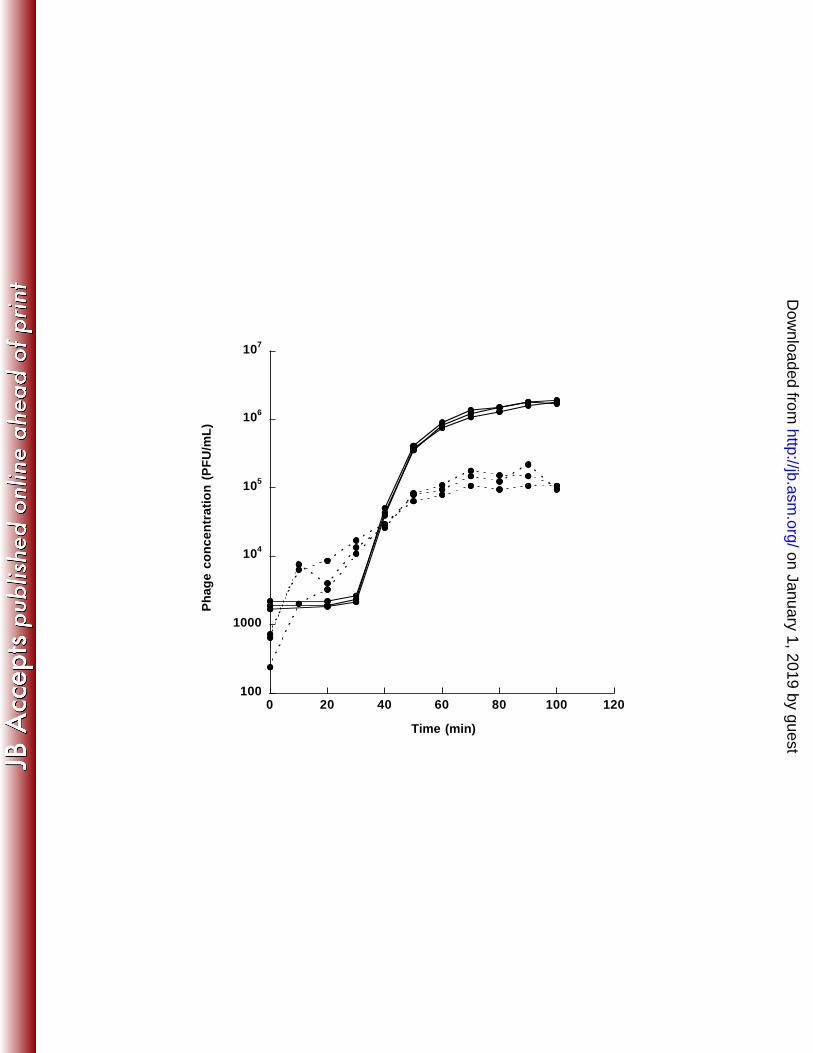
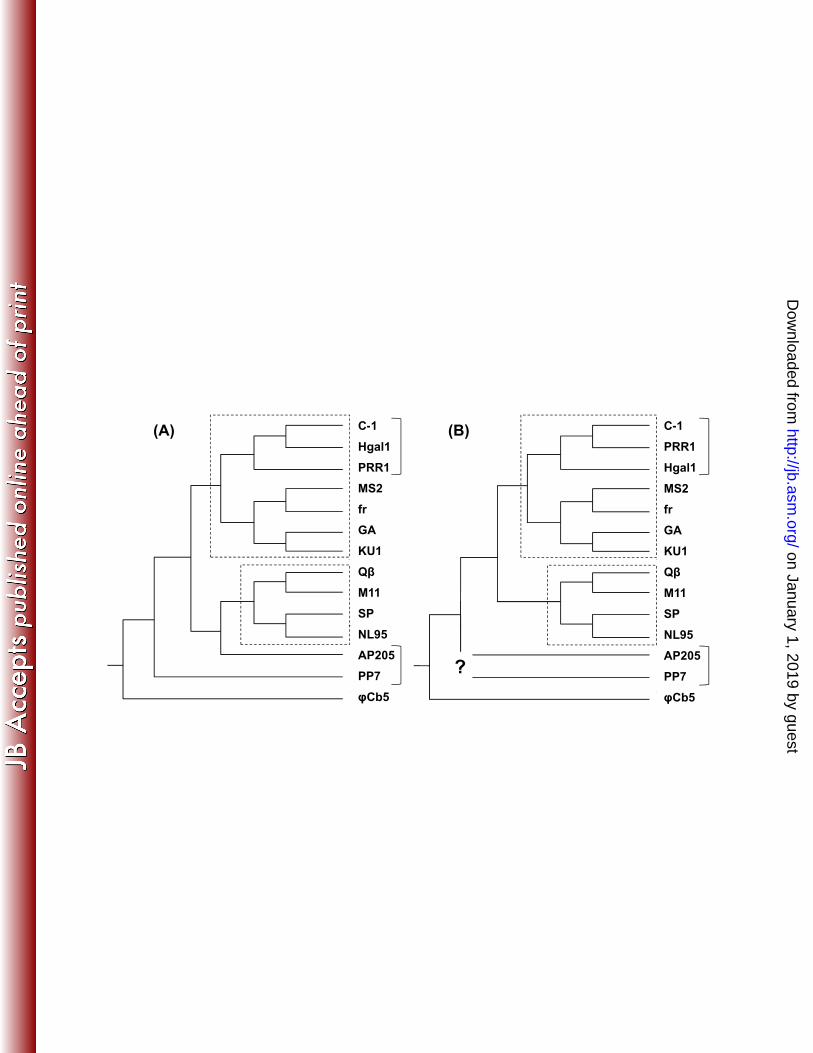
expression vector. Culture declines were visible 38 min and 54 min after induction for 242 expressed C-1 and Hgal1 ORFs, respectively (Figure 2). This result demonstrates the 243 lysis function of both lysis genes. It is interesting to note that the start codon for phage 244 C-1 is GUG, instead of the usual AUG (see Materials and Methods). 245 Growth characteristics of C-1 and Hgal1 phages. The adsorption rate 246 constants for C-1 and Hgal1 are determined to be 8.27 (± 0.997) × 10-11 and 3.44 (± 247 0.126) × 10-11 cell-1 mL-1 min-1, respectively (values in the brackets showed the standard 248 errors after correction for sample sizes (56)). One-step growth studies (Figure 3) showed 249 that C-1 phages were released almost immediately after the 20-min pre-adsorption period 250 (see Materials and Methods), while free Hgal1 phages were not observed until after about 251 40 min after infection (pre-adsorption). The average number of C-1 and Hgal1 phage 252 particles released per infected cell was estimated to be 255 and 940, respectively. 253 Phylogenetic relationship with other ssRNA phages. To investigate how C-1 254 and Hgal1 relate to other known ssRNA phages, we assembled 14 phage sequences to 255 conduct phylogenetic analyses. The gene trees for the maturation/attachment proteins 256 and replicases are reconstructed separately, using the predicted amino acid sequences, to 257 better resolve potential deep relationships. For each gene, a total of 16 trees were 258 reconstructed, each from a combination of four different sequence alignment algorithms 259 and four different phylogenetic inference algorithms (see Materials and Methods for 260 details and Supplemental Material for all inferred trees). 261 It is not unusual that different sequence alignment algorithms and phylogenetic 262 inference algorithms yield slightly different trees (in terms of branching orders). In our 263
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

13
analyses, we were able to obtain consistent branching orders with high branch supports 264 for both the maturation/attachment proteins and replicases. 265 Figure 4 summarizes our best effort in reconstructing these phylogenetic 266 relationships. In almost all cases, we were able to retrieve two well-known clades: the F-267 plasmid-dependent (FPD) Levivirus ((MS2, M12), (GA, KU1)) and Allolevivirus ((Qβ, 268 M11), (SP, NL95)). Furthermore, our analyses showed that both C-1 and Hgal1 are most 269 closely related to PRR1, an ssRNA phage first isolated from sewage in Kalamazoo, 270 Michigan using P. aeruginosa and E. coli harboring the plasmid R1822 (44). These three 271 phages thus constitute a clade that is most closely related to the FPD Levivirus (like MS2, 272 etc.), indicating that both C-1 and Hgal1, like that of PRR1, belong to the Levivirus 273 group. This pan-Levivirus clade is further clustered with the FPD Allolevivirus (like Qβ, 274 etc.) to form an even larger clade that encompasses all the known ssRNA phages that can 275 infect at least E. coli, but most likely other related bacterial species as well. 276 On the other hand, for phages that are seemingly outside this clade, such as 277 AP205 and PP7, we were unable to obtain a consistent branching order with the current 278 collection of ssRNA phages. Interestingly, majority of the maturation/attachment protein 279 trees showed that AP205 is closely related to the FPD Allolevivirus (see Supplemental 280 Material), with PP7 clustering with the others (Figure 4A). But there does not seem to be 281 a consistent pattern for the replicase trees (Figure 4B). Other evidence, such as genome 282 organization and conserved secondary RNA structures, suggest that both are Levivirus-283 like (38, 46). 284 A closer inspection of the phylogenetic relationships among C-1, Hgal1, and 285 PRR1 showed that the maturation/attachment protein tree and the replicase tree are not 286
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

14
concordant; that is, the relative genetic relatedness among these three phages depends on 287 the gene tree considered. For the maturation/attachment protein, the analysis yields the 288 relationship of C-1 clustering with Hgal1 first, then with PRR1. But when the replicase 289 tree is considered, C-1 is clustered with PRR1 first, then with Hgal1. A similar 290 reconstruction effort with the coat proteins (see Supplemental Material) showed that 291 majority of the inferred relationships was in the form of ((C-1, Hgal1), PRR1). 292 Combined together, these results suggest that the Hgal1 may have obtained its replicase 293 gene from a PRR1-like phage, possibly through a recombination event. 294 295 Discussion 296 Phages C-1, Hgal1, and PRR1 share the Levivirus characteristics. In this 297 study, we sequenced the genomes of two R-plasmid-dependent (RPD) ssRNA phages, C-298 1 and Hgal1, and determined some basic characteristics, like the adsorption rate constants 299 and one-step growth kinetics. We also empirically confirmed the predicted lysis genes by 300 demonstrating their effects on cell lysis. Phylogenetic analysis placed these two phages 301 to be closely related to phage PRR1, another RPD ssRNA phage (45). Our preliminary 302 analysis of another RPD ssRNA phage M (16) showed that phage M is even more closely 303 related to either C-1 or Hgal1, when compared to PRR1 (unpublished result). Since these 304 RPD ssRNA phages have consistently clustered with the randomly selected F-plasmid-305 dependent (FPD) ssRNA phages (i.e., MS2, M12, GA, and KU1), it places them firmly in 306 the Levivirus group. It is to be noted that the clustering among these three phages also 307 extends to the majority of the coat gene trees (see Supplemental Material), contradicting a 308 previous study (51) (their Fig. 2) in which the PRR1’s coat protein is clustered with those 309
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

15
from phages in the Allolevivirus group, like Qβ and NL95. Since only a single sequence 310 alignment algorithm (i.e., ClustalW) and phylogenetic inference algorithm (i.e., 311 UPGMA) were used for the analysis, and no statistical evaluation was conducted for 312 branch support, it is doubtful the claim that phage PRR1 shared characteristics with 313 Alloleviviruses could be further substantiated (51). 314 315 Divergence between F-plasmid-dependent Levivirus and Allolevivirus came first, 316 before independent specialization on the F plasmid. Bollack and Huelsenbeck (9) 317 conducted a thorough and rigorous phylogenetic analysis of ssRNA phages, using 318 genomic sequences available at the time. In their analysis, all phages, except for phage 319 PP7, are FPD Levivirus and Allolevivirus. Based on their study, two interesting 320 conclusions regarding the evolution of ssRNA phages were put forward: (a) the extant 321 FPD ssRNA phages are derived from a common ancestor with a genome organization 322 like that of the Levivirus, but became dependent on F plasmid for cell attachment and 323 entry, and (b) subsequent evolution of increased genome size, loss of independent lysis 324 gene, and gain of a read-through protein defines the derived FPD Allolevivirus. That is, 325 for the FPD ssRNA phages, the specialization on F plasmid came first, followed by 326 genome reorganization through the mechanism of “gene contraction” that became the 327 common ancestor for all FPD Allolevivirus. However, the clustering of the FPD 328 Levivirus with phages C-1, Hgal1, and PRR1 (Figure 4), each specializing on a different 329 R plasmid, showed that specialization on F plasmid may be a more recent evolution. 330 Most likely, the reason why the extant FPD Levivirus and Allolevivirus specialize on the 331 F plasmid is because the common ancestors for the Levivirus and Allolevivirus have 332
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

16
independently evolved to utilizing the F plasmid for cell attachment and entry. 333 Furthermore, since C-1, Hgal1, and PRR1 clustered with the FPD Levivirus as a whole, 334 rather than a specific well supported genogroup (either I or II) (25), it suggests that the 335 divergence among genogroups happened after specialization on the F plasmid. 336 337 Are there R-plasmid-dependent Allolevivirus? If the richness of FPD ssRNA phages is 338 typical among ssRNA phages, then we will expect to isolate both Levivirus and 339 Allolevivirus specific to conjugative plasmids other than the F plasmid. One possible 340 scenario is that the RPD Allolevivirus could be clustered with the FPD Allolevivirus 341 (Figure 5). If that is the case, then it will further suggest that the existence of two 342 stereotypical bauplans of genome organizations—i.e., an overall smaller genome with 343 four genes, including an independent lysis gene, for the Levivirus, and a slightly larger 344 genome with three genes, with the attachment protein functioning as the lysis protein (8), 345 and a read-through protein, for the Allolevivirus—predates the phages’ specialization on 346 these conjugative plasmids. Furthermore, because only a few amino acids are involved in 347 the interaction between the phage attachment protein and the corresponding pilin subunit 348 (26, 40), it is possible that independent evolution of the dependency on a specific 349 conjugative plasmid in the Allolevivirus clade may not follow the same evolutionary 350 pathway, thus resulting in a different branching order when compared to the Levivirus. 351 352 The genomic organization of the Allolevivirus likely evolved later. Even though there 353 is an uncertainty in the placement AP205 and PP7 in the phylogenetic tree (Figure 4), our 354 result is consistent with that of Bollack and Huelsenbeck (9) in placing PP7 as the 355
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

17
outgroup for the entire clade of Levivirus and Allolevivirus that depend on conjugative 356 plasmids for cell attachment and entry. This implies that the differentiating traits of 357 increased genome size, missing independent lysis gene, gaining of the read-through 358 protein, etc. are derived from a common ancestor whose genome organization is similar 359 to that of PP7, with the Levivirus-like genome organization. At this point, it is not clear 360 as to why there are two basic bauplans for ssRNA phage genome organization and how 361 immutable these associated genomic traits are. The presence of the lysis genes at a 362 different location (e.g., phages AP205 and φCb5) showed that the phage genomes are 363 somewhat malleable. It is apparent that more ssRNA phages specializing on different 364 conjugative plasmids and different bacterial hosts are needed for us to gain further insight 365 into their evolutionary history. 366 367 Acknowledgement 368 Research reported in this publication was supported by the National Institute of 369 General Medical Sciences of the National Institutes of Health under award number 370 R01GM072815 to INW. The content is solely the responsibility of the authors and does 371 not necessarily represent the official views of the National Institutes of Health. During 372 manuscript revision, INW was supported by the Institute of Plant and Microbial Biology, 373 Academia Sinica, and National Science Council, Taiwan. 374 375 376
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

18
Figure 1. Figure 1. Genome organization of phages C-1, Hgal1, MS2, and Qβ. Phages 377 MS2 and Qβ (GenBank accession number NC_001890.1) are used as the prototypical 378 genomes for the Levivirus and Allolevivirus, respectively. The numbers indicate the 379 beginning and end of the genomes or the hypothetical open reading frames (open bars), 380 which encode the maturation/attachment protein (mat/att), coat protein (cp), lysis protein 381 (lys), and replicase subunit (rep). The maturation/attachment protein of Qβ also 382 functions as a lysis protein. Due to incomplete termination of coat protein translation, 383 Qβ’s virion also incorporates the larger but minor read-through protein (rt). Genomic 384 sequences are obtained from GenBank (see Materials and Methods for accession 385 numbers). 386 387 Figure 2. Figure 2. Lysis curves of recombinant E. coli harboring phage lysis genes. 388 Culture densities (OD550) of E. coli strains, carrying either the cloned lysis gene from 389 phages C-1 (filled circles) or Hgal1 (filled triangles), or plasmid control (open circles), 390 were plotted against time after culture induction with IPTG. 391 392 Figure 3. One-step growth curves of phages C-1 and Hgal1. Increases in C-1 (dashed 393 lines) and Hgal1 (solid lines) phage concentrations are plotted against time after 394 infection. Three replicate experiments are shown for each phage. 395 396 Figure 4. Summary phylogenetic trees. Amino acid sequences for 14 397 maturation/attachment proteins (A) and replicases (B) were used for extensive 398 phylogenetic reconstruction. The summary trees shown are composite results of 16 trees 399
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

19
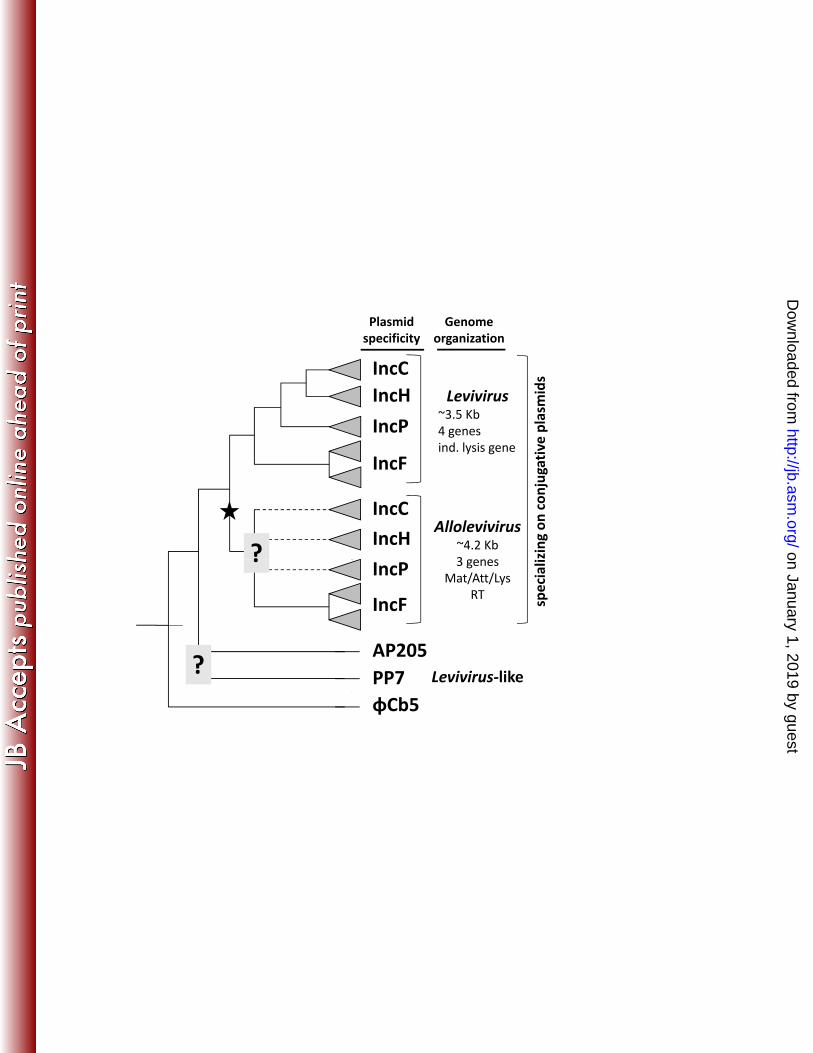
obtained by combinations of sequence alignment and phylogenetic inference algorithms. 400 The dash-lined showed the clades that are consistently clustered together. Question mark 401 denotes uncertain branching order. See Materials and Methods for details of sequence 402 alignments and tree reconstruction, and Supplemental Material for resulting trees. 403 404 Figure 5. A hypothetical phylogenetic tree of ssRNA phages. The main topology is 405 derived from Figure 4. All ssRNA phages that depend on conjugative plasmids for 406 attachment and cell entry belong to one of the two main clades, Levivirus and 407 Allolevivirus, with the later lineage thought to have evolved from a Levivirus-like 408 ancestral phage (indicated by the star sign). Plasmid-dependency evolved independently 409 within each clade, although not necessarily with the same branching order (question mark 410 indicates uncertain branching order and dashed lines hypothetical non-F-plasmid-411 dependent Allolevivirus). However, within each clade, phage strains that depend on the 412 same Inc plasmid (represented by the shaded triangles) are thought to be monophyletic. 413 Phages AP205, PP7, and φCb5 share some, but not all, genomic traits with the Levivirus. 414 The most notable difference is that the placement of the lysis gene is different for AP205 415 (before the maturation/attachment gene) and φCb5 (after the replicase gene). 416 417
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

20
References 418 419 1. Adhin, M. R., A. Avots, V. Berzin, G. P. Overbeek, and J. van Duin. 1990. 420 Complete nucleotide sequence of the group I RNA bacteriophage fr. 421 Biochimica et biophysica acta 1050:104-109. 422 2. Armougom, F., S. Moretti, O. Poirot, S. Audic, P. Dumas, B. Schaeli, V. 423
Keduas, and C. Notredame. 2006. Expresso: automatic incorporation of 424 structural information in multiple sequence alignments using 3D-Coffee. 425 Nucleic Acids Res 34:W604-608. 426 3. Atkins, J. F., J. A. Steitz, C. W. Anderson, and P. Model. 1979. Binding of 427 mammalian ribosomes to MS2 phage RNA reveals an overlapping gene 428 encoding a lysis function. Cell 18:247-256. 429 4. Beekwilder, J., R. Nieuwenhuizen, R. Poot, and J. van Duin. 1996. 430 Secondary structure model for the first three domains of Qβ RNA. Control of 431 A-protein synthesis. J Mol Biol 256:8-19. 432 5. Beekwilder, M. J., R. Nieuwenhuizen, and J. van Duin. 1995. Secondary 433 structure model for the last two domains of single-stranded RNA phage Qβ. J 434 Mol Biol 247:903-917. 435 6. Beremand, M. N., and T. Blumenthal. 1979. Overlapping genes in RNA 436 phage: a new protein implicated in lysis. Cell 18:257-266. 437 7. Bernhardt, T. G., I. N. Wang, D. K. Struck, and R. Young. 2002. Breaking 438 free: "protein antibiotics" and phage lysis. Res Microbiol 153:493-501. 439
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

21
8. Bernhardt, T. G., I. N. Wang, D. K. Struck, and R. Young. 2001. A protein 440 antibiotic in the phage Qβ virion: diversity in lysis targets. Science 292:2326-441 2329. 442 9. Bollback, J. P., and J. P. Huelsenbeck. 2001. Phylogeny, genome evolution, 443 and host specificity of single-stranded RNA bacteriophage (family 444 Leviviridae). J Mol Evol 52:117-128. 445 10. Bradley, D. E. 1966. The structure and infective process of a Pseudomonas 446 aeruginosa bacteriophage containing ribonucleic acid. J Gen Microbiol 45:83-447 96. 448 11. Bradley, D. E., J. N. Coetzee, T. Bothma, and R. W. Hedges. 1981. Phage t: a 449 group T plasmid-dependent bacteriophage. J Gen Microbiol 126:397-403. 450 12. Brion, G. M., J. S. Meschke, and M. D. Sobsey. 2002. F-specific RNA 451 coliphages: occurrence, types, and survival in natural waters. Water Res 452 36:2419-2425. 453 13. Chabbert, Y. A., M. R. Scavizzi, J. L. Witchitz, G. R. Gerbaud, and D. H. 454 Bouanchaud. 1972. Incompatibility groups and the classification of fi- 455 resistance factors. J Bacteriol 112:666-675. 456 14. Coetzee, J. N., D. E. Bradley, J. Fleming, L. du Toit, V. M. Hughes, and R. W. 457 Hedges. 1985. Phage pilHα: a phage which adsorbs to IncHI and IncHII 458 plasmid-coded pili. J Gen Microbiol 131:1115-1121. 459 15. Coetzee, J. N., D. E. Bradley, and R. W. Hedges. 1982. Phages Iα and I2-2: 460 IncI plasmid-dependent bacteriophages. J Gen Microbiol 128:2797-2804. 461
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

22
16. Coetzee, J. N., D. E. Bradley, R. W. Hedges, J. Fleming, and G. Lecatsas. 462 1983. Bacteriophage M: an incompatibility group M plasmid-specific phage. J 463 Gen Microbiol 129:2271-2276. 464 17. Coetzee, J. N., D. E. Bradley, R. W. Hedges, V. M. Hughes, M. M. McConnell, 465 L. Du Toit, and M. Tweehuysen. 1986. Bacteriophages F0lac h, SR, SF: 466 phages which adsorb to pili encoded by plasmids of the S-complex. J Gen 467 Microbiol 132:2907-2917. 468 18. Coetzee, J. N., D. E. Bradley, G. Lecatsas, L. du Toit, and R. W. Hedges. 469 1985. Bacteriophage D: an IncD group plasmid-specific phage. J Gen 470 Microbiol 131:3375-3383. 471 19. Couturier, M., F. Bex, P. L. Bergquist, and W. K. Maas. 1988. Identification 472 and classification of bacterial plasmids. Microbiol Rev 52:375-395. 473 20. Datta, N., and R. W. Hedges. 1971. Compatibility groups among fi- R factors. 474 Nature 234:222-223. 475 21. Dereeper, A., V. Guignon, G. Blanc, S. Audic, S. Buffet, F. Chevenet, J. F. 476 Dufayard, S. Guindon, V. Lefort, M. Lescot, J. M. Claverie, and O. Gascuel. 477 2008. Phylogeny.fr: robust phylogenetic analysis for the non-specialist. 478 Nucleic Acids Res 36:W465-469. 479 22. Di Tommaso, P., S. Moretti, I. Xenarios, M. Orobitg, A. Montanyola, J. M. 480 Chang, J. F. Taly, and C. Notredame. 2011. T-Coffee: a web server for the 481 multiple sequence alignment of protein and RNA sequences using structural 482 information and homology extension. Nucleic Acids Res 39:W13-17. 483
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

23
23. Edgar, R. C. 2004. MUSCLE: multiple sequence alignment with high accuracy 484 and high throughput. Nucleic Acids Res 32:1792-1797. 485 24. Fiers, W., R. Contreras, F. Duerinck, G. Haegeman, D. Iserentant, J. 486 Merregaert, W. Min Jou, F. Molemans, A. Raeymaekers, A. Van den 487 Berghe, G. Volckaert, and M. Ysebaert. 1976. Complete nucleotide 488 sequence of bacteriophage MS2 RNA: primary and secondary structure of the 489 replicase gene. Nature 260:500-507. 490 25. Friedman, S. D., F. J. Genthner, J. Gentry, M. D. Sobsey, and J. Vinje. 2009. 491 Gene mapping and phylogenetic analysis of the complete genome from 30 492 single-stranded RNA male-specific coliphages (family Leviviridae). J Virol 493 83:11233-11243. 494 26. Frost, L. S., and W. Paranchych. 1988. DNA sequence analysis of point 495 mutations in traA, the F pilin gene, reveal two domains involved in F-specific 496 bacteriophage attachment. Mol Gen Genet 213:134-139. 497 27. Furuse, K., A. Ando, S. Osawa, and I. Watanabe. 1981. Distribution of 498 ribonucleic acid coliphages in raw sewage from treatment plants in Japan. 499 Appl Environ Microbiol 41:1139-1143. 500 28. Furuse, K., T. Sakurai, A. Hirashima, M. Katsuki, A. Ando, and I. 501 Watanabe. 1978. Distribution of ribonucleic acid coliphages in south and 502 east Asia. Appl Environ Microbiol 35:995-1002. 503 29. Furuse, K., T. Sakurai, Y. Inokuchi, H. Inoko, A. Ando, and I. Watanabe. 504 1983. Distribution of RNA coliphages in Senegal, Ghana, and Madagascar. 505 Microbiol Immunol 27:347-358. 506
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

24
30. Goloboff, P. A. 1999. Analyzing large data sets in reasonable times: solutions 507 for composite optima. Cladistics 15:415-428. 508 31. Guindon, S., J. F. Dufayard, V. Lefort, M. Anisimova, W. Hordijk, and O. 509 Gascuel. 2010. New algorithms and methods to estimate maximum-510 likelihood phylogenies: assessing the performance of PhyML 3.0. Syst Biol 511 59:307-321. 512 32. Haramoto, E., M. Otagiri, H. Morita, and M. Kitajima. 2012. Genogroup 513 distribution of F-specific coliphages in wastewater and river water in the 514 Kofu basin in Japan. Lett Appl MIcrobiol 54:367-373. 515 33. Hong, Y., T. E. Cole, C. M. Brasier, and K. W. Buck. 1998. Evolutionary 516 relationships among putative RNA-dependent RNA polymerases encoded by 517 a mitochondrial virus-like RNA in the Dutch Elm Disease fungus, Ophiostoma 518 novo-ulmi, by other viruses and virus-like RNAs and by the Arabidopsis 519 mitochondrial genome. Virology 246:158-169. 520 34. Huelsenbeck, J. P., and F. Ronquist. 2001. MRBAYES: Bayesian inference of 521 phylogeny. Bioinformatics 17:754-755. 522 35. Inokuchi, Y., R. Takahashi, T. Hirose, S. Inayama, A. B. Jacobson, and A. 523 Hirashima. 1986. The complete nucleotide sequence of the group II RNA 524 coliphage GA. J Biochem 99:1169-1180. 525 36. Jones, D. T., W. R. Taylor, and J. M. Thornton. 1992. The rapid generation 526 of mutation data matrices from protein sequences. Comput Appl Biosci 527 8:275-282. 528
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

25
37. Kazaks, A., T. Voronkova, J. Rumnieks, A. Dishlers, and K. Tars. 2011. 529 Genome structure of caulobacter phage phiCb5. J Virol 85:4628-4631. 530 38. Klovins, J., G. P. Overbeek, S. H. van den Worm, H. W. Ackermann, and J. 531 van Duin. 2002. Nucleotide sequence of a ssRNA phage from Acinetobacter: 532 kinship to coliphages. J Gen Virol 83:1523-1533. 533 39. Lutz, R., and H. Bujard. 1997. Independent and tight regulation of 534 transcriptional units in Escherichia coli via the LacR/O, the TetR/O and 535 AraC/I1-I2 regulatory elements. Nucleic Acids Res 25:1203-1210. 536 40. Manchak, J., K. G. Anthony, and L. S. Frost. 2002. Mutational analysis of F-537 pilin reveals domains for pilus assembly, phage infection and DNA transfer. 538 Mol Microbiol 43:195-205. 539 41. Murphy, F. A., C. M. Fauquet, D. H. L. Bishop, S. A. Ghabrial, A. W. Javis, M. 540 A. Mayo, and M. D. Summers. 1995. Virus Taxonomy : The Classification and 541 Nomenclature of Viruses. The sixth report of the International Committee on 542 Taxonomy of Viruses. Springer-Verlag, Vienna/New York. 543 42. Nixon, K. C. 1999. The parsimony ratchet, a new method for rapid parsimony 544 analysis. Cladistics 15:407-414. 545 43. Nuttall, D., D. Maker, and E. Colleran. 1987. A method for the direct 546 isolation of IncH plasmid-dependent bacteriophages. Lett Appl MIcrobiol 547 5:37-40. 548 44. Olsen, R. H., and P. Shipley. 1973. Host range and properties of the 549 Pseudomonas aeruginosa R factor R1822. J Bacteriol 113:772-780. 550
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

26
45. Olsen, R. H., and D. D. Thomas. 1973. Characteristics and purification of 551 PRR1, an RNA phage specific for the broad host range Pseudomonas R1822 552 drug resistance plasmid. J Virol 12:1560-1567. 553 46. Olsthoorn, R. C., G. Garde, T. Dayhuff, J. F. Atkins, and J. Van Duin. 1995. 554 Nucleotide sequence of a single-stranded RNA phage from Pseudomonas 555 aeruginosa: kinship to coliphages and conservation of regulatory RNA 556 structures. Virology 206:611-625. 557 47. Osawa, S., K. Furuse, M. S. Choi, A. Ando, T. Sakurai, and I. Watanabe. 558 1981. Distribution of ribonucleic acid coliphages in Korea. Appl Environ 559 Microbiol 41:909-911. 560 48. Osawa, S., K. Furuse, and I. Watanabe. 1981. Distribution of ribonucleic 561 acid coliphages in animals. Appl Environ Microbiol 41:164-168. 562 49. Rambaut, A. 2009, posting date. FigTree v1.3.1: Tree figure drawing tool. 563 [Online.] http://tree.bio.ed.ac.uk/software/figtree/ 564 50. Ronquist, F., M. Teslenko, P. van der Mark, D. L. Ayres, A. Darling, S. 565 Hohna, B. Larget, L. Liu, M. A. Suchard, and J. P. Huelsenbeck. 2012. 566 MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice 567 across a Large Model Space. Syst Biol 61:1-4. 568 51. Ruokoranta, T. M., A. M. Grahn, J. J. Ravantti, M. M. Poranen, and D. H. 569 Bamford. 2006. Complete genome sequence of the broad host range single-570 stranded RNA phage PRR1 places it in the Levivirus genus with 571 characteristics shared with Alloleviviruses. J Virol 80:9326-9330. 572
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

27
52. Saitou, N., and M. Nei. 1987. The neighbor-joining method: a new method 573 for reconstructing phylogenetic trees. Mol Biol Evol 4:406-425. 574 53. Sakurai, T., T. Miyake, T. Shiba, and I. Watanabe. 1968. Isolation of a 575 possible fourth group of RNA phage. Jpn J Microbiol 12:544-546. 576 54. Schaper, M., J. Jofre, M. Uys, and W. O. Grabow. 2002. Distribution of 577 genotypes of F-specific RNA bacteriophages in human and non-human 578 sources of faecal pollution in South Africa and Spain. J Appl Microbiol 579 92:657-667. 580 55. Sirgel, F. A., J. N. Coetzee, R. W. Hedges, and G. Lecatsas. 1981. Phage C-1: 581 an IncC group; plasmid-specific phage. J Gen Microbiol 122:155-160. 582 56. Sokal, R. R., and F. J. Rohlf. 2011. Biometry, 4th Ed. ed. W. H. Freeman. 583 57. Tamura, K., D. Peterson, N. Peterson, G. Stecher, M. Nei, and S. Kumar. 584 2011. MEGA5: molecular evolutionary genetics analysis using maximum 585 likelihood, evolutionary distance, and maximum parsimony methods. Mol 586 Biol Evol 28:2731-2739. 587 58. van Duin, J., and N. Tsareva. 2006. Single-stranded RNA phages, p. 175-196. 588 In R. Calendar (ed.), The Bacteriophages. Oxford University Press, New York, 589 New York. 590 59. Wallace, I. M., O. O'Sullivan, D. G. Higgins, and C. Notredame. 2006. M-591 Coffee: combining multiple sequence alignment methods with T-Coffee. 592 Nucleic Acids Res 34:1692-1699. 593 60. Watanabe, I., T. Miyake, T. Sakurai, T. Shiba, and T. Ohno. 1967. Isolation 594 and grouping of RNA phages. Proc. Japan Acad. 43:204-209. 595
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

28
61. Watanabe, T., H. Nishida, C. Ogata, T. Arai, and S. Sato. 1964. Episome-596 Mediated Transfer of Drug Resistance in Enterobacteriaceae. VII. Two Types 597 of Naturally Occurring R Factors. J Bacteriol 88:716-726. 598 62. Whelan, S., and N. Goldman. 2001. A general empirical model of protein 599 evolution derived from multiple protein families using a maximum-likelihood 600 approach. Mol Biol Evol 18:691-699. 601 63. Zinder, N. D. (ed.). 1975. RNA Phages. Cold Spring Harbor Laboratory, Cold 602 Spring Harbor, New York. 603 604 605
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from
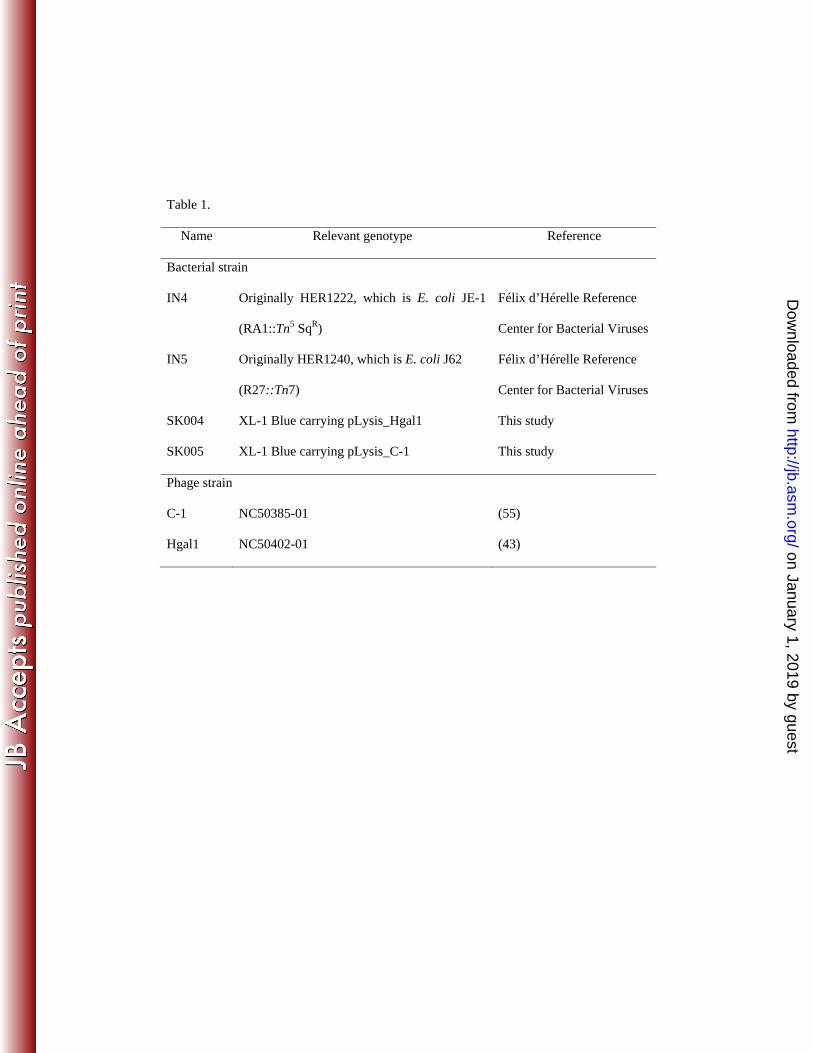
Table 1.
Name Relevant genotype Reference
Bacterial strain
IN4 Originally HER1222, which is E. coli JE-1
(RA1::Tn5 SqR)
Félix d’Hérelle Reference
Center for Bacterial Viruses
IN5 Originally HER1240, which is E. coli J62
(R27::Tn7)
Félix d’Hérelle Reference
Center for Bacterial Viruses
SK004 XL-1 Blue carrying pLysis_Hgal1 This study
SK005 XL-1 Blue carrying pLysis_C-1 This study
Phage strain
C-1 NC50385-01 (55)
Hgal1 NC50402-01 (43)
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from

C 1
1 152 1372 1379 1780 1826 3394 3523
t/ ttC-1
1749 1946
mat/att cplys
rep
Hgal1
1 174 1397 1403 1801 1880 3451 3562
mat/att cplys
rep
1794 1991
MS2
1 130 1311 1335 1727 1761 3398 3569
/MS2
1678 1905
mat/att cplys
rep
Qβ
1 56 1321 1343 1744 2351 4111 42152332
mat/att/lys cp rt rep
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from
0.4
0.35
0.3
0.25OD
550
0.2
0 1
0.15
0.10 10 20 30 40 50 60 70 80
Time after induction (min)
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from
107
106
)
105
n (P
FU/m
L)
104
once
ntra
tio
1000
Phag
e co
100
1000
1000 20 40 60 80 100 120
Time (min)
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from
C-1(A) (B)C-1 C-1
PRR1
Hgal1
MS2
(A) (B)C-1
Hgal1
PRR1
MS2
fr
GA
KU1
fr
GA
KU1
Qβ
M11
SP
Qβ
M11
SP
NL95
AP205
PP7?
NL95
AP205
PP7
φCb5φCb5
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from
Plasmidspecificity
Genome organization
IncCIncH
IncP
Levivirus~3.5 Kb4 pl
asm
ids
IncP
IncF
4 genesind. lysis gene
njug
ativ
e p
IncC
IncHzi
ng o
n co
nAllolevivirus
~4.2 Kb3?
IncP
IncF spec
iali3 genes
Mat/Att/LysRT
?
PP7AP205
? Levivirus-like
φCb5
on January 1, 2019 by guesthttp://jb.asm
.org/D
ownloaded from


















