
1-s2.0-S1096717608000578-main
12
Engineering Escherichia coli for the efficient conversion of glycerol to ethanol and co-products Syed Shams Yazdani a , Ramon Gonzalez a,b, a Department of Chemical and Biomolecular Engineering, Rice University, Houston, TX, USA b Department of Bioengineering, Rice University, Houston, TX, USA article info Article history: Received 13 January 2008 Accepted 13 August 2008 Available online 9 September 2008 Keywords: Metabolic engineering Glycerol fermentation Biofuels and biochemicals Escherichia coli Ethanol Hydrogen Formic acid abstract Given its availability, low prices, and high degree of reduction, glycerol has become an ideal feedstock for producing reduced compounds via anaerobic fermentation. We recently identified environmental conditions enabling the fermentative metabolism of glycerol in E. coli, along with the pathways and mechanisms mediating this metabolic process. In this work, we used the knowledge base created in previous studies to engineer E. coli for the efficient conversion of crude glycerol to ethanol. Our strategy capitalized on the high degree of reduction of carbon in glycerol, thus enabling the production of not only ethanol but also co-products hydrogen and formate. Two strains were created for the co- production of ethanol–hydrogen and ethanol–formate: SY03 and SY04, respectively. High ethanol yields were achieved in both strains by minimizing the synthesis of by-products succinate and acetate through mutations that inactivated fumarate reductase (DfrdA) and phosphate acetyltransferase (Dpta), respectively. Strain SY04, which produced ethanol–formate, also contained a mutation that inactivated formate–hydrogen lyase (DfdhF), thus preventing the conversion of formate to CO 2 and H 2 . High rates of glycerol utilization and product synthesis were achieved by simultaneous overexpression of glycerol dehydrogenase (gldA) and dihydroxyacetone kinase (dhaKLM), which are the enzymes responsible for the conversion of glycerol to glycolytic intermediate dihydroxyacetone phosphate. The resulting strains, SY03 (pZSKLMgldA) and SY04 (pZSKLMgldA), produced ethanol–hydrogen and ethanol–formate from unrefined glycerol at yields exceeding 95% of the theoretical maximum and specific rates in the order of 15–30 mmol/gcell/h. These yields and productivities are superior to those reported for the conversion of glycerol to ethanol–H 2 or ethanol–formate by other organisms and equivalent to those achieved in the production of ethanol from sugars using E. coli. & 2008 Elsevier Inc. All rights reserved. 1. Introduction Glycerol has become an inexpensive and abundant carbon source due to its generation as inevitable by-product of biodiesel fuel production. With every 100 lbs of biodiesel produced by the transesterification of vegetable oils or animal fats, 10lbs of crude glycerol are generated. The tremendous growth of the biodiesel industry has created a glycerol surplus that resulted in a dramatic decrease in crude glycerol prices (Yazdani and Gonzalez, 2007 and references cited therein). This decrease in prices poses a problem for the glycerol-producing and -refining industries, and the economic viability of the biodiesel industry itself has been greatly affected (McCoy, 2006, 2005). The conversion of low-priced glycerol streams to higher value products has been proposed as a path to economic viability for the biofuels industry (Yazdani and Gonzalez, 2007). Such technologies could be readily integrated into existing biodiesel facilities, thus establishing true biorefi- neries and revolutionizing the biodiesel industry by dramatically improving its economics. While availability and low prices make glycerol an attractive carbon source for fermentation processes, there is yet another advantage in using this compound: fuels and reduced chemicals can be produced from glycerol at yields higher than those obtained from common sugars (Yazdani and Gonzalez, 2007). The latter is possible because the degree of reduction per carbon, k (Nielsen et al., 2003), of glycerol is significantly higher (C 3 H 8 O 3 : k ¼ 4.67) than that of sugars such as glucose (C 6 H 12 O 6 : k ¼ 4) or xylose (C 5 H 10 O 5 : k ¼ 4). To fully realize the aforemen- tioned advantages, the use of anaerobic fermentation is highly desirable. While many microorganisms are able to metabolize glycerol in the presence of external electron acceptors (respiratory metabolism), few are able to do so fermentatively (i.e., in the ARTICLE IN PRESS Contents lists available at ScienceDirect journal homepage: www.elsevier.com/locate/ymben Metabolic Engineering 1096-7176/$ - see front matter & 2008 Elsevier Inc. All rights reserved. doi:10.1016/j.ymben.2008.08.005 Corresponding author at: Department of Chemical and Biomolecular Engineer- ing, Rice University, 6100 Main Street, MS-362, Houston, TX 77005, USA. Fax: +1713 348 5478. E-mail address: [email protected] (R. Gonzalez). Metabolic Engineering 10 (2008) 340–351
-
Upload
vaibhav-nikhar -
Category
Documents
-
view
220 -
download
0
description
123
Transcript of 1-s2.0-S1096717608000578-main

ARTICLE IN PRESS
Metabolic Engineering 10 (2008) 340–351
Contents lists available at ScienceDirect
Metabolic Engineering
1096-71
doi:10.1
� Corr
ing, Ric
Fax: +1
E-m
journal homepage: www.elsevier.com/locate/ymben
Engineering Escherichia coli for the efficient conversion of glycerol to ethanoland co-products
Syed Shams Yazdani a, Ramon Gonzalez a,b,�
a Department of Chemical and Biomolecular Engineering, Rice University, Houston, TX, USAb Department of Bioengineering, Rice University, Houston, TX, USA
a r t i c l e i n f o
Article history:
Received 13 January 2008
Accepted 13 August 2008Available online 9 September 2008
Keywords:
Metabolic engineering
Glycerol fermentation
Biofuels and biochemicals
Escherichia coli
Ethanol
Hydrogen
Formic acid
76/$ - see front matter & 2008 Elsevier Inc. A
016/j.ymben.2008.08.005
esponding author at: Department of Chemica
e University, 6100 Main Street, MS-362, Hous
713 348 5478.
ail address: [email protected] (R. Gon
a b s t r a c t
Given its availability, low prices, and high degree of reduction, glycerol has become an ideal feedstock
for producing reduced compounds via anaerobic fermentation. We recently identified environmental
conditions enabling the fermentative metabolism of glycerol in E. coli, along with the pathways and
mechanisms mediating this metabolic process. In this work, we used the knowledge base created in
previous studies to engineer E. coli for the efficient conversion of crude glycerol to ethanol. Our strategy
capitalized on the high degree of reduction of carbon in glycerol, thus enabling the production of not
only ethanol but also co-products hydrogen and formate. Two strains were created for the co-
production of ethanol–hydrogen and ethanol–formate: SY03 and SY04, respectively. High ethanol yields
were achieved in both strains by minimizing the synthesis of by-products succinate and acetate through
mutations that inactivated fumarate reductase (DfrdA) and phosphate acetyltransferase (Dpta),
respectively. Strain SY04, which produced ethanol–formate, also contained a mutation that inactivated
formate–hydrogen lyase (DfdhF), thus preventing the conversion of formate to CO2 and H2. High rates of
glycerol utilization and product synthesis were achieved by simultaneous overexpression of glycerol
dehydrogenase (gldA) and dihydroxyacetone kinase (dhaKLM), which are the enzymes responsible for
the conversion of glycerol to glycolytic intermediate dihydroxyacetone phosphate. The resulting strains,
SY03 (pZSKLMgldA) and SY04 (pZSKLMgldA), produced ethanol–hydrogen and ethanol–formate from
unrefined glycerol at yields exceeding 95% of the theoretical maximum and specific rates in the order of
15–30 mmol/gcell/h. These yields and productivities are superior to those reported for the conversion of
glycerol to ethanol–H2 or ethanol–formate by other organisms and equivalent to those achieved in the
production of ethanol from sugars using E. coli.
& 2008 Elsevier Inc. All rights reserved.
1. Introduction
Glycerol has become an inexpensive and abundant carbonsource due to its generation as inevitable by-product of biodieselfuel production. With every 100 lbs of biodiesel produced by thetransesterification of vegetable oils or animal fats, 10 lbs of crudeglycerol are generated. The tremendous growth of the biodieselindustry has created a glycerol surplus that resulted in a dramaticdecrease in crude glycerol prices (Yazdani and Gonzalez, 2007 andreferences cited therein). This decrease in prices poses a problemfor the glycerol-producing and -refining industries, and theeconomic viability of the biodiesel industry itself has been greatlyaffected (McCoy, 2006, 2005). The conversion of low-priced
ll rights reserved.
l and Biomolecular Engineer-
ton, TX 77005, USA.
zalez).
glycerol streams to higher value products has been proposed asa path to economic viability for the biofuels industry (Yazdani andGonzalez, 2007). Such technologies could be readily integratedinto existing biodiesel facilities, thus establishing true biorefi-neries and revolutionizing the biodiesel industry by dramaticallyimproving its economics. While availability and low prices makeglycerol an attractive carbon source for fermentation processes,there is yet another advantage in using this compound: fuels andreduced chemicals can be produced from glycerol at yields higherthan those obtained from common sugars (Yazdani and Gonzalez,2007). The latter is possible because the degree of reduction percarbon, k (Nielsen et al., 2003), of glycerol is significantly higher(C3H8O3: k ¼ 4.67) than that of sugars such as glucose (C6H12O6:k ¼ 4) or xylose (C5H10O5: k ¼ 4). To fully realize the aforemen-tioned advantages, the use of anaerobic fermentation is highlydesirable.
While many microorganisms are able to metabolize glycerolin the presence of external electron acceptors (respiratorymetabolism), few are able to do so fermentatively (i.e., in the
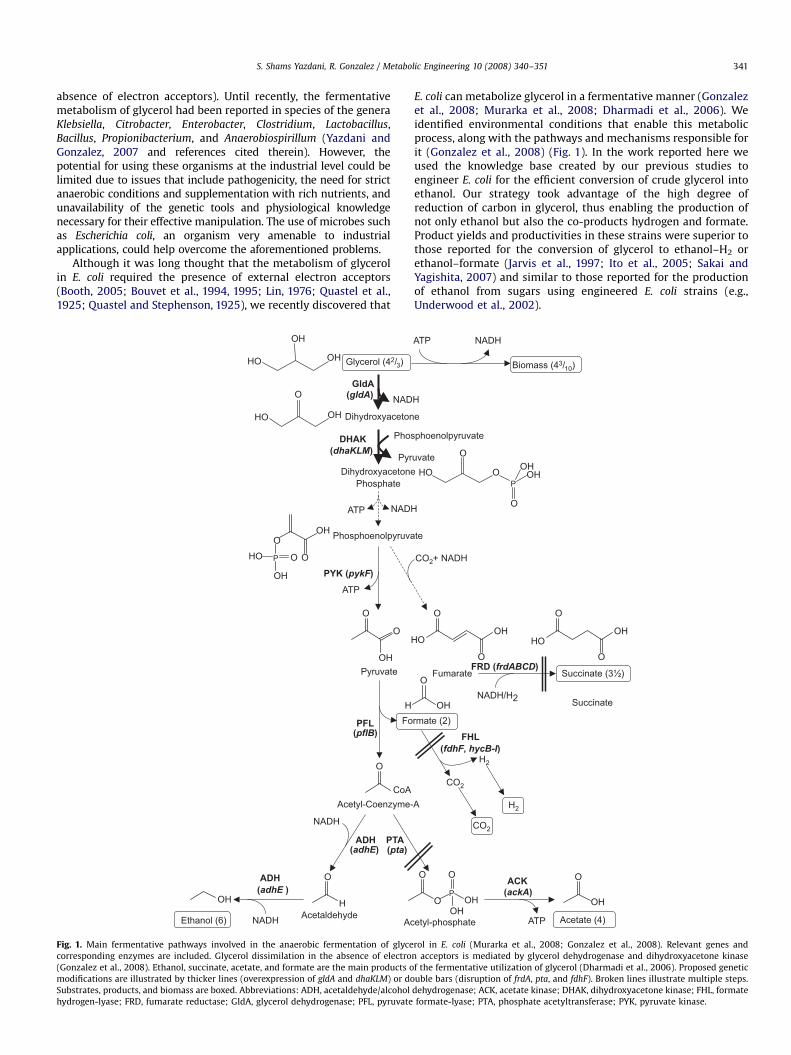
ARTICLE IN PRESS
S. Shams Yazdani, R. Gonzalez / Metabolic Engineering 10 (2008) 340–351 341
absence of electron acceptors). Until recently, the fermentativemetabolism of glycerol had been reported in species of the generaKlebsiella, Citrobacter, Enterobacter, Clostridium, Lactobacillus,Bacillus, Propionibacterium, and Anaerobiospirillum (Yazdani andGonzalez, 2007 and references cited therein). However, thepotential for using these organisms at the industrial level could belimited due to issues that include pathogenicity, the need for strictanaerobic conditions and supplementation with rich nutrients, andunavailability of the genetic tools and physiological knowledgenecessary for their effective manipulation. The use of microbes suchas Escherichia coli, an organism very amenable to industrialapplications, could help overcome the aforementioned problems.
Although it was long thought that the metabolism of glycerolin E. coli required the presence of external electron acceptors(Booth, 2005; Bouvet et al., 1994, 1995; Lin, 1976; Quastel et al.,1925; Quastel and Stephenson, 1925), we recently discovered that
PTA
FoPFL
ADH
Ethanol (6)
NADH
NADH
ATP
O
OH
OH
OH
OP
OH
O
O
HO
H
OH H
O
H
O
CoA
Phosphate
O
HO OH
NADHATP
OH
OHHO Glycerol (42/3)
GldA(gldA) NAD
Dihydroxyaceton
Phos
PyrDihydroxyacetone
Phosphoenolpyruva
PYK (pykF)
Pyruvate
(pflB)
Acetyl-Coenzyme-
(adhE) (pta)
AcetaldehydeAc
ADH(adhE )
DHAK(dhaKLM)
Fig. 1. Main fermentative pathways involved in the anaerobic fermentation of glyce
corresponding enzymes are included. Glycerol dissimilation in the absence of electro
(Gonzalez et al., 2008). Ethanol, succinate, acetate, and formate are the main products o
modifications are illustrated by thicker lines (overexpression of gldA and dhaKLM) or do
Substrates, products, and biomass are boxed. Abbreviations: ADH, acetaldehyde/alcohol
hydrogen-lyase; FRD, fumarate reductase; GldA, glycerol dehydrogenase; PFL, pyruvate
E. coli can metabolize glycerol in a fermentative manner (Gonzalezet al., 2008; Murarka et al., 2008; Dharmadi et al., 2006). Weidentified environmental conditions that enable this metabolicprocess, along with the pathways and mechanisms responsible forit (Gonzalez et al., 2008) (Fig. 1). In the work reported here weused the knowledge base created by our previous studies toengineer E. coli for the efficient conversion of crude glycerol intoethanol. Our strategy took advantage of the high degree ofreduction of carbon in glycerol, thus enabling the production ofnot only ethanol but also the co-products hydrogen and formate.Product yields and productivities in these strains were superior tothose reported for the conversion of glycerol to ethanol–H2 orethanol–formate (Jarvis et al., 1997; Ito et al., 2005; Sakai andYagishita, 2007) and similar to those reported for the productionof ethanol from sugars using engineered E. coli strains (e.g.,Underwood et al., 2002).
ACK
CO2
rmate (2)
H2
FHL
ATP
H2
CO2
O
O
OH
O
HO
O
O
OH
OH
O
OH
O
OP
OH
O
OH
O
O
HO OP
OH
O
OH
Biomass (43/10)
ATP NADH
H
e
phoenolpyruvate
uvate
te
CO2+ NADH
(fdhF, hycB-I)
A
etyl-phosphate
(ackA)
Acetate (4)
Succinate
Succinate (3½)
NADH/H2
FumarateFRD (frdABCD)
rol in E. coli (Murarka et al., 2008; Gonzalez et al., 2008). Relevant genes and
n acceptors is mediated by glycerol dehydrogenase and dihydroxyacetone kinase
f the fermentative utilization of glycerol (Dharmadi et al., 2006). Proposed genetic
uble bars (disruption of frdA, pta, and fdhF). Broken lines illustrate multiple steps.
dehydrogenase; ACK, acetate kinase; DHAK, dihydroxyacetone kinase; FHL, formate
formate-lyase; PTA, phosphate acetyltransferase; PYK, pyruvate kinase.

ARTICLE IN PRESS
S. Shams Yazdani, R. Gonzalez / Metabolic Engineering 10 (2008) 340–351342
2. Materials and methods
2.1. Strains, plasmids, and genetic methods
Wild-type K12 E. coli strain MG1655 was obtained from theUniversity of Wisconsin E. coli Genome Project (www.genome.wisc.edu) (Kang et al., 2004) and used as the host to implementthe proposed metabolic engineering strategies. All resultingstrains, along with primers and plasmids used in their construc-tion, are listed in Table 1.
Gene knockouts were introduced in MG1655 and its deriva-tives by P1 phage transduction as described elsewhere (Miller,1972). Single gene knock out mutants from the NationalBioResource Project (NIG, Japan) (Baba et al., 2006) were usedas donors of specific mutations. Various dilutions of P1 phagewere mixed with the donor cells at exponential growth phase andtryptone top agar (1% tryptone, 0.5% NaCl, 0.65% Agar, 5 mMCaCl2) and overlaid on Luria-Bertani broth (LB) agar platescontaining 5 mM CaCl2. Plates showing many isolated plaqueswere used to collect phage lysates by adding 2 ml P1 adsorptionmedia (LB+5 mM CaCl2) and incubating overnight at 4 1C. Thephage lysate was further shaken with chloroform to removebacterial cell background and used to reinfect the donor cells toenrich for the mutation. The enriched phages were mixed with therecipient strain at a multiplicity of infection (MOI) of 0.5 andincubated in 1XA media (10.5 g/L K2HPO4, 4.5 g/L KH2PO4, 1 g/L(NH4)2SO4, and 0.5 g/L sodium citrate.2H2O) for 2 h. Cells werecentrifuged and plated on LB plates supplemented with appro-priate antibiotics. The plates were incubated overnight at roomtemperature and then shifted to 30 1C and further grown for 1–2days until colonies appeared. Colonies were purified on fresh LBplates, incubated at 42 1C to cure the remaining lysogens, andfurther grown at 37 1C. To eliminate the kanamycin resistancemarker, the mutants were transformed with pCP20 (Datsenko and
Table 1Strains, plasmids, and primers used in this study
Strain/plasmid/primer Description/Genotype/Sequence
Strains
MG1655 F- l- ilvG- rfb-50 rph-1
YD01 MG1655, DfdhF::FRT-tet-FRT; deletion mutant for fdhF gene
SY01 MG1655, DfrdA::FRT-tet-FRT; deletion mutant for frdA gene
SY02 MG1655, DfdhF::FRT DfrdA::FRT-tet-FRT; deletion mutant fo
SY03 MG1655, DfrdA::FRT Dpta::FRT-tet-FRT; deletion mutant for
SY04 MG1655, DfdhF::FRT DfrdA::FRT Dpta::FRT-tet-FRT; deletion
Plasmids
pZSKLM E. coli dhaKLM gene under control of PLtetO-1 (tetR, oriR SC1
pZSKLcf C. freundii dhaKL gene under control of PLtetO-1 (tetR, oriR S
pZSGldA E. coli gldA gene under control of PLtetO-1 (tetR, oriR SC101*
pZSKLMGldA E. coli dhaKLM and gldA genes under control of PLtetO-1 (tet
pZSblank Blank plasmid created by removing C. freundii dhaKL fragm
oriR SC101*, cat)
Primersa
v-fdhF cgtaatatcagggaatgaccc
gggcaaagaatgtcaaaaacaa
v-frdA accctgaagtacgtggctg
gcaccacctcaattttcagg
v-pta gctgttttgtaacccgcc
gcagcgcaaagctgcgg
c1-gldA agtcacggtaccatggaccgcattattcaatcac
gatcgtctgcagttattcccactcttgcaggaaac
c2-glda gacactgcagaggagcaattatggaccgca
caagctacgcgtttattcccactcttgcagga
a ‘‘v’’ indicates the primer sequences (5’ to 3’) that were used for verification purpo
the primers that were used for cloning purposes, ‘‘c1’’ to clone gldA alone (pZSGldA) and
the reverse sequence in each case. Genes or operons deleted or cloned are apparent fr
Wanner, 2000), a temperature sensitive plasmid expressingflippase (FLP) recombinase. FLP expressed from this plasmidremoved the Km region from the FRT::Km::FRT site, leaving oneFRT site behind. pCP20 was then removed by growing the cells at42 1C. All mutations created in the host cells were confirmed bypolymerase chain reaction using the ‘‘verification’’ primers listedin Table 1. The disruption of multiple genes in a common host wasachieved by sequentially implementing the procedure describedabove. This sequential procedure, in turn, could lead to chromo-somal rearrangements resulting from FLP-promoted recombina-tion events between FRT sites at different loci (Datsenko andWanner, 2000). To verify that these events did not occur in strainswith multiple gene disruptions, each strain was rechecked inall disruption sites by PCR every time a new mutation wasintroduced.
The construction of plasmids used in this study is illustrated inFig. 2. The control plasmid pZSblank was constructed by digestingpZSKLcf (Bachler et al., 2005) with KpnI and BamHI restrictionendonucleases, blunting overhangs with T4 DNA polymerase, andligating the vector backbone with T4 DNA ligase. The expressionvector pZSgldA was constructed as follows. The coding region ofthe gldA gene was PCR amplified using genomic DNA of E. coli
MG1655 as template and ‘‘c1-gldA’’ primers (Table 1). Therestriction enzyme sites KpnI and PstI were introduced throughthe forward and reverse primers, respectively, to facilitate cloningof the PCR product in the expression vector pZSKLM (Bachler et al.,2005). The amplified product (1.1 kB) was digested with KpnI andPstI and used for ligation at the corresponding sites of the pZSKLMplasmid. The expression vector pZSKLMGldA was constructed byamplifying the gldA gene along with its ribosome binding sitefrom the genomic DNA of E. coli MG1655 using ‘‘c2-gldA’’ primers(Table 1). The amplified product was digested with PstI and MluIand cloned at the corresponding sites of pZSKLM, downstream ofthe dhaKLM gene. The PCR was performed using Pfu turbo DNA
Source
Kang et al., 2004
in MG1655 strain This study
in MG1655 strain This study
r frdA gene in YD01 strain This study
pta gene in SY01 strain This study
mutant for pta gene in SY02 strain This study
01*, cat) Bachler et al., 2005
C101*, cat) Bachler et al., 2005
, cat) This study
R, oriR SC101*, cat) This study
ent from pZSKLcf and self-ligating the plasmid (tetR, This study
This study
This study
This study
This study
This study
ses during the creation of disruption mutants by phage transduction. ‘‘c’’ indicates
‘‘c2’’ to clone gldA along with dhaKLM (pZSKLMGldA). The forward sequence follows
om primer names.

ARTICLE IN PRESS
Fig. 2. Diagram illustrating the construction of plasmids used in this study. Plasmids pZSKLcf and pZSKLM have been described elsewhere (Bachler et al., 2005).
S. Shams Yazdani, R. Gonzalez / Metabolic Engineering 10 (2008) 340–351 343
polymerase (Stratagene, CA, USA) under standard conditionsdescribed by the supplier. The ligated products described abovewere used to transform E. coli DH5aT1 (Invitrogen, CA, USA).Positive clones were screened by plasmid isolation and restrictiondigestion.
Standard recombinant DNA procedures were used for genecloning, plasmid isolation, and electroporation. Manufacturerprotocols and standard methods (Miller, 1972; Sambrook et al.,1989) were followed for DNA purification (Qiagen, CA, USA),restriction endonuclease digestion (New England Biolabs, MA,USA), and DNA amplification (Stratagene, CA, USA and Invitrogen,CA, USA). The strains were kept in 32.5% glycerol stocks at �80 1C.Plates were prepared using LB medium containing 1.5% agar, andappropriate antibiotics were included at the following concentra-tions: 34mg/ml chloramphenicol and 50mg/ml kanamycin.
2.2. Culture medium and cultivation conditions
The minimal medium (MM) designed by Neidhardt et al.(1974) supplemented with 2 g/L tryptone (Difco, USA), 5mMsodium selenite, 1.32 mM Na2HPO4 in place of K2HPO4, andvarying concentrations of glycerol was used unless otherwisespecified. Morpholino-propanesulfonic (MOPS) acid was onlyincluded in the inoculum preparation phase or in experimentsconducted in tubes (see below). Antibiotics and inducer (an-hydrotetracycline) were included when appropriate. Chemicalswere obtained from Fisher Scientific (Pittsburgh, PA) and Sigma-
Aldrich Co. (St Louis, MO). Experiments using crude glycerolobtained in the production of biodiesel were conducted in themedium described above. The crude glycerol, kindly provided byFutureFuel Chemical Company (Batesville, AR), had the followingcomposition (w/v): 84% glycerol, 5% salts, and 0.02% methanol.
Fermentations were conducted in a SixFors multi-fermentationsystem (Infors HT, Bottmingen, Switzerland) with six 500-mlworking volume vessels and independent control of temperature(37 1C), pH, stirrer speed (200 rpm), and dissolved oxygen (notcontrolled) (Dharmadi et al., 2006). The system is fully equippedand computer controlled using manufacturer IRIS NT software.Each vessel was fitted with a condenser operated with a 2 1Ccooling methanol–water supply to minimize evaporation. Anae-robic conditions were maintained by sparging the medium withultrahigh purity argon (Matheson, Tri-Gas, Houston, TX) at0.01 L/min. In some experiments (specified in each case), air wasflushed through the headspace at 0.01 L/min to create a micro-aerobic environment. Operation under these conditions resultedin decreasing dissolved oxygen concentrations that reachedundetectable levels after 1 h of cultivation (i.e., undetectabledissolved oxygen concentrations during almost the entire courseof the fermentation). Experiments in tubes were conducted using17-ml Hungate tubes (Bellco Glass, Inc., Vineland, NJ), which weremodified by piercing the septa with two luer lock needles, one foroxygen-free argon sparging (20 G�8 in, Hamilton Company-USA,Reno, NV) and one for gas efflux (20 G�2 in, Hamilton Company-USA, Reno, NV). The working volume of these modified Hungatetubes was 10 ml.

ARTICLE IN PRESS
S. Shams Yazdani, R. Gonzalez / Metabolic Engineering 10 (2008) 340–351344
Prior to use, the cultures (stored as glycerol stocks at �80 1C)were streaked onto LB plates and incubated overnight at 37 1C. Asingle colony was used to inoculate 17.5-ml Hungate tubescompletely filled with the medium (MM supplemented with10 g/L tryptone, 5 g/L yeast extract, and 10 g/L glycerol). The tubeswere incubated at 37 1C until an OD550 of �0.4 was reached. Anappropriate volume of this actively growing pre-culture wascentrifuged, and the pellet was washed and used to inoculate350 ml of medium in each fermenter, with a target initial opticaldensity of 0.05 at 550 nm.
2.3. Analytical methods
Optical density was measured at 550 nm and used as anestimate of cell mass (1 OD ¼ 0.34 g dry weight/L). Aftercentrifugation, the supernatant was stored at �20 1C for HPLC(high performance liquid chromatography) analysis. Glycerol,organic acids, ethanol, and hydrogen were quantified as pre-viously described (Dharmadi et al., 2006; Dharmadi and Gonzalez,2005). The transfer of oxygen in microaerobic cultures wascharacterized by estimating the volumetric oxygen transfer rate(NO2
in mg O2/L/h) as NO2¼ kLaðC � �CLÞ. kL is the oxygen
transport coefficient (cm/h), a is the gas–liquid interfacial area(cm2/cm3), kLa is the volumetric oxygen transfer coefficient (h�1),C* is the saturated dissolved oxygen concentration (mg/L), and CL
is the actual dissolved oxygen concentration in the liquid (mg/L)(Shuler and Kargi, 2002). The volumetric oxygen transfercoefficient (kLa) was estimated by the static gassing out method(Shuler and Kargi, 2002). The oxygen transfer rate, which, underoxygen limiting conditions, also represents the volumetric oxygenuptake rate, was then calculated as described above. In thesecalculations, C* was assumed to be 8 mg/L.
2.4. Enzyme activities
The activity of glycerol dehydrogenase in the oxidation ofglycerol was measured as described by Truniger and Boos (1994).Cells from anaerobic cultures (OD550 of �0.7) were harvested bycentrifugation (2 min, 10,000� g), washed twice with 9 g/L NaCl,and stored as cell pellets at �20 1C. For glycerol dehydrogenaseassays, the cells were resuspended in 0.2 ml of the appropriatebuffer (depending on the pH of the assay: see below) andpermeabilized by vortex mixing with chloroform (Osman et al.,1987; Tao et al., 2001). Activity towards glycerol was assayed bymeasuring the change in absorbance at 340 nm and 25 1C in a 1 mlreaction mixture containing 2 mM MgCl2, 500mM NAD+, 100 mMglycerol, 30ml crude cell extract, and 100 mM of the appropriatebuffer according to the pH of the assay (Truniger and Boos, 1994).To study the effect of pH on glycerol dehydrogenase activity, thefollowing buffers were used (pH in parenthesis): sodium phos-phate (pH 6–8), potassium carbonate (pH 9.5), and sodium citrate(pH 5–6). Dihydroxyacetone kinase activity was assayed using themethod reported by Kornberg and Reeves (1972) with minormodifications. Cells from anaerobic cultures (OD550 of �0.7) wereharvested by centrifugation (2 min, 10,000� g), washed withdecryptification buffer (0.1 M sodium–potassium phosphate, pH7.2, and 5 mM MgCl2) and stored as cell pellets at �20 1C. Forenzyme assays, cells were resuspended in decryptification bufferto obtain �1 mg dry cell weight/ml. A portion of the ice-cold cellsuspension was placed in a test tube and vigorously mixed for1 min; 0.2 volume of toluene–ethanol (1:9, v/v) was added. Theassay was conducted in a 1 ml reaction mixture containing 1 mMphosphoenolpyruvate trisodium, 0.1 mM NADH, 2 U lactate dehy-drogenase, and the toluene–ethanol treated cells (�50mg dryweight). The assay mixture was incubated at 30 1C for 15 min.
Dihydroxyacetone was added to a concentration of 1 mM and thedecrease in absorbance was followed at 340 nm. All spectro-photometric measurements were conducted in a BioMate 5spectrophotometer (Thermo Scientific, MA, USA). The linearity ofreactions (protein concentration and time) was established for allassays and the nonenzymatic rates were subtracted from theobserved initial reaction rates. Enzyme activities are reported asmmol of substrate/min/mg of cell protein and represent averagesfor at least three cell preparations.
2.5. Calculation of fermentation parameters
Data for cell growth, glycerol consumption, and productsynthesis were used to calculate volumetric rates/productivities(mmol/L/h) for each 12-h interval. Specific rates (mmol/gcell/h)were calculated by dividing the volumetric rates by the time–average concentration of cells over the same period of time.Growth and product yields (mmol/mmol of glycerol) werecalculated as the amount of cell mass or product synthesizedper amount of glycerol consumed once the cultures reached thestationary phase. In the above calculations, we used an averagemolecular weight of 24.7, which corresponds to an average cell ofa molecular formula CH1.9O0.5N0.2 (Nielsen et al., 2003).
3. Results and discussion
3.1. Metabolic engineering strategy to efficiently convert glycerol into
ethanol and co-products hydrogen and formate
Improvement of yield and rates are two of the most anticipatedoutcomes of a metabolic engineering effort. In this study, wefocused on the improvement of these two parameters during theconversion of glycerol to ethanol and co-products hydrogen andformate. In the following we briefly describe our engineeringstrategies.
Yield of ethanol and co-products: the fermentation of glycerol byE. coli results in a product mixture containing predominantlyethanol but also acetate, succinate, and minor amounts of formate(Dharmadi et al., 2006). The synthesis of ethanol is ‘‘preferred’’over other fermentation products because it represents a redox-balanced pathway that produces ATP via substrate-level phos-phorylation: one molecule of ATP is generated per each moleculeof glycerol converted to ethanol (Fig. 1). Since ethanol wasconsidered the target product in this study, succinate and acetatebecome competing by-products that would eventually decreasethe ethanol yield by diverting carbon to their formation (Fig. 1).Our strategy to minimize the generation of these by-productsinvolves blocking the metabolic pathways responsible for theirsynthesis (Fig. 1). Unlike succinate and acetate, formate representsa potential co-product that could enhance the value of thefermentation process. Note that glycerol could be converted intoethanol plus either hydrogen or formate (Fig. 1). The significantdifferences in the properties of ethanol and formate (boiling point,ionization constant, etc.) would facilitate their separation. Analternative approach, also explored in this work, is the completeconversion of formate to carbon dioxide and hydrogen, a processthat would yield another important biofuel—H2. Our study usedthese two approaches, namely: (i) co-production of ethanol andH2, and (ii) co-production of ethanol and formate (Fig. 1). Themaximum theoretical yield in both cases is 1 mol of ethanol plus
1 mol of either formate or hydrogen per each mol of glycerolutilized.
Rates of glycerol utilization and synthesis of ethanol and co-
products: we previously reported that a two-enzyme pathway is
ARTICLE IN PRESS
0
0.1
0.2
0.3
0.4
0.5
0.6
Cells Succinate Acetate Ethanol Formate
Prod
uct y
ield
(g/g
gly
cero
l)
2
4
6
8
10
12
Gly
cero
l util
izat
ion
(g/L
)
0.2
0.4
0.6
0.8
Cel
l gro
wth
(OD
550)
S. Shams Yazdani, R. Gonzalez / Metabolic Engineering 10 (2008) 340–351 345
responsible for the conversion of glycerol to glycolytic intermedi-ates (Fig. 1) (Gonzalez et al., 2008). These two enzymes convertglycerol to dihydroxyacteone (DHA) (enzyme glycerol dehydro-genase, GldA, encoded by the gldA gene) and DHA to dihydrox-yacetone phosphate (DHAP) (enzyme DHA kinase, DHAK, encodedby the dhaKLM operon). This trunk pathway, however, appears tobe expressed at low levels, as can be inferred from the slowkinetics of glycerol fermentation; the fermentation of 8.5 g/Lglycerol took approximately four days (Dharmadi et al., 2006). Ourkey metabolic engineering strategy to increase the rate of glycerolutilization then involves overexpression of the enzymes GldA andDHAK (Fig. 1), an approach expected to accelerate the conversionof glycerol to DHAP. Common glycolytic enzymes known to beexpressed at high levels, and therefore able to support high-carbon fluxes, catalyze the remaining steps in the conversion ofDHAP to pyruvate (Fig. 1). Two fermentative enzymes, also knownto support high fluxes, catalyze the conversion of pyruvate toacetyl coenzyme-A (acetyl-CoA) and acetyl-CoA to acetaldehydeto ethanol: pyruvate formate lyase (PFL) and acetaldehyde/alcoholdehydrogenase (ADH), respectively (Fig. 1). Therefore, overexpres-sion of GldA and DHAK (individually or in combination) shouldincrease the rate of conversion of glycerol to DHAP, which in turnwould increase the rate of synthesis of ethanol and co-productsfrom glycerol.
The metabolic engineering strategies described above aresummarized in Fig. 1. In each case, the engineered pathways thatconvert glycerol to ethanol and co-products are redox-balancedand ATP generating, and thus represented viable alternatives forthe functioning of the cells.
00 30 60 90 120
0.0
Time (h)
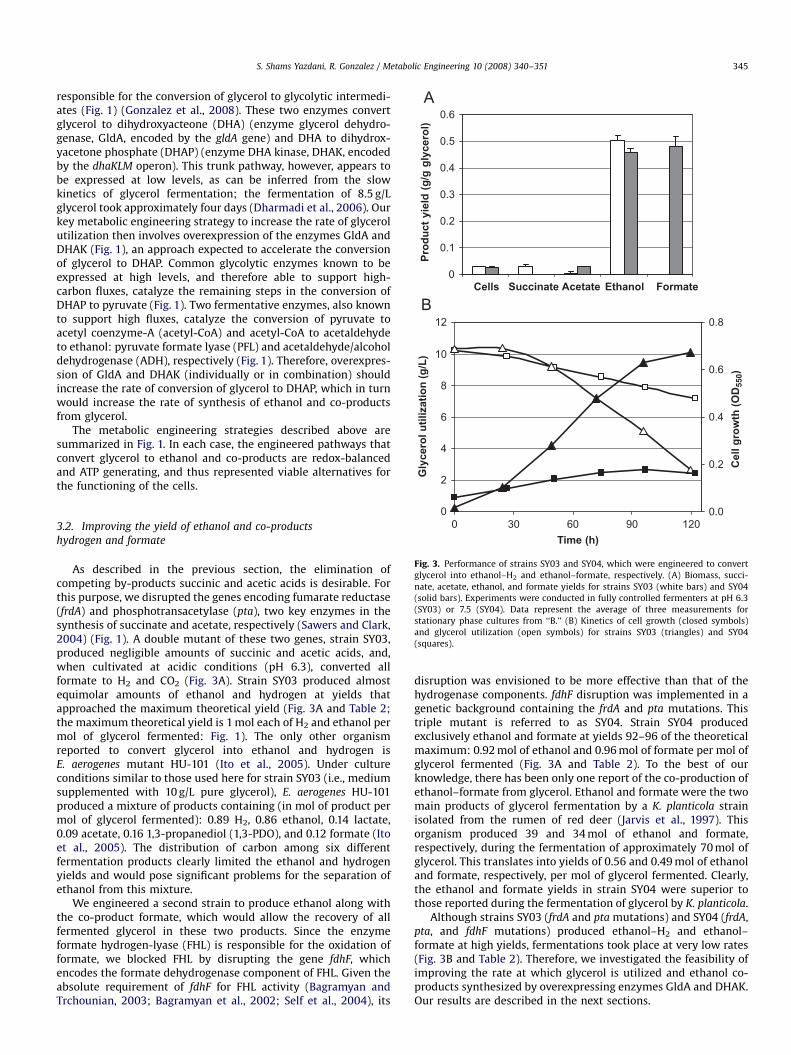
Fig. 3. Performance of strains SY03 and SY04, which were engineered to convert
glycerol into ethanol–H2 and ethanol–formate, respectively. (A) Biomass, succi-
nate, acetate, ethanol, and formate yields for strains SY03 (white bars) and SY04
(solid bars). Experiments were conducted in fully controlled fermenters at pH 6.3
(SY03) or 7.5 (SY04). Data represent the average of three measurements for
stationary phase cultures from ‘‘B.’’ (B) Kinetics of cell growth (closed symbols)
and glycerol utilization (open symbols) for strains SY03 (triangles) and SY04
(squares).
3.2. Improving the yield of ethanol and co-products
hydrogen and formate
As described in the previous section, the elimination ofcompeting by-products succinic and acetic acids is desirable. Forthis purpose, we disrupted the genes encoding fumarate reductase(frdA) and phosphotransacetylase (pta), two key enzymes in thesynthesis of succinate and acetate, respectively (Sawers and Clark,2004) (Fig. 1). A double mutant of these two genes, strain SY03,produced negligible amounts of succinic and acetic acids, and,when cultivated at acidic conditions (pH 6.3), converted allformate to H2 and CO2 (Fig. 3A). Strain SY03 produced almostequimolar amounts of ethanol and hydrogen at yields thatapproached the maximum theoretical yield (Fig. 3A and Table 2;the maximum theoretical yield is 1 mol each of H2 and ethanol permol of glycerol fermented: Fig. 1). The only other organismreported to convert glycerol into ethanol and hydrogen isE. aerogenes mutant HU-101 (Ito et al., 2005). Under cultureconditions similar to those used here for strain SY03 (i.e., mediumsupplemented with 10 g/L pure glycerol), E. aerogenes HU-101produced a mixture of products containing (in mol of product permol of glycerol fermented): 0.89 H2, 0.86 ethanol, 0.14 lactate,0.09 acetate, 0.16 1,3-propanediol (1,3-PDO), and 0.12 formate (Itoet al., 2005). The distribution of carbon among six differentfermentation products clearly limited the ethanol and hydrogenyields and would pose significant problems for the separation ofethanol from this mixture.
We engineered a second strain to produce ethanol along withthe co-product formate, which would allow the recovery of allfermented glycerol in these two products. Since the enzymeformate hydrogen-lyase (FHL) is responsible for the oxidation offormate, we blocked FHL by disrupting the gene fdhF, whichencodes the formate dehydrogenase component of FHL. Given theabsolute requirement of fdhF for FHL activity (Bagramyan andTrchounian, 2003; Bagramyan et al., 2002; Self et al., 2004), its
disruption was envisioned to be more effective than that of thehydrogenase components. fdhF disruption was implemented in agenetic background containing the frdA and pta mutations. Thistriple mutant is referred to as SY04. Strain SY04 producedexclusively ethanol and formate at yields 92–96 of the theoreticalmaximum: 0.92 mol of ethanol and 0.96 mol of formate per mol ofglycerol fermented (Fig. 3A and Table 2). To the best of ourknowledge, there has been only one report of the co-production ofethanol–formate from glycerol. Ethanol and formate were the twomain products of glycerol fermentation by a K. planticola strainisolated from the rumen of red deer (Jarvis et al., 1997). Thisorganism produced 39 and 34 mol of ethanol and formate,respectively, during the fermentation of approximately 70 mol ofglycerol. This translates into yields of 0.56 and 0.49 mol of ethanoland formate, respectively, per mol of glycerol fermented. Clearly,the ethanol and formate yields in strain SY04 were superior tothose reported during the fermentation of glycerol by K. planticola.
Although strains SY03 (frdA and pta mutations) and SY04 (frdA,pta, and fdhF mutations) produced ethanol–H2 and ethanol–formate at high yields, fermentations took place at very low rates(Fig. 3B and Table 2). Therefore, we investigated the feasibility ofimproving the rate at which glycerol is utilized and ethanol co-products synthesized by overexpressing enzymes GldA and DHAK.Our results are described in the next sections.

ARTICLE IN PRESS
Ta
ble
2Fe
rme
nta
tio
np
ara
me
ters
for
cell
gro
wth
,g
lyce
rol
uti
liza
tio
n,
an
dp
rod
uct
syn
the
sis
Stra
ins
Co
nd
itio
na
Ca
rbo
n
reco
ve
ryb
Yie
ld(m
mo
lep
rod
uct
/mm
ole
gly
cero
l)M
ax
spe
cifi
cp
rod
uct
ivit
y(m
mo
les/
gce
lls/
h)c
Ma
xv
olu
me
tric
pro
du
ctiv
ity
(mm
ole
s/L/
h)d
Ce
lls
Eth
an
ol
Form
ate
H2
Ce
lls
Gly
cero
lE
tha
no
lFo
rma
teH
2C
ell
sG
lyce
rol
Eth
an
ol
Form
ate
H2
SY0
3p
H6
.31
06
0.1
11
.01
–1
.02
2.4
98
.07
7.6
7–
7.9
00
.13
1.0
91
.10
–1
.11
SY0
4p
H7.
51
02
0.1
00
.92
0.9
6–
0.7
38
.33
7.7
39
.96
–0
.02
0.3
30
.33
0.3
9–
MG
16
55
(pZ
SB
lan
k)
pH
7.5
96
0.1
01
.00
0.7
5–
1.4
58
.96
7.8
48
.63
–0
.09
0.7
80
.83
0.7
5–
MG
16
55
(pZ
SK
LMg
ldA
)p
H7.
58
40
.14
0.9
10
.85
–3
.39
12
.62
8.7
07
.88
–0
.24
2.0
92
.06
1.8
0–
SY0
4(p
ZS
KLM
gld
A)
pH
7.5
10
20
.13
1.0
40
.92
–5
.23
15
.85
16
.02
19
.67
–0
.55
3.5
83
.58
3.1
8–
SY0
3(p
ZS
KLM
gld
A)
pH
6.3
10
30
.10
1.0
2–
1.0
22
.41
6.1
68
.16
–8
.17
0.1
40
.91
0.9
4–
0.9
4
MG
16
55
(pZ
SK
LMg
ldA
)p
H6
.39
40
.13
0.9
0–
0.9
11
.91
11.1
611
.14
–11
.16
0.2
11
.89
1.7
0–
1.7
0
MG
16
55
pH
6.0
,a
irin
he
ad
spa
ce1
03
0.2
60
.75
–0
.83
6.2
68
.30
6.8
9–
7.4
80
.90
2.9
62
.48
–2
.61
SY0
3p
H6
.0,
air
inh
ea
dsp
ace
97
0.2
60
.82
–0
.88
6.2
86
.13
5.6
3–
6.2
80
.64
2.5
32
.22
–2
.33
SY0
3(p
ZS
KLM
gld
A)
pH
6.0
,a
irin
he
ad
spa
ce9
90
.28
0.9
6–
0.9
66
.04
9.6
58
.82
–8
.84
1.1
94
.68
4.6
5–
4.6
5
SY0
4(p
ZS
KLM
gld
A)
pH
7.5
,10
g/L
cru
de
gly
cero
l9
80
.13
0.9
70
.93
–4
.03
20
.26
15
.83
18.3
4–
0.3
93
.28
3.3
93
.00
–
SY0
4(p
ZS
KLM
gld
A)
pH
7.5
,2
0g
/Lcr
ud
eg
lyce
rol
10
00
.09
1.0
20
.88
–3
.79
29
.54
27
.18
31
.96
–0
.55
3.4
13
.52
2.9
7–
aC
ell
sw
ere
cult
iva
ted
usi
ng
the
me
diu
ma
nd
con
dit
ion
sd
esc
rib
ed
inth
eM
ate
ria
lsa
nd
Met
ho
ds
sect
ion
.U
nle
sso
the
rwis
esp
eci
fie
d,
the
cult
ure
sw
ere
spa
rge
dw
ith
arg
on
an
d1
0g
/Lp
ure
gly
cero
lw
as
use
da
sca
rbo
n
sou
rce
.b
Ca
rbo
nre
cov
ery
acc
ou
nts
for
the
pe
rce
nta
ge
of
carb
on
reco
ve
red
ina
llp
rod
uct
sa
nd
bio
ma
ss.
cM
ax
imu
msp
eci
fic
pro
du
ctiv
itie
s,in
mm
ol
pe
rg
ram
of
cell
sp
er
ho
ur,
we
reca
lcu
late
db
yta
kin
gin
toa
cco
un
tth
ein
terv
al
at
wh
ich
ma
xim
um
mm
ol
of
sub
stra
tew
ere
con
sum
ed
an
dce
lls
an
dp
rod
uct
sfo
rme
d.
Th
e
con
cen
tra
tio
no
fce
lls
du
rin
gth
eg
ive
np
eri
od
of
tim
eis
calc
ula
ted
as
tim
eav
era
ge
.d
Ma
xim
um
vo
lum
etr
icp
rod
uct
ivit
ies
are
rep
ort
ed
for
the
cult
iva
tio
np
eri
od
at
wh
ich
the
ma
xim
um
mm
ol
of
sub
stra
tew
ere
con
sum
ed
an
dce
lls
an
dp
rod
uct
sfo
rme
dp
er
lite
rp
er
ho
ur.
S. Shams Yazdani, R. Gonzalez / Metabolic Engineering 10 (2008) 340–351346
3.3. Improving the rate of glycerol utilization: effect of
overexpressing glycerol dehydrogenase and dihydroxyacetone
kinase activities
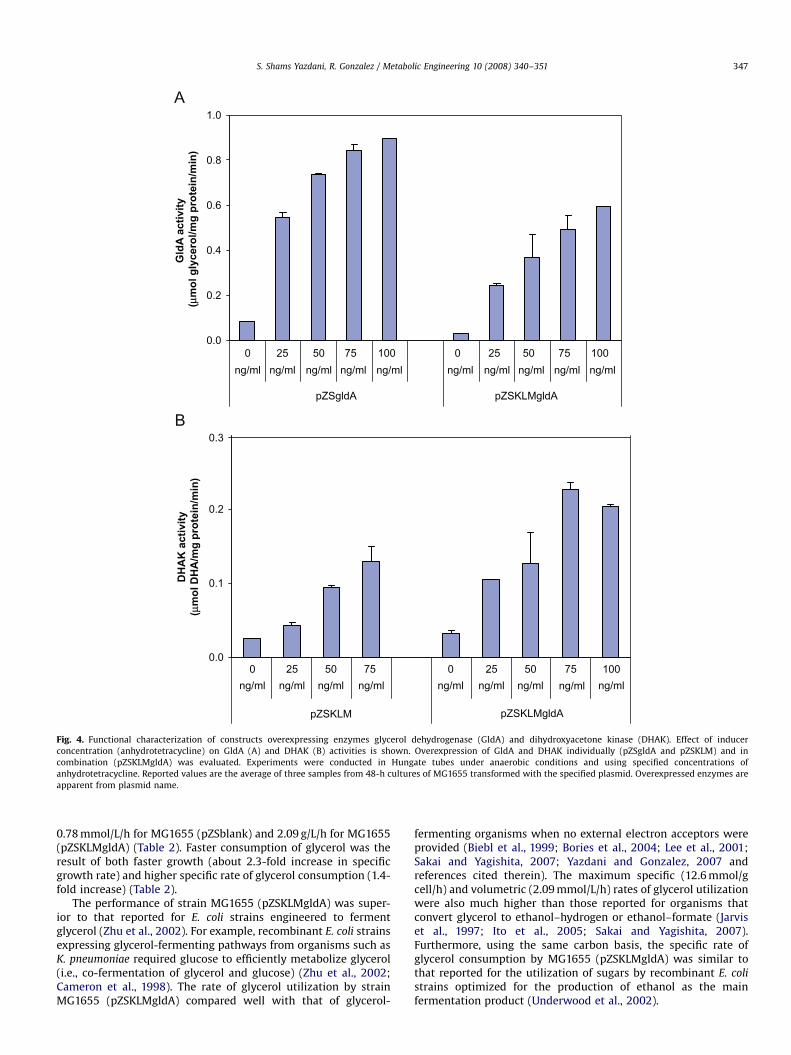
To assess the effect of overexpressing glycerol dehydrogenase(GldA, encoded by the gene gldA) and dihydroxyacetone kinase(DHAK, encoded by the operon dhaKLM) on the kinetics of glycerolfermentation, we used the following plasmids that express thesetwo enzymes individually and in combination: pZSgldA (expres-sing glycerol dehydrogenase), pZSKLM (expressing dihydroxyace-tone kinase), and pZSKLMgldA (expressing both glyceroldehydrogenase and dihydroxyacetone kinase). Plasmid pZSblankwas used as a control and it contains the ‘‘backbone’’ of theplasmids described above (see Fig. 2, Table 1, and Materials and
Methods section for details about plasmids and their construc-tion). We conducted a functional characterization of each of theseconstructs by measuring the changes in GldA and DHAK activitiesin response to different concentrations of inducer anhydrotetra-cycline (Fig. 4). These experiments were carried out in Hungatetubes under anaerobic conditions (see Materials and Methods). Thereported values represent the average for 48-h cultures of strainMG1655 transformed with each of the above plasmids. Weobserved a 10- to 20-fold increase in the activity of GldA,depending on the construct and inducer concentration (Fig. 4A).Overexpression of the dhaKLM operon, on the other hand, resultedin up to a 5- to 6-fold increase in PEP-dependent DHAK activities(Fig. 4B). These results demonstrate the tunability of GldA andDHAK activities using the above constructs.
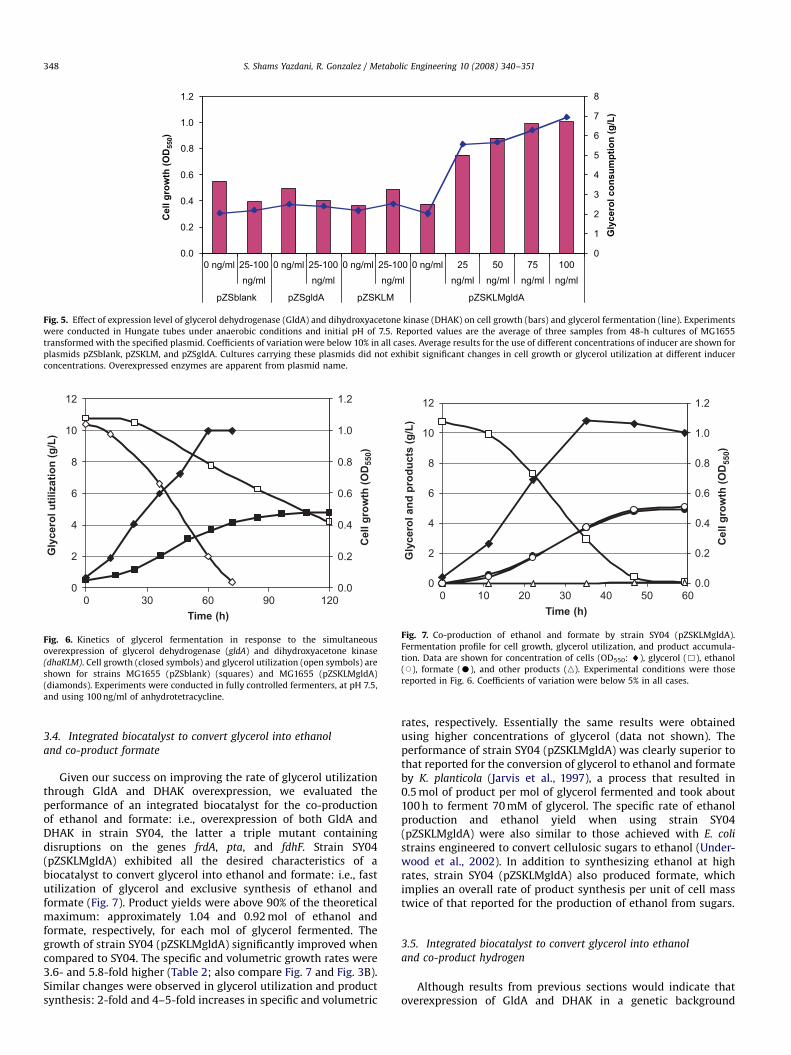
The studies described above also provided an initial assess-ment on how cell growth and glycerol fermentation are affectedby the overexpression of GldA and DHAK. We observed a 3.4-foldincrease in the amount of glycerol fermented by 48-h Hungatetube cultures when these enzymes were simultaneously over-expressed from plasmid pZSKLMgldA (Fig. 5). However, individualoverexpression of either GldA or DHAK did not have a significanteffect on glycerol fermentation (Fig. 5). We speculate that thisbehavior was due to the low activities of GldA and DHAK in wild-type E. coli: 6.6 and 1.7 mmol/g cell/h, respectively. In fact, theseactivities are very similar to the rate at which glycerol is utilizedby MG1655 (5.6 mmol/g cell/h) and therefore might be a limitingfactor in glycerol metabolism. As an alternative explanation, theindividual overexpression of these enzymes could lead to ametabolic imbalance, which in turn would result in the accumu-lation of toxic intermediates. However, the 1D 1H-NMR (nuclearmagnetic resonance) spectra of the extracellular media of culturesoverexpressing GldA and DHAK, individually or in combination,did not show evidence of DHA or methylglyoxal accumulation(data not shown). These are likely intermediates that could begenerated by the imbalanced metabolism of glycerol (Booth,2005). Therefore, the lack of changes in glycolytic fluxes upon theindividual overexpression of either GldA or DHAK does not appearto be due to the creation of a metabolic imbalance, but rather tothe low activities of both enzymes, which in turn required theirsimultaneous overexpression. Our findings suggest a metaboliccontrol structure in which GldA and DHAK share the control of theglycolytic flux during fermentative utilization of glycerol.
To further assess the impact of GldA and DHAK overexpression,we conducted fermentations in a more controlled environment(fully controlled fermenters). Fig. 6 shows complete profiles forstrains MG1655 (pZSblank) and MG1655 (pZSKLMgldA). Theadvantages of GldA and DHAK overexpression are very evident.While a culture of strain MG1655 (pZSKLMgldA) consumedalmost 8 g/L of glycerol when reaching stationary phase at 60 h,the reference strain MG1655 (pZSblank) only consumed 6.5 g/Lglycerol over a period of 120 h (Fig. 6). This translates into a 2.7-fold increase in the volumetric rate of glycerol consumption:
ARTICLE IN PRESS
0.0
0.2
0.4
0.6
0.8
1.0
0 25 50 75 100 0 25 50 75 100
Gld
A ac
tivity
( µm
ol g
lyce
rol/m
g pr
otei
n/m
in)
0.0
0.1
0.2
0.3
0 25 50 75 0 25 50 75 100
pZSKLMgldA
DH
AK
act
ivity
( µm
ol D
HA
/mg
prot
ein/
min
)
ng/ml ng/ml ng/ml ng/ml ng/ml ng/mlng/ml ng/ml ng/ml ng/ml
pZSKLMgldApZSgldA
pZSKLM
ng/ml ng/ml ng/ml ng/ml ng/ml ng/ml ng/ml ng/mlng/ml
Fig. 4. Functional characterization of constructs overexpressing enzymes glycerol dehydrogenase (GldA) and dihydroxyacetone kinase (DHAK). Effect of inducer
concentration (anhydrotetracycline) on GldA (A) and DHAK (B) activities is shown. Overexpression of GldA and DHAK individually (pZSgldA and pZSKLM) and in
combination (pZSKLMgldA) was evaluated. Experiments were conducted in Hungate tubes under anaerobic conditions and using specified concentrations of
anhydrotetracycline. Reported values are the average of three samples from 48-h cultures of MG1655 transformed with the specified plasmid. Overexpressed enzymes are
apparent from plasmid name.
S. Shams Yazdani, R. Gonzalez / Metabolic Engineering 10 (2008) 340–351 347
0.78 mmol/L/h for MG1655 (pZSblank) and 2.09 g/L/h for MG1655(pZSKLMgldA) (Table 2). Faster consumption of glycerol was theresult of both faster growth (about 2.3-fold increase in specificgrowth rate) and higher specific rate of glycerol consumption (1.4-fold increase) (Table 2).
The performance of strain MG1655 (pZSKLMgldA) was super-ior to that reported for E. coli strains engineered to fermentglycerol (Zhu et al., 2002). For example, recombinant E. coli strainsexpressing glycerol-fermenting pathways from organisms such asK. pneumoniae required glucose to efficiently metabolize glycerol(i.e., co-fermentation of glycerol and glucose) (Zhu et al., 2002;Cameron et al., 1998). The rate of glycerol utilization by strainMG1655 (pZSKLMgldA) compared well with that of glycerol-
fermenting organisms when no external electron acceptors wereprovided (Biebl et al., 1999; Bories et al., 2004; Lee et al., 2001;Sakai and Yagishita, 2007; Yazdani and Gonzalez, 2007 andreferences cited therein). The maximum specific (12.6 mmol/gcell/h) and volumetric (2.09 mmol/L/h) rates of glycerol utilizationwere also much higher than those reported for organisms thatconvert glycerol to ethanol–hydrogen or ethanol–formate (Jarviset al., 1997; Ito et al., 2005; Sakai and Yagishita, 2007).Furthermore, using the same carbon basis, the specific rate ofglycerol consumption by MG1655 (pZSKLMgldA) was similar tothat reported for the utilization of sugars by recombinant E. coli
strains optimized for the production of ethanol as the mainfermentation product (Underwood et al., 2002).
ARTICLE IN PRESS
0.0
0.2
0.4
0.6
0.8
1.0
1.2
0 ng/ml 25-100 0 ng/ml 25-100 0 ng/ml 25-100 0 ng/ml 25 50 75 100ng/ml ng/ml ng/ml ng/ml ng/ml ng/ml ng/ml
pZSblank pZSgldA pZSKLM pZSKLMgldA
Cel
l gro
wth
(OD
550)
0
1
2
3
4
5
6
7
8
Gly
cero
l con
sum
ptio
n (g
/L)
Fig. 5. Effect of expression level of glycerol dehydrogenase (GldA) and dihydroxyacetone kinase (DHAK) on cell growth (bars) and glycerol fermentation (line). Experiments
were conducted in Hungate tubes under anaerobic conditions and initial pH of 7.5. Reported values are the average of three samples from 48-h cultures of MG1655
transformed with the specified plasmid. Coefficients of variation were below 10% in all cases. Average results for the use of different concentrations of inducer are shown for
plasmids pZSblank, pZSKLM, and pZSgldA. Cultures carrying these plasmids did not exhibit significant changes in cell growth or glycerol utilization at different inducer
concentrations. Overexpressed enzymes are apparent from plasmid name.
0
2
4
6
8
10
12
0 30 60 90 120Time (h)
Gly
cero
l util
izat
ion
(g/L
)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Cel
l gro
wth
(OD
550)
Fig. 6. Kinetics of glycerol fermentation in response to the simultaneous
overexpression of glycerol dehydrogenase (gldA) and dihydroxyacetone kinase
(dhaKLM). Cell growth (closed symbols) and glycerol utilization (open symbols) are
shown for strains MG1655 (pZSblank) (squares) and MG1655 (pZSKLMgldA)
(diamonds). Experiments were conducted in fully controlled fermenters, at pH 7.5,
and using 100 ng/ml of anhydrotetracycline.
0
2
4
6
8
10
12
0 10 20 4030 50 60
Gly
cero
l and
pro
duct
s (g
/L)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
Cel
l gro
wth
(OD
550)
Time (h)
Fig. 7. Co-production of ethanol and formate by strain SY04 (pZSKLMgldA).
Fermentation profile for cell growth, glycerol utilization, and product accumula-
tion. Data are shown for concentration of cells (OD550: ~), glycerol (&), ethanol
(J), formate (K), and other products (n). Experimental conditions were those
reported in Fig. 6. Coefficients of variation were below 5% in all cases.
S. Shams Yazdani, R. Gonzalez / Metabolic Engineering 10 (2008) 340–351348
3.4. Integrated biocatalyst to convert glycerol into ethanol
and co-product formate
Given our success on improving the rate of glycerol utilizationthrough GldA and DHAK overexpression, we evaluated theperformance of an integrated biocatalyst for the co-productionof ethanol and formate: i.e., overexpression of both GldA andDHAK in strain SY04, the latter a triple mutant containingdisruptions on the genes frdA, pta, and fdhF. Strain SY04(pZSKLMgldA) exhibited all the desired characteristics of abiocatalyst to convert glycerol into ethanol and formate: i.e., fastutilization of glycerol and exclusive synthesis of ethanol andformate (Fig. 7). Product yields were above 90% of the theoreticalmaximum: approximately 1.04 and 0.92 mol of ethanol andformate, respectively, for each mol of glycerol fermented. Thegrowth of strain SY04 (pZSKLMgldA) significantly improved whencompared to SY04. The specific and volumetric growth rates were3.6- and 5.8-fold higher (Table 2; also compare Fig. 7 and Fig. 3B).Similar changes were observed in glycerol utilization and productsynthesis: 2-fold and 4–5-fold increases in specific and volumetric
rates, respectively. Essentially the same results were obtainedusing higher concentrations of glycerol (data not shown). Theperformance of strain SY04 (pZSKLMgldA) was clearly superior tothat reported for the conversion of glycerol to ethanol and formateby K. planticola (Jarvis et al., 1997), a process that resulted in0.5 mol of product per mol of glycerol fermented and took about100 h to ferment 70 mM of glycerol. The specific rate of ethanolproduction and ethanol yield when using strain SY04(pZSKLMgldA) were also similar to those achieved with E. coli
strains engineered to convert cellulosic sugars to ethanol (Under-wood et al., 2002). In addition to synthesizing ethanol at highrates, strain SY04 (pZSKLMgldA) also produced formate, whichimplies an overall rate of product synthesis per unit of cell masstwice of that reported for the production of ethanol from sugars.
3.5. Integrated biocatalyst to convert glycerol into ethanol
and co-product hydrogen
Although results from previous sections would indicate thatoverexpression of GldA and DHAK in a genetic background
ARTICLE IN PRESS
S. Shams Yazdani, R. Gonzalez / Metabolic Engineering 10 (2008) 340–351 349
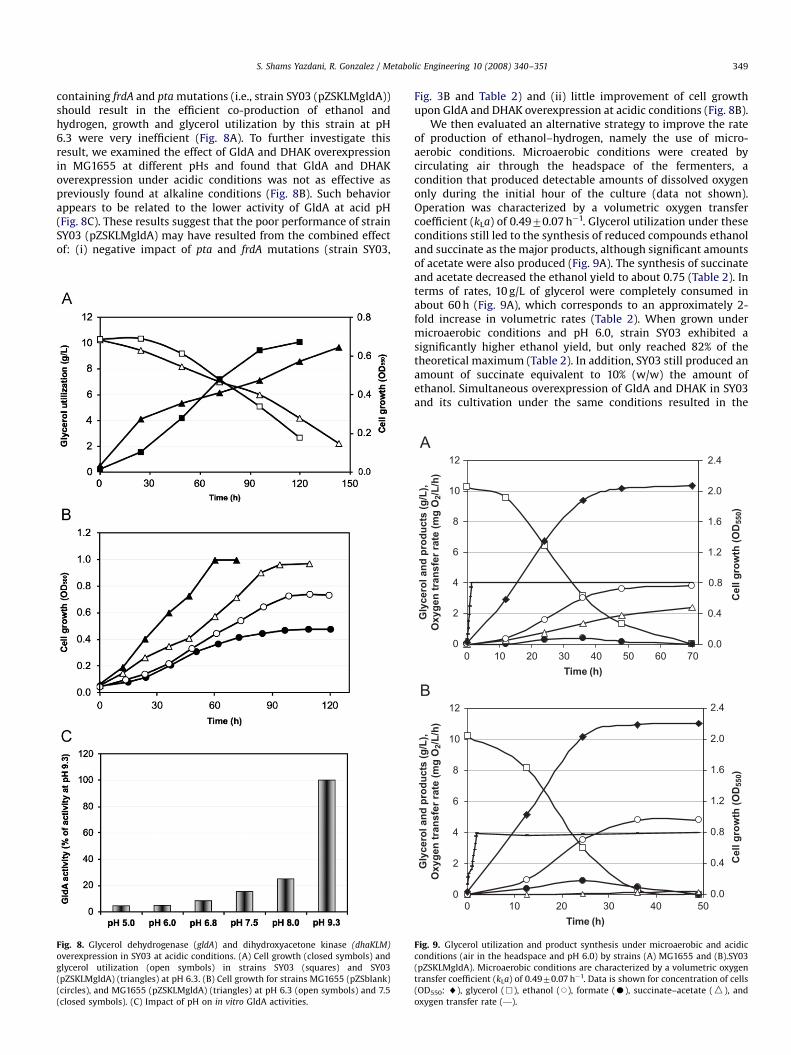
containing frdA and pta mutations (i.e., strain SY03 (pZSKLMgldA))should result in the efficient co-production of ethanol andhydrogen, growth and glycerol utilization by this strain at pH6.3 were very inefficient (Fig. 8A). To further investigate thisresult, we examined the effect of GldA and DHAK overexpressionin MG1655 at different pHs and found that GldA and DHAKoverexpression under acidic conditions was not as effective aspreviously found at alkaline conditions (Fig. 8B). Such behaviorappears to be related to the lower activity of GldA at acid pH(Fig. 8C). These results suggest that the poor performance of strainSY03 (pZSKLMgldA) may have resulted from the combined effectof: (i) negative impact of pta and frdA mutations (strain SY03,
Fig. 8. Glycerol dehydrogenase (gldA) and dihydroxyacetone kinase (dhaKLM)
overexpression in SY03 at acidic conditions. (A) Cell growth (closed symbols) and
glycerol utilization (open symbols) in strains SY03 (squares) and SY03
(pZSKLMgldA) (triangles) at pH 6.3. (B) Cell growth for strains MG1655 (pZSblank)
(circles), and MG1655 (pZSKLMgldA) (triangles) at pH 6.3 (open symbols) and 7.5
(closed symbols). (C) Impact of pH on in vitro GldA activities.
Fig. 3B and Table 2) and (ii) little improvement of cell growthupon GldA and DHAK overexpression at acidic conditions (Fig. 8B).
We then evaluated an alternative strategy to improve the rateof production of ethanol–hydrogen, namely the use of micro-aerobic conditions. Microaerobic conditions were created bycirculating air through the headspace of the fermenters, acondition that produced detectable amounts of dissolved oxygenonly during the initial hour of the culture (data not shown).Operation was characterized by a volumetric oxygen transfercoefficient (kLa) of 0.4970.07 h�1. Glycerol utilization under theseconditions still led to the synthesis of reduced compounds ethanoland succinate as the major products, although significant amountsof acetate were also produced (Fig. 9A). The synthesis of succinateand acetate decreased the ethanol yield to about 0.75 (Table 2). Interms of rates, 10 g/L of glycerol were completely consumed inabout 60 h (Fig. 9A), which corresponds to an approximately 2-fold increase in volumetric rates (Table 2). When grown undermicroaerobic conditions and pH 6.0, strain SY03 exhibited asignificantly higher ethanol yield, but only reached 82% of thetheoretical maximum (Table 2). In addition, SY03 still produced anamount of succinate equivalent to 10% (w/w) the amount ofethanol. Simultaneous overexpression of GldA and DHAK in SY03and its cultivation under the same conditions resulted in the
0
2
4
6
8
10
12
0 10 20 30 40 50Time (h)
0 10 20 30 40 50 60 70Time (h)
Gly
cero
l and
pro
duct
s (g
/L),
Oxy
gen
tran
sfer
rate
(mg
O2/
L/h)
0.0
0.4
0.8
1.2
1.6
2.0
2.4
Cel
l gro
wth
(OD
550)
0.0
0.4
0.8
1.2
1.6
2.0
2.4
Cel
l gro
wth
(OD
550)
0
2
4
6
8
10
12
Gly
cero
l and
pro
duct
s (g
/L),
Oxy
gen
tran
sfer
rate
(mg
O2/
L/h)
Fig. 9. Glycerol utilization and product synthesis under microaerobic and acidic
conditions (air in the headspace and pH 6.0) by strains (A) MG1655 and (B).SY03
(pZSKLMgldA). Microaerobic conditions are characterized by a volumetric oxygen
transfer coefficient (kLa) of 0.4970.07 h�1. Data is shown for concentration of cells
(OD550: ~), glycerol (&), ethanol (J), formate (K), succinate–acetate (W), and
oxygen transfer rate (—).
ARTICLE IN PRESS
S. Shams Yazdani, R. Gonzalez / Metabolic Engineering 10 (2008) 340–351350
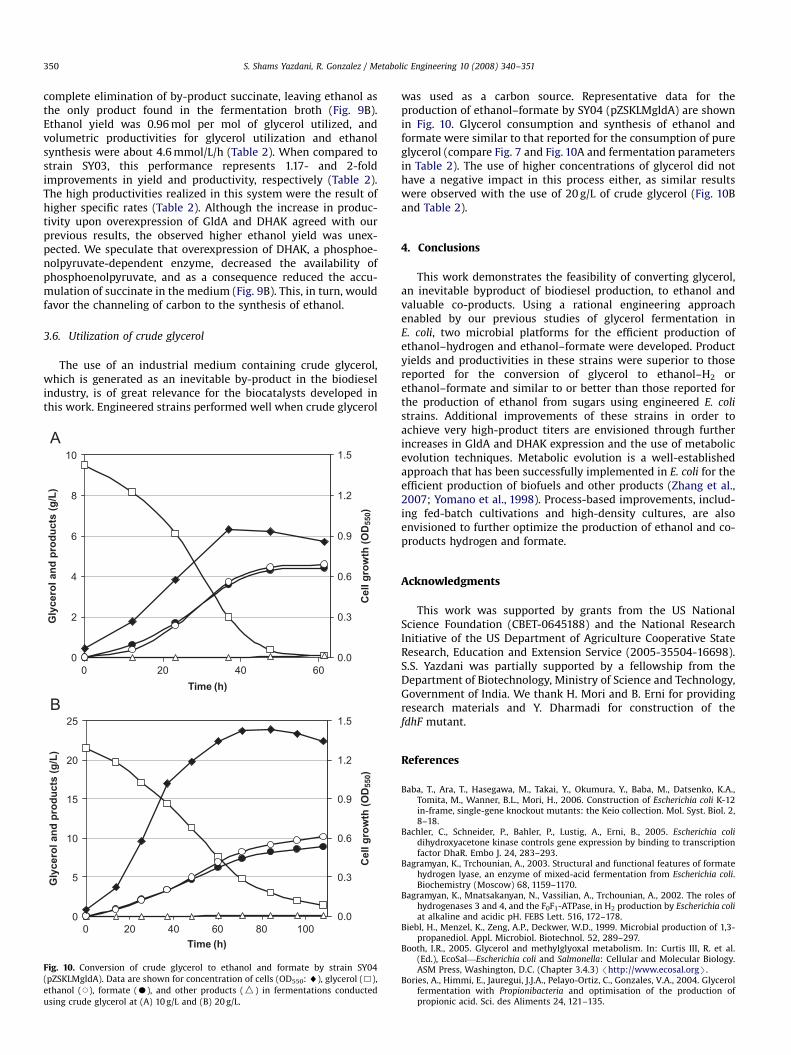
complete elimination of by-product succinate, leaving ethanol asthe only product found in the fermentation broth (Fig. 9B).Ethanol yield was 0.96 mol per mol of glycerol utilized, andvolumetric productivities for glycerol utilization and ethanolsynthesis were about 4.6 mmol/L/h (Table 2). When compared tostrain SY03, this performance represents 1.17- and 2-foldimprovements in yield and productivity, respectively (Table 2).The high productivities realized in this system were the result ofhigher specific rates (Table 2). Although the increase in produc-tivity upon overexpression of GldA and DHAK agreed with ourprevious results, the observed higher ethanol yield was unex-pected. We speculate that overexpression of DHAK, a phosphoe-nolpyruvate-dependent enzyme, decreased the availability ofphosphoenolpyruvate, and as a consequence reduced the accu-mulation of succinate in the medium (Fig. 9B). This, in turn, wouldfavor the channeling of carbon to the synthesis of ethanol.
3.6. Utilization of crude glycerol
The use of an industrial medium containing crude glycerol,which is generated as an inevitable by-product in the biodieselindustry, is of great relevance for the biocatalysts developed inthis work. Engineered strains performed well when crude glycerol
0
2
4
6
8
10
0 20 40 60
Gly
cero
l and
pro
duct
s (g
/L)
0.0
0.3
0.6
0.9
1.2
1.5
Cel
l gro
wth
(OD
550)
0
5
10
15
20
25
0 20 40 60 80 100
Gly
cero
l and
pro
duct
s (g
/L)
0.0
0.3
0.6
0.9
1.2
1.5
Cel
l gro
wth
(OD
550)
Time (h)
Time (h)
Fig. 10. Conversion of crude glycerol to ethanol and formate by strain SY04
(pZSKLMgldA). Data are shown for concentration of cells (OD550: ~), glycerol (&),
ethanol (J), formate (K), and other products (W) in fermentations conducted
using crude glycerol at (A) 10 g/L and (B) 20 g/L.
was used as a carbon source. Representative data for theproduction of ethanol–formate by SY04 (pZSKLMgldA) are shownin Fig. 10. Glycerol consumption and synthesis of ethanol andformate were similar to that reported for the consumption of pureglycerol (compare Fig. 7 and Fig. 10A and fermentation parametersin Table 2). The use of higher concentrations of glycerol did nothave a negative impact in this process either, as similar resultswere observed with the use of 20 g/L of crude glycerol (Fig. 10Band Table 2).
4. Conclusions
This work demonstrates the feasibility of converting glycerol,an inevitable byproduct of biodiesel production, to ethanol andvaluable co-products. Using a rational engineering approachenabled by our previous studies of glycerol fermentation inE. coli, two microbial platforms for the efficient production ofethanol–hydrogen and ethanol–formate were developed. Productyields and productivities in these strains were superior to thosereported for the conversion of glycerol to ethanol–H2 orethanol–formate and similar to or better than those reported forthe production of ethanol from sugars using engineered E. coli
strains. Additional improvements of these strains in order toachieve very high-product titers are envisioned through furtherincreases in GldA and DHAK expression and the use of metabolicevolution techniques. Metabolic evolution is a well-establishedapproach that has been successfully implemented in E. coli for theefficient production of biofuels and other products (Zhang et al.,2007; Yomano et al., 1998). Process-based improvements, includ-ing fed-batch cultivations and high-density cultures, are alsoenvisioned to further optimize the production of ethanol and co-products hydrogen and formate.
Acknowledgments
This work was supported by grants from the US NationalScience Foundation (CBET-0645188) and the National ResearchInitiative of the US Department of Agriculture Cooperative StateResearch, Education and Extension Service (2005-35504-16698).S.S. Yazdani was partially supported by a fellowship from theDepartment of Biotechnology, Ministry of Science and Technology,Government of India. We thank H. Mori and B. Erni for providingresearch materials and Y. Dharmadi for construction of thefdhF mutant.
References
Baba, T., Ara, T., Hasegawa, M., Takai, Y., Okumura, Y., Baba, M., Datsenko, K.A.,Tomita, M., Wanner, B.L., Mori, H., 2006. Construction of Escherichia coli K-12in-frame, single-gene knockout mutants: the Keio collection. Mol. Syst. Biol. 2,8–18.
Bachler, C., Schneider, P., Bahler, P., Lustig, A., Erni, B., 2005. Escherichia colidihydroxyacetone kinase controls gene expression by binding to transcriptionfactor DhaR. Embo J. 24, 283–293.
Bagramyan, K., Trchounian, A., 2003. Structural and functional features of formatehydrogen lyase, an enzyme of mixed-acid fermentation from Escherichia coli.Biochemistry (Moscow) 68, 1159–1170.
Bagramyan, K., Mnatsakanyan, N., Vassilian, A., Trchounian, A., 2002. The roles ofhydrogenases 3 and 4, and the F0F1-ATPase, in H2 production by Escherichia coliat alkaline and acidic pH. FEBS Lett. 516, 172–178.
Biebl, H., Menzel, K., Zeng, A.P., Deckwer, W.D., 1999. Microbial production of 1,3-propanediol. Appl. Microbiol. Biotechnol. 52, 289–297.
Booth, I.R., 2005. Glycerol and methylglyoxal metabolism. In: Curtis III, R. et al.(Ed.), EcoSal—Escherichia coli and Salmonella: Cellular and Molecular Biology.ASM Press, Washington, D.C. (Chapter 3.4.3) /http://www.ecosal.orgS.
Bories, A., Himmi, E., Jauregui, J.J.A., Pelayo-Ortiz, C., Gonzales, V.A., 2004. Glycerolfermentation with Propionibacteria and optimisation of the production ofpropionic acid. Sci. des Aliments 24, 121–135.

ARTICLE IN PRESS
S. Shams Yazdani, R. Gonzalez / Metabolic Engineering 10 (2008) 340–351 351
Bouvet, O.M., Lenormand, P., Carlier, P., Grimont, P.A., 1994. Phenotypic diversity ofanaerobic glycerol dissimilation shown by seven enterobacterial species. Res.Microbiol. 145, 129–139.
Bouvet, O.M., Lenormand, P., Ageron, E., Grimont, P.A., 1995. Taxonomic diversity ofanaerobic glycerol dissimilation in the Enterobacteriaceae. Res. Microbiol. 146,279–290.
Cameron, D.C., Altaras, N.E., Hoffman, M.L., Shaw, A.J., 1998. Metabolic engineeringof propanediol pathways. Biotechnol. Prog. 14, 116–125.
Datsenko, K.A., Wanner, B.L., 2000. One-step inactivation of chromosomal genes inEscherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 97,6640–6645.
Dharmadi, Y., Gonzalez, R., 2005. A better global resolution function and a noveliterative stochastic search method for optimization of high-performance liquidchromatographic separation. J. Chromatogr. A. 1070, 89–101.
Dharmadi, Y., Murarka, A., Gonzalez, R., 2006. Anaerobic fermentation of glycerolby Escherichia coli: a new platform for metabolic engineering. Biotechnol.Bioeng. 94, 821–829.
Gonzalez, R., Murarka, A., Dharmadi, Y., Yazdani, S.S., 2008. A new model for theanaerobic fermentation of glycerol in enteric bacteria: trunk and auxiliarypathways in Escherichia coli. Metab. Eng. 10, 234–245.
Ito, T., Nakashimada, Y., Senba, K., Matsui, T., Nishio, N., 2005. Hydrogen andethanol production from glycerol-containing wastes discharged after biodieselmanufacturing process. J. Biosci. Bioeng. 100, 260–265.
Jarvis, G.N., Moore, E.R.B., Thiele, J.H., 1997. Formate and ethanol are the majorproducts of glycerol fermentation produced by a Klebsiella planticola strainisolated from red deer. J. Appl. Microbiol. 83, 166–174.
Kang, Y., Durfee, T., Glasner, J.D., Qiu, Y., Frisch, D., Winterberg, K.M., Blattner, F.R.,2004. Systematic mutagenesis of the Escherichia coli genome. J. Bacteriol. 186,4921–4930.
Kornberg, H.L., Reeves, R.E., 1972. Inducible phosphoenolpyruvate-dependenthexose phosphotransferase activities in Escherichia coli. Biochem. J. 128,1339–1344.
Lee, P.C., Lee, W.G., Lee, S.Y., Chang, H.N., 2001. Succinic acid production withreduced by-product formation in the fermentation of Anaerobiospirillumsucciniciproducens using glycerol as a carbon source. Biotechnol. Bioeng. 72,41–48.
Lin, E.C., 1976. Glycerol dissimilation and its regulation in bacteria. Annu. Rev.Microbiol. 30, 535–578.
McCoy, M., 2005. An unlikely impact. Chem. Eng. News 83, 24–26.McCoy, M., 2006. Glycerin surplus. Chem. Eng. News 84, 7–8.Miller, J.H., 1972. Experiments in Molecular Genetics. Cold Spring Harbor
Laboratory Press, Cold Spring Harbor, NY.Murarka, A., Dharmadi, Y., Yazdani, S.S., Gonzalez, R., 2008. Fermentative
utilization of glycerol in Escherichia coli and its implications for the productionof fuels and chemicals. App. Environ. Microbiol. 74, 1124–1135.
Neidhardt, F.C., Bloch, P.L., Smith, D.F., 1974. Culture medium for enterobacteria.J. Bacteriol. 119, 736–747.
Nielsen, J., Villadsen, J., Liden, G., 2003. Bioreaction engineering principles. KluwerAcademic/Plenum Publishers, New York, pp. 60–73.
Osman, Y.A., Conway, T., Bonetti, S.J., Ingram, L.O., 1987. Glycolytic flux inZymomonas mobilis: enzyme and metabolite levels during batch fermentation.J. Bacteriol. 169, 3726–3736.
Quastel, J.H., Stephenson, M., 1925. Further observations on the anaerobic growthof bacteria. Biochem. J. 19, 660–666.
Quastel, J.H., Stephenson, M., Whetham, M.D., 1925. Some reactions of restingbacteria in relation to anaerobic growth. Biochem. J. 14, 304–316.
Sakai, S., Yagishita, T., 2007. Microbial production of hydrogen and ethanol fromglycerol-containing wastes discharged from a biodiesel fuel production plantin a bioelectrochemical reactor with thionine. Biotechnol. Bioeng. 98, 340–348.
Sambrook, J., Fritsch, E.F., Maniatis, T., 1989. Molecular Cloning: A LaboratoryManual, second ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbour,NY.
Sawers, R.G., Clark, D.P., 2004. Fermentative pyruvate and acetyl-coenzyme Ametabolism. In: Curtis III, R. et al. (Ed.), EcoSal—Escherichia coli and Salmonella:Cellular and Molecular Biology. ASM Press, Washington, D.C. (Chapter 3.5.3)/http://www.ecosal.orgS.
Self, W.T., Hasona, A., Shanmugam, K.T., 2004. Expression and regulation of a silentoperon, hyf, coding for hydrogenase 4 isoenzyme in Escherichia coli. J. Bacteriol.186, 580–587.
Shuler, M.L., Kargi, F., 2002. Bioprocess Engineering Basic Concepts, second ed.Prentice-Hall, Upper Saddle River, NJ.
Tao, H., Gonzalez, R., Martinez, A., Rodriguez, M., Ingram, L.O., Preston, J.F.,Shanmugam, K.T., 2001. Engineering a homo-ethanol pathway in Escherichiacoli: increased glycolytic flux and levels of expression of glycolytic genesduring xylose fermentation. J. Bacteriol. 183, 2979–2988.
Truniger, V., Boos, W., 1994. Mapping and cloning of gldA, the structural gene of theEscherichia coli glycerol dehydrogenase. J. Bacteriol. 176, 1796–1800.
Underwood, S.A., Zhou, S., Causey, T.B., Yomano, L.P., Shanmugam, K.T., Ingram, L.O.,2002. Genetic changes to optimize carbon patitioning between ethanol andbiosynthesis in ethanologenic Escherichia coli. Appl. Environ. Microbiol. 68,6263–6272.
Yazdani, S.S., Gonzalez, R., 2007. Anaerobic fermentation of glycerol: a path toeconomic viability for the biofuels industry. Curr. Opin. Biotechnol. 18,213–219.
Yomano, L.P., York, S.W., Ingram, L.O., 1998. Isolation and characterization ofethanol-tolerant mutants of Escherichia coli KO11 for fuel ethanol production.J. Ind. Microbiol. Biotechnol. 20, 132–138.
Zhang, X., Jantama, K., Moore, J.C., Shanmugam, K.T., Ingram, L.O., 2007. Productionof L-alanine by metabolically engineered Escherichia coli. Appl. Microbiol.Biotechnol. 77, 355–366.
Zhu, M.M., Lawman, P.D., Cameron, D.C., 2002. Improving 1,3-propanediolproduction from glycerol in a metabolically engineered Escherichia coli byreducing accumulation of sn-glycerol-3-phosphate. Biotechnol. Prog. 18,694–699.


















