
БЪЛГАРСКА АКАДЕМИЯ НА НАУКИТЕibir.bas.bg/uploads/user/Процедури...
34
БЪЛГАРСКА АКАДЕМИЯ НА НАУКИТЕ ИНСТИТУТ ПО БИОЛОГИЯ И ИМУНОЛОГИЯ НА РАЗМНОЖАВАНЕТО „Акад. К. Братанов” Секция по „Репродуктивни Биотехнологии и Криобиология на Гаметите” Александър Димитров Куков ИЗСЛЕДВАНЕ РОЛЯТА НА СЕЛЕКТИРАНИ СПЕРМАЛНО ПЛАЗМЕНИ БЕЛТЪЦИ ПРИ IN VITRO СЪХРАНЕНИЕ НА СПЕРМАТОЗОИДИ ОТ КОЧ АВТОРЕФЕРАТ на дисертация за придобиване на образователна и научна степен „Доктор” Научен Ръководител: Доц. Мария Иванова-Кичева, дссн София 2011
Transcript of БЪЛГАРСКА АКАДЕМИЯ НА НАУКИТЕibir.bas.bg/uploads/user/Процедури...

БЪЛГАРСКА АКАДЕМИЯ НА НАУКИТЕ
ИНСТИТУТ ПО БИОЛОГИЯ И ИМУНОЛОГИЯ НА РАЗМНОЖАВАНЕТО„Акад. К. Братанов”
Секция по „Репродуктивни Биотехнологии и Криобиология на Гаметите”
Александър Димитров Куков
ИЗСЛЕДВАНЕ РОЛЯТА НА СЕЛЕКТИРАНИ СПЕРМАЛНО ПЛАЗМЕНИ БЕЛТЪЦИ ПРИ IN VITRO СЪХРАНЕНИЕ НА
СПЕРМАТОЗОИДИ ОТ КОЧ
АВТОРЕФЕРАТна дисертация за придобиване на
образователна и научна степен „Доктор”
Научен Ръководител: Доц. Мария Иванова-Кичева, дссн
София 2011

Дисертационния труд е написан на 187 стр., илюстриран с 31 фигури и 26 таблици. В библиографския списъл са цитирани 241 литературни източника, от които 18 на кирилица.
Изказвам искрени благодарности на научният ми ръководител за напътствията и помощта при подготвянето на този дисертационен труд. Благодаря на колегите от Секцията и Института за помощта при заснемането на част от снимковия материал и SCA анализите.

БЪЛГАРСКА АКАДЕМИЯ НА НАУКИТЕ
ИНСТИТУТ ПО БИОЛОГИЯ И ИМУНОЛОГИЯ НА РАЗМНОЖАВАНЕТО„Акад. К. Братанов”
Секция по „Репродуктивни Биотехнологии и Криобиология на Гаметите”
Александър Димитров Куков
ИЗСЛЕДВАНЕ РОЛЯТА НА СЕЛЕКТИРАНИ СПЕРМАЛНО ПЛАЗМЕНИ БЕЛТЪЦИ ПРИ IN VITRO СЪХРАНЕНИЕ НА
СПЕРМАТОЗОИДИ ОТ КОЧ
АВТОРЕФЕРАТна дисертация за придобиване на
образователна и научна степен „Доктор”
Научен Ръководител: Доц. Мария Иванова-Кичева, дссн
София 2011

ИЗПОЛЗВАНИ СЪКРАЩЕНИЯ
СП – спермална плазма; семенна плазмаСПП – спермално плазмени протеини; семенни протеиниИО – изкуствено осеменяванеПМ – плазмена мембранаBSP – говежди семенни протеиниHDL – high-density lipoprotein; ФЛ – фосфолипидиFAA – fertility associating antigenpI – изоелектрична точкаZP – zona pellucidaММ – молекулна масаОПН – остеопонтинICSI – intra cytoplasmic sperm injectionIVF – in vitro fertilization АТФ – аденозин трифосфатСЗО (WHO) – Свтовна Здравна ОрганизацияФС – фосфатидилсеринФЕА – фосфатидилетаноламинCFDA – карбоксифлуоресцин диацетатPI – пропидиев йодидСТС – хлортетрациклин (тест за капацитация)R123 – родамин 123, флуорохром оцветяващ митохондриите и други клетъчни органели.АР – акрозомна реакция.

I.УводЕдна от основните цели на животновъдната практика е максимално
съхранение на биологичния потенциал на гаметите. За постигането на тази цел се правят опити за търсене на ефективни криопротективни субстанции и разработване на нови среди за замразяване и размразяване на семенна течност. Криопротекторите трябва да са вещества, които да не са токсични, да се разтварят във вода и да не причиняват увреждания на биологичните обекти. Освен добре познатите протектори като глицерин, DMSO, някои полиетиленгликоли и яйчен жълтък, учените са насочили своите усилия в търсене на нови подходи за протекция. В този аспект са изследванията върху влиянието на някои спермално плазмени фактори върху функционалността на половите клетки. Някои от спермално плазмените протеини могат да моделират по определен начин плазмената мембрана на сперматозоидите и по този начин да се постигне протекция на клетките от негативното влияние на ниските температури. В тази насока има много непроучени моменти, особено що се касае за сперматозоидите от коч.
Спермалната плазма (СП) е смес от секретите на допълнителните полови жлези и на отделящия се, макар и в малко количество, секрет от надсеменника, семепровода и уретрата. Съдържанието и обемът на СП варира при различните видове селскостопански животни, като това зависи от броя и размера на допълнителните полови жлези (Hafez ESE. 1993). СП съдържа почти всички елементи, характерни за живата материя. СП е много богата на фруктоза, протеини, витамини, лимонена киселина, простагландини, аминокиселини, алкална и кисела фосфатаза, ензими, соли, въглехидрати, хормони и др. субстанции (Hafez ESE. 1993). СП е биологична среда с твърде сложен състав на белтъчните вещества, поради особената роля на спермата във връзка с оплодителния процес. При отделни видове животни белтъците се различават в качествено и количествено отношение. В спермата се съдържат в свободна или в свързана форма всички природни α-аминокиселини. Свободните аминокиселини се съдържат в СП и в незначителни количества в сперматозоидите. В СП се съдържат и вещества с антимикробиологично действие. Това са имуноглобулини, главно от IgA клас. Освен имуноглобулините в СП се намират и други протеини, които имат антимикробно действие. Тези белтъци се отделят в епидидимиса, където съзряват сперматозоидите (Yenugu S. 2004). Антимикробното действие на някои от тях се изразява в свързване към бактериалната стена и повишаване на пропускливостта на последната. Други протеини се разграждат до по-малки деривати, които проникват в бактериалните клетки и инхибират някои метаболитни процеси, като например синтеза на рРНК (Reddy ES. 1979). Добре проучени антимикробни протеини в спермалната плазма са: кателицидин (човешки катионен антимикробен протеин), дефенсин (Palladino MA. 2003; Com E. 2003), лактоферин, спермалния плазмин, семиногелини др. Семиногелина след еякулацията в присъствието на Zn2+ бива фрагментиран и се счита, че тези по-малки фрагменти притежават антибактериално (септично) действие (Bourgeon F. 2004).
В СП има различни хормонални субстанции, като андрогени, естрогени, простагландини, фоликулостимулиращ хормон (FSH), хорион гонадотропин подобен материал, хормони на растежа, инсулин, глюкагон, пролактин, релаксин, тероид-рилизинг хормон и енкефалин. В СП се съдържа спермин,

който притежава цитотоксично действие за лимфоцитите. Спермина играе важна роля при така наречения феномен на спермално плазмената имуносупресия, който е необходимо условие за успешно оплождане.
В спермата са установени редица ензими, които според това, дали са в СП или в сперматозоидите, се разделят на плазмени и интрацелуларни (клетъчни). В СП се сърдъжат протеолитични ензими, аминооксидази, кисела и алкална фосфатаза, 5 - нуклеотидаза, пирофосфатаза, аденозинтрифосфатаза и др. Един от важните ензими в СП е хиалуронидазата, който разлага хиалуроновата киселина. При бозайниците хиалуронидазата играе важна роля при оплождането като фактор, освобождаващ яйцеклетката от външния слой фоликулни клетки и белтъчна материя и обуславящ проникването през оолемата. Друг важен ензим, това е муциназата. Негова функция е да разтапя муцина във влагалищния и в маточния секрети, което улеснява предвижването на сперматозоидите. Каталазата, участва в преобразуването на токсичния за клетките Н2О2 във вода и моларен кислород. Липазата участва в обменните процеси на липоидите. В секрета на простатната жлеза се наблюдава твърде активния ензим фибринолизин (протеолитичен ензим). СП има богато ензимно съдържание и поради това, когато се постави в оптимална температура, в нея стават интензивни ензимни процеси. В резултат на това СП бързо променя своите качества и сперматозоидите умират по–бързо. Ето защо, един от подходите за съхраняване на спермата е при ниски температури, което инхибира до известна степен ензимните процеси.
Свободните радикали и най-вече реактивните кислородни видове имат както полезно, така и вредно въздействие върху сперматозоидите на бозайниците (Murawski M. 2007). Предполага се, че супероксидния анион в определени ниски концентрации играе роля при хиперактивацията и капацитацията на сперматозоидите (de Lamirande E. 1995). От друга страна обаче, високите концентрации на свободните радикали като Н2О2, индуцират липидна пероксидация, която води до клетъчна смърт. Известно е, че плазмената мембрана на сперматозоидите на селскостопанските животни и човека са богати на полиненаситени мастни киселини. Това прави клетките много чувствителни на окислителното въздействие на свободните радикали, които инициират липидна пероксидация в ПМ (Dandekar SP. 2002). Зрелите сперматозоиди притежават ензимни и не-ензимни системи за предпазване от уврежданията на свободните радикали. Също така, СП съдържа високо специализирани антиоксидантни системи, които предпазват мембраните на половите клетки от липидна пероксидация. Тези системи съдържат ензимите супероксид дисмутаза, каталаза и глутатион пероксидаза (Lenzi A. 2002). Освен ензимите, в СП се съдържат витамин Е, аскорбинова киселина и Zn, които имат антиоксидантна функция. В плазмата на заек и човек са открити мембранни гранулки, които се отделят от простатата и са изключително богати на витамин Е (Mourvaki E. 2010). Не е установена все още точната роля на тези гранулки, но се счита, че те могат да участват в антиоксидантната защита на клетките. Прекаленото образуване на свободни радикали в спермата може да играе ключова роля при безплодието при хора и животни (Ford WC. 1997). Предполага се, че реактивните кислородни видове в спермата се образуват основно от неутрофилите и увредените сперматозоиди (Murawski M. 2007). При оплождане сперматозоидите са подложени на атака и от свободните радикали в женския репродуктивен път, в този случай наличието на антиоксидантни защитни системи в СП е от голямо значение. При сперматозоиди от коч са

проведени редица изследвания в този аспект. Така например, изследвано е влиянието на оксидативния стрес върху виталитета на сперматозоиди при in vitro съхранение, като е анализирана антиоксидантна защита на иницииращи апоптоза свободни радикали, но все още няма пълна информация, която да позволи оптимизирането на репродуктивните биотехнологии за нискотемпературно съхранение на сперматозоидите при този вид (Стефанов Р. 2007).
Особено актуални в последните години са проучванията върху наличието на специфични протеини и фактори в СП във връзка с диагностиката на редица заболявания на репродуктивната система при мъжки индивиди. Изследванията върху СП от човек показва, че в нея се съдържа голям арсенал от протеини, които са необходими за нормалната физиология и метаболизъм на сперматозоидите. Някои от тези протеини са изследвани като биомаркери за наличие на различни карциноми. Установено е, че нивата на тези специфични протеини в плазмата зависят от прогресивното развие на тумора. Това са така наречените простатни карциномни протеини, идентифицирани като простатен специфичен антиген, Zn α2-гликопротеин и прогастриксин. Така че, освен физиологичната роля, някои от спермално плазмените протеини могат да са сигнал и за сериозни заболявания (Hassan IM. 2007). Простатен специфичен антиген (PSA) е 33 кДа едноверижен гликопротеин, който принадлежи към голямата група на извънклетъчните серинови протеази. PSA се образува и се секретира от епитела на простатата, и притежава химотрипсин подобна ензимна активност. Основния физиологичен субстрат на ензима са гел-образуващите протеини семиногелин І и ІІ, които се секретират от семенните везикули. PSA е един от белтъците, който се среща в голямо количество в СП (Henttu P. 1992). PSA обикновено присъства в кръвния серум на здрави мъже в много ниски концентрации (до 4 ng/ml). При пациенти с карцином на простатата или други заболявания на жлезата концентрацията на белтъка рязко се повишава. Днес изследването на PSA се счита за ефективен тест при ранното диагностициране на рака на простатата (Schröder FH. 2009). Изследванията в този аспект показват, че е възможно някои белтъци от спермалната плазма да се използват като биомаркери при редица заболявания, свързани с проблеми в мъжката репродукция (Yamakawa K. 2007).
В последните години особено внимание се обръща на протеините от СП и тяхната роля при клетъчната трансформация на сперматозоидите след еякулация. По химичен състав плазмата е един от най-сложните биологични продукти на организма (Братанов K. 1977; Hafez ESE. 1993). Някои СП белтъци повлияват подвижността и преживяемостта на половите клетки, а също така може да окажат влияние върху оплодителния им потенциал (Mortarino M. 1998; Cross NL. 1993; Strzezek J. 2002). Различни спермално плазмени белтъци са описани като инфертилни фактори при коне (Brandon CI. 1999), бик (Bhargava PM. 1986) и човек (Audhya T. 1987). Според авторите някои СПП участват в процеса капацитация (Manjunath P. 2002). Има предположения, че прикрепянето на някои протеини към сперматозоидите, води до забавяне на капацитацията и акрозомната реакция (Fraser GS. 1996; Manjunath P. 1993). Тези протеини са описани като „декапацитиращ фактор”. Killian и колеги описват присъствието на т.нар. "fertility-associated proteins” в СП на бик, които са важни за оплодителната способност на сперматозоидите (Killian GJ. 1993). Протеомните проучвания в последните години показаха, че СПП могат да се

използват като биомаркери при различни заболявания на мъжката репродуктивна система. Barrios и съавтори откриват, че в СП се съдържат протеини, които могат да предпазят сперматозоидите от влиянието на ниските температури при in vitro съхраняване (Barrios B. 2000; Barrios B. 2005). Има данни, че СП е важна за: подържането на мотилитета на сперматозоидите при бик (Baas JW. 1983) и коч (Graham JK. 1994); за подобряването на преживяемостта на сперматозоиди от коч (Maxwell WMC. 1997); за повишаване на устойчивостта на сперматозоиди от нерез срещу температурен шок (Berger T. 1985) и най-вероятно има влияние върху оплодителната способност (Strzezek J. 2002).От друга страна, вредният ефект на СП върху подвижността (Iwamoto T. 1993), преживяемостта (Garcia and Graham 1987) и оцеляването на сперматозоидите след ТШ също са описани в литературата. Тези противоречиви резултати показват, че СП е комплексен флуид съдържащ разнообразни компоненти, които повлияват мотилитета и преживяемостта на сперматозоидите по различни начини. ПМ на сперматозоидите от коч е изключително чувствителна на ниските температури, което обяснява лошите резултати от замразяването и липсата на утвърдени биотехнологии за изкуствено осеменяване със замразена сперма. Съхраняването на молекулната цялост на ПМ е от важно значение за биологичната роля на сперматозоидите от коч, а именно оплождането на яйцеклетката. Митохондриите са много чувствителни органели към ниските температури, като основните увреждания засягат предимно митохондриалните мембрани. Ядрото на сперматозоидите е много компактно и това е структурата, която най-слабо се засяга от влиянието на ниските температури.
Противоречивите данни ни накараха да изследваме какво е влиянието на отделните сепарирани спермално плазмени белтъци върху мотилитета и преживяемостта на сперматозоидите от коч. И дали е възможно някои СПП да се използват като биологични компоненти в криопротективните среди прилагани в ИО на овцете.

ІІ. Цел и задачиЦелта на настоящата дисертационна работа е да се проучи ролята на
селектирани спермално плазмени протеини върху биологичните качества на сперматозоиди от коч, с оглед изучаване на механизма на протекция и преодоляване негативното влиянието на ниските температури при съхраняването на половите клетки в охладено състояние.
За изпълнение на така поставената цел бяха формулирани следните задачи:
1. Ефект на ниските температури върху биологичните качества и адаптационните способности на сперматозоиди от коч при криоконсервация.
2. Хроматографско сепариране на протеини от спермална плазма на коч и получаване на имунен серум
3. Анализ на ролята на селектираните протеини върху функционалността на сперматозоидите от коч при условия на индуциране на температурен шок и съхранение на 50С.
4. Сравнителен анализ върху ролята на селектирани спермално плазмени протеини върху целостта на плазмената мембрана чрез 6CFDA/PI тест и флоуцитометър във връзка с жизнеността на половите клетки след температурен шок и след инкубация на 50С за 24 часа.
5. Анализ върху поведението на вътреклетъчни структури – митохондрии и ДНК при нискотемпературно съхранение на 50С на сперматозоиди от коч, в присъствие на селектирани спермално плазмени протеини.
6. Изкуствено осеменяване на овце с поликомпонентна среда обогатена със селектирани спермално плазмени протеини с доказан протективен ефект.
III.Материал и Методи- Получаване, преценка на сперматологични параметри по стандартни методи и чрез компютърен спермоанализ, SCA анализ.- Подготовка на среди (6А) и буфери за провеждане на експериментите.- Прилагане на биотехнология за криоконсервация на сперматозоиди от коч в пайети.- Изолиране на спермална плазма. Молекулно ситова хроматография на спермално плазмени протеини чрез Sephacryl (FPLC) Течна хроматография (HPLC) на фракции семенни белтъци.- Получаване и имунохимична характеристика на имунен серум срещу протеини от спермална плазма.- Характеристика на селектирани протеини чрез SDS PAGE. - Електротрансфер и Western-имуноблот за определяне на активната форма на каспази 3 и 9. - Прилагане на метод за сребърно оцветяване на полиакриламиден гел.- Морфологични анализи на сперматозоиди чрез тристепенен метод за оцветяване. - Изследване на промяната във фосфолипидната асиметрия при сперматозоиди след криоконсервация чрез анексин V/6CFDA тест.- Изследвания върху интегритета на ПМ чрез прилагане на 6CFDA/PI тест.- Флоуцитометричен анализ на ролята на селектирани спермално плазмени протеини върху интегритета на ПМ, чрез CFDA/PI тест

- Анализ на митохондриалния трансмембранен потенциал чрез прилагане на родамин 123 (R123) тест.- Определяне интегритета на ДНК чрез акридин оранж оцветяване.- Имуноцитохимичен метод.- Изкуствено осеменяване при овце.- Статистически анализ на резултатите.

IV. Резултати и Обсъждане
Задача 1. Ефект на ниските температури върху биологичните качества и адаптационните способности на сперматозоиди от коч при криоконсервацияАнализът на фактическото състояние на въпроса за ролята на ниските
температури върху биологичните качества на сперматозоиди от коч при “in vitro” съхранение на ниски температури показа, че независимо от известните за практиката голямо количество методи за консервиране на сперма, все още проблемът с преодоляването на студовия шок при клетките от този вид не е изучен достатъчно ясно.
С настоящите изследвания са направени сравнителни проучвания върху промени, които настъпват в морфологичната структура на спермалната плазмена мембрана, структурата на митохондриите и ДНК, наличието на каспазна активност при криоконсервация на сперматозоиди от коч.
1.1. Анализ на биологичните параметри – мотилитет, преживяемост и морфология.
За провеждане на експериментите са използвани еякулати с нормални качествени и количествени параметри. Анализът на преживяемостта на сперматозоидите се проведе чрез определяне на началния мотилитет след размразяване и чрез проследяване подвижността на сперматозоидите през 60 минути до 300-тата минута при 390С (Фигура 11).
0
5
10
15
20
25
30
35
40
45
50
10 60 120 180 240 300
минути
%
група А група Б
Фигура 11. Мотилитет и преживяемост на сперматозоиди от коч след размразяване, проследена до 300-тата мин. при 39 0С. Група А - проби с понижена криотолерантност; Група Б - проби с добра криотолерантност.
Големите разлики в преживяемостта на отделните проби ни позволи да ги разделим на две групи - група А - проби с понижена криотолерантност и група Б - проби с добра криотолерантност. Стойностите на началния мотилитет са значително по-ниски на сперматозоидите в пробите с понижена криотолерантност (група А), като на 60-тата, 120-тата и особено на 300-тата мин. от съхранението тази разлика е статистически значима, в сравнение с мотилитета на сперматозоидите от група Б (р<0.05 и р<0.001). При тези изследвания еякулатите, които показват тенденция за добра криотолерантност

на сперматозоидите запазват своя мотилитет до 17.77±3.10 на 300-тата мин (Таблица 2).
Таблица 2. Проследяване на подвижността на сперматозоиди от коч, при
еякулати след получаване и след криоконсервация до 300 минута.проби 10 мин 60 мин 120 мин 180 мин 240 мин 300 мин
група А
(n=7)
26.58±2.50 19.5±3.25a 16.61±4.8a 14.55±2.50 10.1±2.05 3.78±1.55a
група Б
(n=13)
46.13±5.20 41.12±2.0b 37.54±6.15b 27.65±3.75 22.42±3.15 17.77±3.10c
контрола
(n=8)
80.24±2.45 80.0±4.04 75.60±3.63 50.63±6.79 40.25±4.47 18.50±6.25с
Данните са x ± SD, разликата между a и b р<0.05; a и c р<0.001 във вертикалните колони
Особено значими са тези разлики при сравняване с параметрите от свежа сперма – контрола. Данните са представени на Таблица 2. Вижда се, че до 240-тата минута % на сперматозоидите с активно настъпателни движения на контролните сперматозоиди е около два пъти по-голям в сравнение с тези на група Б. Независимо от тази тенденция на 300-тата мин. няма статистически достоверна разлика между подвижните сперматозоиди от група Б (с добра криотолерантност) и контролната проба (свежа сперма). Докато тези резултати сравнени с група А (с понижена криотолерантност) са статистически значими - р<0.001.
Изследване на морфологичните изменения в структурата на половите клеткиМорфологичните промени бяха анализирани след тристепенно
оцветяване с еозин, конго рот и генциан виолет. С това оцветяване се визуализират много добре главичката и опашката на сперматозоидите. Анализът на морфологичните промени показват, че % нормални сперматозоиди спада от 79.36±9.70 за свежите еякулати до 53.16±6.60 в група Б и 49.10±5.37 в група А за криоконсервираните.
От направения сравнителен анализ се установи, че най-значими и статистически достоверно доказани различия се наблюдават в процента сперматозоиди с набъбнали акрозоми. Има проучвания, които също показват, че при кочове най-чувствителна е акрозомата на сперматозоидите (Белоус А. 1986). В повечето случаи акрозомата е набъбнала и процента достига до 33.60±3.90% за замразените сперматозоиди, докато в сравнение със свежите сперматозоиди, този процент е само 6.33±3.31% (р<0.01 – Таблица 3).

Таблица 3. Морфологичен анализ на сперматозоиди след получаване и криоконсервирани.
проба нормални сперматозоиди
набъбнали акрозоми
разкъсани акрозоми
промени в опашката
свежа (n=20) 79.36±9.70 6.33±3.31а 4.01±0.15 10.30±2.60група А (n=7) 49.10±5.37 33.60±3.90b 6.05±1.13 11.25±2.69група Б (n=13) 53.16±6.60 29.66±4.10 5.63±1.95 11.55±1.95
Данните са x ± SD, разликата между a и b р<0.01
Процентът клетки с разкъсани акрозоми както и броя на сперматозоидите с промени в тялото и опашката са без статистически достоверна разлика в отделните групи, наблюдавани клетки (Таблица 3). От проведения опит става ясно, че след замразяване най-чувствителната структура от морфологична гледна точка е акрозомата на сперматозоидите от коч. В останалите структури не се наблюдават големи изменения.
1.2. Анализ на поведението на повърхностно мембранни молекули чрез анексин V.За визуализация на молекулните промени във фосфолипидната
асиметрия на плазмената мембрана на сперматозоидите се направи анализ чрез анексин V/6CFDA.
От представените резултати се вижда, че при контролната група % живи сперматозоиди с промени в позицията на ФС в ПМ (анексин V+, живи) е сравнително нисък – 2.30±0.89, в сравнение с пробите след криоконсервация, където се наблюдава увеличаване на броя живи сперматозоиди с транслоциран ФС съответно на 31.854.53 (p<0.001) (Таблица 4). Също така, при свежите сперматозоиди прави впечатление високия процент на живи клетки (анексин V-, живи) със съхранена структура на ПМ (90.17±5.75) в сравнение със размразените клетки (54.446.16).
След криоконсервация на сперматозоидите, нараства процента на живите клетки с транслокация на ФС от ПМ и той достига до 31.854.53, повишава се и процента на анексин позитивните, мъртви клетки (13.743.82) в сравнение с контролните сперматозоиди като разликата е статистически достоверна (р<0.05).
Таблица 4. Изследвания на сперматозоиди от коч преди и след криоконсервация чрез анексин V-Cу3.18/6CFDA тест (n=9).
проби мъртвиaнексин V+/6
CF-
живи клетки с транслоциран ФСaнексин V +/6CF+
живиaнексин V-/6 CF+
контрола 6.53±2.13a 2.30±0.89а 90.17±5.75a
криоконсервирани
13.743.82b 31.854.53с 54.446.16b
Данните са x ± SD, разликата между a и b р<0.05; a и c р<0.001
На фигура 14 са показани сперматозоиди оцветени чрез анексин V теста (виж приложение 1). Интерес представлява специфичната локализация на екстернализирания ФС от ПМ. При сперматозоидите след криоконсервация транслокацията на ФС е най-вече в областта на тялото, където е разположен митохондриалния апарат – фигура 12. Тези данни са потвърждение на наши
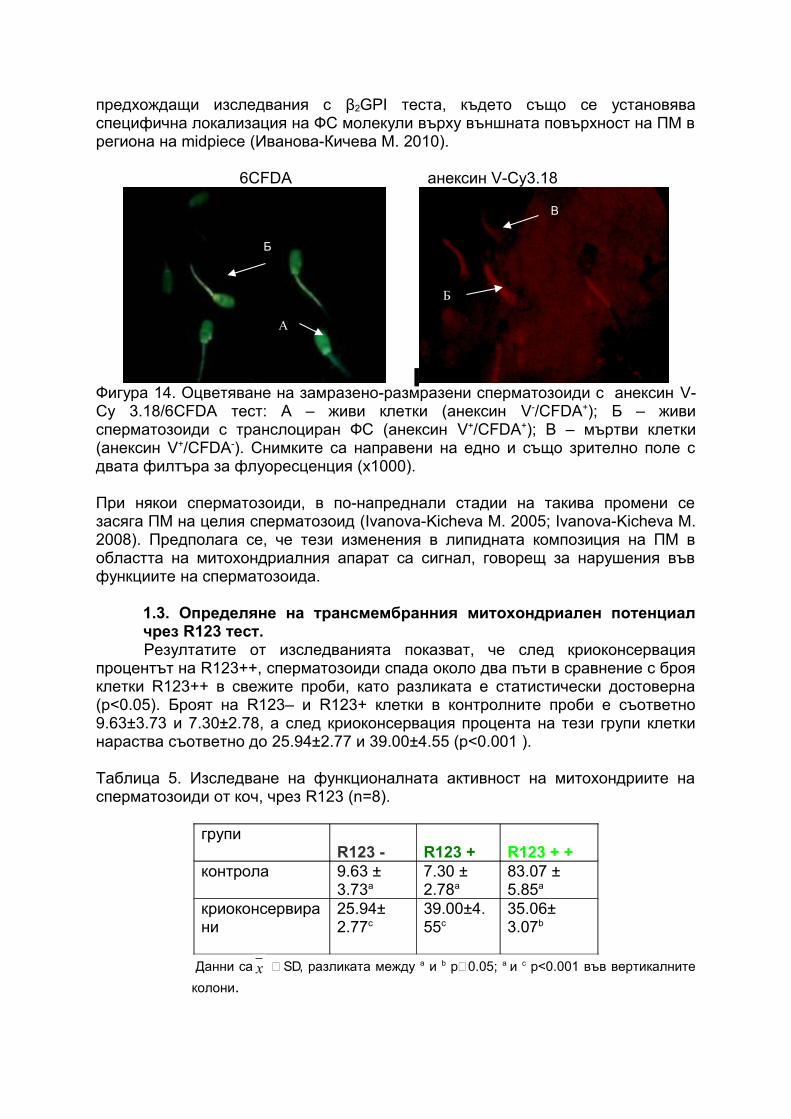
предхождащи изследвания с β2GPI теста, където също се установява специфична локализация на ФС молекули върху външната повърхност на ПМ в региона на midpiece (Иванова-Кичева М. 2010).
6CFDA анексин V-Cy3.18
Фигура 14. Оцветяване на замразено-размразени сперматозоиди с анексин V-Cу 3.18/6CFDA тест: А – живи клетки (анексин V-/CFDA+); Б – живи сперматозоиди с транслоциран ФС (анексин V+/CFDA+); В – мъртви клетки (анексин V+/CFDA-). Снимките са направени на едно и също зрително поле с двата филтъра за флуоресценция (х1000).
При някои сперматозоиди, в по-напреднали стадии на такива промени се засяга ПМ на целия сперматозоид (Ivanova-Kicheva M. 2005; Ivanova-Kicheva M. 2008). Предполага се, че тези изменения в липидната композиция на ПМ в областта на митохондриалния апарат са сигнал, говорещ за нарушения във функциите на сперматозоида.
1.3. Определяне на трансмембранния митохондриален потенциал чрез R123 тест.Резултатите от изследванията показват, че след криоконсервация
процентът на R123++, сперматозоиди спада около два пъти в сравнение с броя клетки R123++ в свежите проби, като разликата е статистически достоверна (p<0.05). Броят на R123– и R123+ клетки в контролните проби е съответно 9.63±3.73 и 7.30±2.78, а след криоконсервация процента на тези групи клетки нараства съответно до 25.94±2.77 и 39.00±4.55 (р<0.001 ).
Таблица 5. Изследване на функционалната активност на митохондриите на сперматозоиди от коч, чрез R123 (n=8).
групи R123 - R123 + R123 + +
контрола 9.63 ± 3.73а
7.30 ± 2.78а
83.07 ± 5.85a
криоконсервирани
25.94± 2.77c
39.00±4.55c
35.06± 3.07b
Данни са x SD, разликата между a и b p0.05; а и c р<0.001 във вертикалните колони.
А
Б
В
ФигуБ

От анализа на данните се установи, че след криоконсревация статистически достоверно нараства броят на родамин негативните полови клетки. Също така, процентът на R123++, сперматозоиди драстично намалява след размразяване (р<0.05). Тези данни позволяват да се каже, че ултраниските температури при замразяването на сперматозоидите от коч водят до увреждане на митохондриите, като статистически достоверно намалява броят на полови клетки с нормално функциониращи митохондрии.
1.5. Сравнителен анализ на интегритета на ДНК на сперматозоидите чрез микрогел електрофореза на единични клетки (comet assay) и АО тест.В нашите опити чрез кометен метод не можахме да установим ДНК
нарушения в сперматозоиди от коч след криоконсервация (Фигура 17, приложение 2). Предполагаме, че възможни причини за тези резултати са здраво пакетираното ядро и че в експериментите е работено с еякулати, имащи добри качествени и количествени показатели. Тези резултати са в унисон с изследванията на други автори, където се посочва изключително малко нарастване на броя клетки с увредени ядра след размразяване (Nur 2010).
Фигура 17. А. Кометен анализ на сперматозоиди от коч след криоконсервация. ДНК остава интактна без миграция извън клетката (х400).
Таблица 6. Анализ на ДНК интегритета на сперматозоиди от коч преди и след замразяване, чрез акридин оранж оцветяване (n=4).
Проби Зелена флуоресценция
Оранжева флуоресценция
контрола 99.34±0.47 0.66±0.40a
след криоконсервация
94.55±2.50 5.45±1.20b
Данните са x ± SD, разликата между a и b р<0.05; във вертикалните колони
Проведения анализ чрез акридин оранж на сперматозоиди преди и след криоконсервация също показа, че само в единични клетки се регистрира увредена ДНК. При еднакви начални параметри на еякулатите, след

криоконсервация незначително се увеличава процента сперматозоидите с увредена ДНК. Наблюдаваните клетки с оранжева флуоресценция, което говори за промени в структурата на ДНК са със степен на достоверност - р<0.05 (Таблица 6).
Задача 2. Хроматографско сепариране на протеини от спермална плазма на коч и получаване на имунен серум.За изпълнението на поставените задачи в дисертацията, СПП бяха
разделени чрез молекулно-ситова хроматография и разделителен гел - Sephacryl S-100 high resolution (Sigma, St. Louis, MO, USA).
2.1 Молекулно ситовата хроматография на спермално плазмени протеиниОт проведената хроматография са получени по 4 мл от всяка фракция
при скорост на събиране на фракциите - 4 мл/мин. На получените 15 спермално плазмени фракции е определено спектрофотометрично белтъчното съдържание. В отделните фракции се съдържаха протеини с различна молекулна маса, която варираше от 10 до 200 кДа. Концентрацията на протеините варира от 0.121 мг/мл във фракция 13 до 1.026 мг/мл във фракция 3.
2.2 Доказване и характеристика на селектираните белтъци в отделните фракции, чрез SDS-PAGE електрофореза.На всички получени фракции е направена характеристика на
селектираните чрез хроматография белтъци чрез SDS-PAGE електрофореза. За работа е използван 15% ПАА гел. Оцветяването на гела беше направено чрез метода на сребърното оцветяване. Установява се, че във фракции 2, 3, 4, 5 и 6 се наблюдават ивици на белтъци с ММ около и над 66 кДа. В някои от тези фракции се наблюдават малко количество ниско молекулни белтъци. Фракции 2, 4 и 6 съдържат белтъци с ММ по-голяма от 30 кДа. Във фракция 7 се открива един банд на около 20 кДа, а във фракция 8 има три ивици на протеини с ММ под 20 кДа. Фракции 9 и 10 съдържат белтъци с ниска ММ, под 20 кДа.Във фракция 11 се наблюдават освен ниско молекулни протеини и една ивица на белтък с ММ около 66 кДа, и една много бледа ивица на протеин с ММ около 30 кДа. Във фракция 12 се открояват две ивици, белтък с ММ около 25 кДа, а другият с ММ около 14 кДа. Във фракции 13, 14 и 15 се открояват две ивици на белтъци с ММ около 20 кДа и на такива с ММ около 14 кДа.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
66 кДа
29 кДа
20 кДа
14.2 кДa
200 кДа
36 кДа

Фигура 18. ПАА електрофореза на белтъците съдържащи се във фракциите получени след молекулно ситова хроматография. 1 – маркер за молекулни тегла; от 2 до 15 – фракции от 2 до 15.
2.3 Разделяне, идентифициране и определяне на белтъчното съдържание в някои фракции от сепарирани протеини на спермална плазма, чрез течна хроматография (HPLC).Резултатите от настоящите изследвания позволяват да се твърди, че
във фракция 15 преобладават ниско молекулни белтъци с ММ под 30 кДа, отделени в два пика. Фракция 11 се състои от четири ниско молекулни белтъка с ММ под 30 кДа и един протеин с ММ около 66 кДа.
Фигура 19. Хроматограми на: А) маркер за молекулни тегла; Б) проба 15; В) проба 11
2.4. Получаване, пречистване и характеристика на имунен серум срещу спермално плазмени протеини от коч.Получаването на имунен серум срещу протеини от спермална плазма се
наложи, поради необходимостта за биологичен и цитохимичен анализ на селектираните от нас СПП чрез хроматография. За целта е работено със опитни зайци. Резултатите след имунизиране на лабораторните животни показаха, че е получен имунен заешки серум срещу спермално плазмени белтъци от коч.
0
0,5
1
1,5
2
2,5
3
3,5
1/100
0
1\200
0
1\400
0
1\800
0
1\160
00
1\320
00
1\640
00
1\128
000
256000
Е
имунен серум преимунен серум
Фигура 20. Определяне титъра на имунният заешки серум срещу спермално плазмени протеини от коч, при падащи разреждания на антитялото. За контрола на реакцията е използван преимунен заешки серум.

0
0,5
1
1,5
2
2,5
3
3,5
1/1000 2000 4000 8000 16000 32000 64000
Е
фракция 3 фракция 14 фракция 15
фракция 6 фракция 4 фракция 11
Фигура 21. Реакция на имунния серум с отделните фракции съдържащи различни СПП.
От направения анализ чрез ELISA, с падащи разреждания на антитялото се установи, че титъра на полученият имунен серум е около 1:16000 (Фигура 20). Допълнително е направен тест на антитялото относно реакцията му със сепарираните от нас чрез хроматография протеинови фракции. От изследваните проби най-силна беше реакцията на имунния серум срещу белтъците съдържащи се във фракция 3. От направената SDS PAGE се установи, че това са високо молекулни белтъци, чието съдържание в СП е представени в по-голямо количество, в сравнение с другите протеини. Забелязва се, че реакцията на имунния серум към фракциите, които съдържат ниско молекулни белтъци е по-слабо проявена (фигура 21).
Задача 3. Анализ на ролята на селектираните протеини върху функционалността на сперматозоидите от коч при условия на индуциране на температурен шок.Изследването на подвижността и преживяемостта на сперматозоидите
при съхраняване на 50С след инкубиране с отделните фракционирани СПП ще ни даде информация за влиянието на тези белтъци върху половите клетки.
3.2 Влияние на селектираните протеини върху скоростните параметри на сперматозоиди от коч след индуциране на температурен шок и след инкубация на 50С за 24, определени чрез компютърен спермоанализатор.Анализ на неподвижни (статични), прогресивно подвижни и не-прогресивно подвижи сперматозоиди
За настоящите изследвания са използвани СПП от фракции 3, 4, 6, 8, 11, 15 и две контроли - К+ е контрола с цяла СП, а К- е контрола без СП. За експеримента е работено с еякулати от кочове порода Мутон-Шароле (Mouton Sharole) и Романовски (Romanov Breed). Спермалните параметри са следните: цвят от бял до кремав; pH 6.8-7; без мирис; без аглутинация; концентрация – 3.3-4х109 кл/мл. На Таблици 11 и 12 са представени данните за прогресивната подвижност на сперматозоидите, инкубирани със сепарираните СПП след температурен шок и след 24h инкубиране на 50С.

Таблица 11. Подвижност на сперматозоидите (в %) инкубирани със СПП след температурен шок (n=5).
проби статичнине прогресивно подвижни
прогресивно подвижни
К+ 42.21±20.83 41.50±8.60 16.29±7.62К- 60.28±15.63a 26.79±12.38 12.93±4.62проба 3 66.86±14.27a 28.48±8.46 4.66±1.56a
проба 4 52.68±9.05 35.98±8.73 11.34±8.73проба 6 50.10±15.81 46.20±9.96 3.70±1.75a
проба 8 33.74±10.29 50.52±12.93 15.74±9.67проба 11 27.22±16.95b 48.14±17.53 24.64±6.39c
проба 15 30.97±11.64b 44.21±16.23 24.82±4.32c
Данните са x ± SD, разликите между а и b p<0.05; a и c p<0.001 във вертикалните колони.
Резултатите от изследванията показват, че процентът на статичните сперматозоиди е най-висок при проби К-, 3 и 4, а най-нисък съответно при проби 8, 11 и 15. Като разликата между проба 3 и 11 е статистически достоверна (p<0.05). Процентът на прогресивно подвижните сперматозоиди е най-висок в проби 11 и 15 (респективно 24.64±6,39 и 24.82±4,32) и е близък до процента прогресивно подвижни сперматозоиди в свежите проби. Значително по-нисък процента на прогресивно подвижните сперматозоиди бе наблюдаван в проби 3 и 6 съответно 4.66±1.56 и 3.70±1.75 (p<0.001).
От направения анализ се установи, че в сравнение с началните данни след ТШ, рязко се увеличава процента на статичните клетки, като в проба 3 това увеличение е около десет пъти. Индуцирането на ТШ води до намаляване на процента не-прогресивно подвижни клетки, а при проби 3 и 6 се вижда рязко намаляване на броя на прогресивно подвижните сперматозоиди. Тези резултати ни насочват към предположението, че във фракции – 3 и 6, не се съдържат протеини с протективен ефект.
Таблица 12. Подвижност на сперматозоидите (в %) след 24h инкубиране на 50С (n=5).
проби статичнине прогресивно подвижни
прогресивно подвижни
К+ 44.86±14.4 51.43±16.31 3.71±1.09К- 70.25±9.60a 28.48±8.77 1.27±0.94проба 3 69.90±12.60a 29.8±12.54 0.30±0.07a
проба 4 66.80±6.26 31.83±6.87 1.37±0.89a
проба 6 63.25±4.15 36.50±4.15 0.25±0.05a
проба 8 59.6±10.15 34.30±9.68 6.10±0.64проба 11 41.25±13.42b 51.10±13.99 7.65±2.29с
проба 15 56.99±6.46 40.48±7.41 2.53±1.39 Данните са x ± SD; разликите между а и b p<0.05; а и с p<0.01 във вертикалните колони
За нас особено интерeсен резултат е, че при проби 11 и 15 броят на прогресивно подвижните клетки след ТШ се запазва и е много близък до стойностите в началото на експеримента. Тези резултати позволяват да се

изкаже, че селектираните протеините, съдържащи се в тези две проби имат протективен ефект върху мотилитета на сперматозоидите и това се доказва с добре съхранената прогресивна подвижност, която остава почти непроменена преди и след ТШ.
Резултатите от прогресивното движение на сперматозоидите след 24 часа съхраняване на 50С са представени на Таблица 12. Процентът на неподвижните (статичните) полови клетки беше висок в проби К-, 3, 4 и 6, докато в проби К+ и 11 този процент е по-нисък. Тези резултати показват, че спермалната плазма като цяло, има протективен ефект и доставя нужната енергия на сперматозоидите при продължително съхраняване. От друга страна, най-висок брой прогресивно подвижни сперматозоиди бяха отчетени в проби 8 и 11 (респективно 6.10±0.64 и 7.65±2.29), най-нисък процент прогресивно подвижни сперматозоиди се откри в проби К-, 3, 4 и 6 съответно 1.27±0.94, 0.30±0.07, 1.37±0.89 и 0.25±0.05 процента. Разликите между проба 11 и проби 3 и 6 са статистически достоверни (р<0.01).
Скорост на движение – бързи и средно подвижни сперматозоидиДруг изследван параметър беше скоростта на движението на
сперматозоидите. Анализът е направен върху бързи и средно подвижни сперматозоиди – Таблици 15 и 16. Установява се, че процентът бързи клетки е най-висок в пробите К+, 11 и 15 (21.27±5.89, 33.23±16.47 и 31.90±17.13), а най-нисък в проби К-, 3 и 6, съответно 3.13±1.23, 1.90±1.00 и 4.60±0.90. Разликите са статистически достоверни (p<0.01). Вижда се, че в проби 11 и 15 се запазва висок % на бързо подвижни сперматозоиди в сравнение с проби К-, 4, 6 и особено с проба 3. Този анализ отново потвърждава, че нискомолекулните протеини от фракция 11 и 15, както и наличието на цяла СП, протектират гаметите. Отсъствието на СП или наличие на протеини с големи молекулни тегла – фракции 3 и 6 води до драстично намалява на процента бързоподвижни сперматозоиди.
Таблица 15. Скорост на движение на сперматозоидите след температурен шок. Данните показват процента на бързо подвижните сперматозоиди (n=5).
Данните са x ± SD; разликите между а и b p<0.05; a и c p <0.01, във вертикалнните колони
проби бързи средно подвижни
К+ 21.27±5.89c 9.23±4.20К- 3.13±1.23a 5.68±3.92проба 3 1.90±1.00a 5.23±2.60проба 4 9.48±7.60 5.72±2.79проба 6 4.60±0.90 2.02±0.48проба 8 8.34±4.84b 15.00±8.54проба 11 33.23±16.47c 11.83±4.47проба 15 31.90±17.13c 12.98±3.18

Таблица 16. Скорост на движение на сперматозоидите след 24 часа инкубиране на 50С. Данните показват процента на бързо подвижните сперматозоиди (n=5).
Данните са x ± SD
След продължително съхраняване на ниска температура при проба 11 се установи най-висок брой на бързите клетки, съответно 4.44±0.78. Въпреки това дълговременното съхраняване понижава стойностите на този показател и се вижда, че в сравнение с началните параметри (Таблица 9) намалението на процента бързи клетки е около осем пъти. При сравнение на тези резултати между пробите съхранявани за 24 часа може да се каже, че в проби K+ и 11 се запазва броят на бързо подвижните клетки около 4% за разлика от проби К-, 3, 4 и 6, където % бързи клетки е от 0.3 до 0.75 – Таблица 16. Тези резултати показват, че съхраняването на сперматозоидите на ниски температури води до значително намаляване на процента бързо подвижни клетки. Това намаляване се наблюдава, както при сперматозоиди с индуциран температурен шок, така и при тези, които са съхранени на 50С за 24 часа. Визуализирането на резултатите от Таблица 15 и 16 са представени на Фигура 24 А и Б.
скорост
0
5
10
15
20
25
30
35
К+ К- проба3
проба4
проба6
проба8
проба11
проба15
%
бързи бавни
скорост
0
2
4
6
8
10
12
К+ К- проба3
проба4
проба6
проба8
проба11
проба15
%
бързи бавни
А) Б)
Фигура 24. А) Данни за скоростта на сперматозоидите след ТШ, а на фигура Б) Резултати за скоростта на половите клетки след 24 часа инкубиране на 50С.
Скоростни параметри на сперматозоидите - VCL, VSL, VAPИзследването на скоростните параметри дава допълнителна
информация за състоянието на сперматозоидите. VCL показва скоростта на сперматозоида при изминаването на действителния път за единица време (сек.). VSL показва праволинейната скорост на сперматозоида, Измерва се изминатото разстояние по права линия от началото до края на движението на клетката и се разделя на съответното време. VAP е скоростта на средния
пробибързи средно
подвижниК+ 1.95±1.3 5.2±2.20К- 0.75±0.07 1.10±0.35проба 3 0.30±0.20 1.16±0.75проба 4 0.63±0.37 3.53±0.58проба 6 0.30±0.10 1.70±0.56проба 8 1.47±0.12 5.10±1.15проба 11 4.44±0.78 10.05±2.92проба 15 1.89±0.55 5.98±1.02

изминат път. Тези данни ни показват каква е скоростта на движение на мъжките полови клетки.
Скоростни параметри на сперматозоидите след ТШЗа сравнение на резултатите, същите скоростни параметри бяха
изследвани след ТШ при инкубиране на пробите сперматозоиди с отделните фракции СПП. Резултатите са показани на таблица 18 и 19. Установява се, че скоростта на сперматозоидите при изминаването на действителни път (VCL) е най-висока в проби К+, 11 и 15 (67.90±26.60, 67.24±16.60 и 62.01±17.81) в сравнение с проби 3, 4 и 6 като разликата е статистически достоверна при сравняване на проби 3 и 11 (р<0.05) (виж таблица 18). Подобно е положението при скоростните параметри VSL и VAP, като най-ниски са данните при проби 3, 4 и 6, а най-високи при проби К+, 11 и 15.
Таблица 18. Данни за скоростта след ТШ - VCL µm/sec, VSL µm/sec и VAP µm/sec (n=5).
проби VCL µm/sec VSL µm/sec VAP µm/secК+ 67.90±26.60 37.29±13.78 49.04±19.92К- 42.89±16.14 21.63±11.5 28.47±16.75проба 3 29.04±11.56b 10.53±3.65b 15.33±3.79b
проба 4 39.40±14.54 21.46±15.93 27.32±16.85проба 6 36.13±5.11 14.85±6.07b 24.50±12.59проба 8 49.33±13.55 28.93±14.25 37.23±15.32проба 11 67.24±16.60a 41.21±10.40a 53.22±13.45a
проба 15 62.01±17.81 38.21±8.78a 42.66±10.33 Данните са x ± SD; разликите между а и b p<0.05, във вертикалните колони.
Скоростни параметри на сперматозоидите след инкубиране на 50С за 24 часа.Скоростните параметри на сперматозоидите бяха измерени след 24
часово инкубиране на 50С. Данните показаха, че при всички проби стойностите на измерените параметри (VCL, VSL и VAP) намаляват, в сравнение с тези при свежи проби и след ТШ. Стойностите на VCL са по-високи в проби К+, 8, 11 и 15, за разлика от проби 3 и 6. Като при сравнение между проби 3 и 11 се наблюдава статистически достоверна разлика (p<0.05). Данните за VSL са по-високи в проби К+ и К-, при останалите проби стойностите са много близки и няма статистически достоверна разлика. VAP показателя беше най-висок при проби К+ и 11. Получените резултати показват, че продължителното инкубиране на сперматозоидите на ниски температури оказва пагубно влияние върху мотилитета на гаметите и скоростта им на придвижване. Все пак, в пробите инкубирани с ниско молекулни СПП се наблюдава тенденция към запазване на по-добри скоростни параметри (VCL, VSL, VAP).

Таблица 20. Данни за скоростта след 24 часа инкубиране на сперматозоидите на 50С. VCL µm/sec, VSL µm/sec и VAP µm/sec (n=5).
проби VCL µm/sec VSL µm/sec VAP µm/secК+ 30.45±5.77a 15.38±4.87 21.18±5.71К- 23.70±6.50 10.93±2.71 13.42±6.17проба 3 20.5±2.68b 8.40±3.40 12.23±2.95проба 4 24.63±0.72 9.93±2.60 11.20±4.05проба 6 22.60±1.62 5.93±2.70 10.33±2.50проба 8 27.23±5.75 7.33±3.15 13.53±3.82проба 11 30.06±6.32a 9.34±5.01 15.24±4.85проба 15 25.73±4.95 7.47±2.70 10.73±3.53
Данните са x ± SD; разликите между а и b p<0.05;
Задача 4. Сравнителен анализ върху ролята на селектирани спермално плазмени протеини върху целостта на плазмената мембрана чрез 6CFDA/PI тест и флоуцитометър във връзка с жизнеността на половите клетки след температурен шок и след инкубация на 50С за 24.Анализът по тази задача е направен с оглед изследване структурната
цялост на ПМ и жизнеспособността на половите клетки, под влияние на селектираните чрез хроматография протеини на СП. Редица данни в литературата показват, че ПМ на сперматозоида е много деликатна и важна клетъчна структура за живота на мъжките полови клетки.
4.2 Изследване на мембранния интегритет и преживяемостта на сперматозоидите след температурен шок и при инкубация на 50С за 24 часа.Определянето на мембранния интегритет след температурен шок е
направен чрез флуоресцентните маркери CFDA/PI – Фигура 27, приложение 3. Резултатите показаха, че от 10те изследвани проби в К+, К-, 8, 14 и 15 се наблюдава най-висок процент на CFDA оцветени сперматозоиди. Сперматозоидите инкубирани с протеини от тези фракции показват по-голям брой CFDA положителни клетки в сравнение с пробите инкубирани с фракции 3, 4, 6, 10 и 11 като разликата е статистически достоверна само при сравняване на проби 3 и 15 (р<0.05).
Изследването на преживяемостта на сперматозоидите след 24 часа инкубация на 50С показа, че CFDA оцветените клетки в проби К+, 14 и 15 са повече на брой в сравнение с проби 4, 10. Сперматозоидите инкубирани в пробите с нискомолекулни протеини или цяла СП, показват достоверно по-високо съхраняване на скоростните параметри след индуциране на ТШ, в сравнение с останалите проби. След съхраняване за 24 часа прави впечатление рязкото увеличаване на броя на PI позитивните сперматозоиди във всички проби.

0
10
20
30
40
50
60
70
80
90
100
ТШ 24h ТШ 24h ТШ 24h ТШ 24h ТШ 24h ТШ 24h ТШ 24h ТШ 24h ТШ 24h ТШ 24h
K+ K- 3 4 6 8 10 11 14 15
проби
%
cfda+/PI- cfda-/PI+ cfda+/PI- cfda-/PI+
Фигура 25. Оцветяване с CFDA/PI на сперматозоиди от коч веднага след температурен шок и след 24 часа инкубиране на 50С.
Резултатите от изследването показват, че след ТШ пробите, в които присъстват нискомолекулни СПП (14 и 15) предпазват по-добре ПМ на сперматозоидите, в сравнение с пробите, в които се намират високомолекулни СПП (3, 4 и 6). Оттук и процентът на жизнеспособните полови клетки е по-висок в проби 14 и 15.
Фигура 27. Оцветяване на сперматозоиди с CFDA (ляво) и PI (дясно) (х400 – дигитално увеличен размер на клетките).
4.3 Изследване на морфологичните изменения в структурата на половите клетки след съхраняване на 50С.Изследванията са направени чрез метод за тристепенно оцветяване на
сперматозоидите с боите – еозин, конгорот и генцианвиолет (Nikolov I. 1991). Анализът на морфологичните промени показват, че % нормални сперматозоиди в проби К+, 11, 14 и 15 е най-висок, съответно 81.91±10.56, 79.33±15.63, 82.42±10.84, 81.80±8.15.00, а в проби 4 и 6 е най-малък, съответно 56.24±12.05 и 51.01±15.26. Разликите са статистически значими (Таблица 22). Наблюдава се, че няма статистически достоверни разлики в отделни проби при процента разкъсани акрозоми. При следващият изследван параметър, обаче

се вижда, че в проби 4 и 6 се установява висок процент на клетки с набъбнали акрозоми в сравнение с останалите проби. Като има статистически значими разлики с К+, проби 14 и 15 (p<0.01). Предполагаме, че промените на морфологичните параметри се дължат на протеините, които се съдържат в средата за съхранение и оказват своето влияние върху сперматозоидите. Възможно е тези спермално плазмени белтъци да стимулират капацитацията и да правят ПМ по-рехава. Също така инкубирането на сперматозоидите на ниска температура води до появата на т. нар. криокапацитация (Morrier A. 2002), която се изразява с дестабилизиране на ПМ. Най-вероятно влиянието на ниските температури и отделните СПП водят до промяна в структурата на плазмалемата и акрозомните мембрани, което e възможно да се проявява като набъбване на акрозомите на сперматозоидите инкубирани в проби 4 и 6. Промените в опашката и нейните сегметни се откриват при всички проби са в близки граници при всички проби. Разликите между отделните проби не са статистически достоверни.
Таблица 22. Данни от ролята на СПП върху морфологичните промени на сперматозоиди съхранявани на 50С (n=6).
нормални разкъсана акрозома
набъбнала акрозома
промени в опашката
K+ 81.91±10.56 b 1.75±0.80 1.75±0.56 с 14.56±3.65K- 72.56±6.65 2.73±0.35 4.73±1.25 19.98±4.903 69.00±8.65 1.50±0.15 2.50±0.95 27.00±7.504 56.24±12.05 a 1.00±0.45 18.98±2.65a 23.78±4.506 51.01±15.26a 1.50±0.25 19.13±5.56a 28.37±5.658 79.02±10.56 0.50±0.15 1.50±0.25 19.00±7.5610 73.03±12.30 0.50±0.10 3.01±0.95 23.50±4.5011 79.33±15.63 1.01±0.35 3.27±1.25 16.40±3.5614 82.42±10.84 b 1.02±0.25 1.42±0.65 с 15.16±4.2515 81.80±8.15.00b 0.75±0.15 1.95±0.42с 15.50±5.56
Данните са x ± SD, като разликите между а и b p<0.05; разликата a и с р<0.01.
При провеждането на опитите се установи, че след инкубиране на сперматозоидите със СПП от фракция 3 се наблюдаваше аглутинация на половите клетки, съпроводено със струпване на група клетки на едно място, които са захванати една с друга в областта на главичката - Фигура 25. Тези резултати представляваха интерес за нас, тъй като в литературата не намерихме подобни резултати. Анализирайки резултатите до сега и получените данни от нас ние си позволяваме да изкажем хипотезата, че във фракция 3 се съдържат СПП, които повлияват върху електрическия заряд на повърхността на половите клетки, което от своя страна води до поява на аглутинация и слепване на сперматозоидите по между им. По този начин сперматозоидите придобиват силно изразени адхизиращи качества и полепват един към друг, а в също време губят своя мотилитет. На тази проба решихме да направим имуноцитохимично изследване, за да проверим дали някои от СПП се локализират по повърхността на сперматозоидите и взаимодействайки с ПМ причиняват наблюдаваната аглутинация. След имуноцитохимично изследване с полученият от нас имунен серум се установи, че там където са аглутиниралите сперматозоиди се откриват и СПП. Считаме, че това са белтъците взаимодействали с клетъчната повърхност на сперматозоидите,

което води до тяхното аглутиниране - Фигура 26. Резултатите от този анализ доказват, че някои от СПП се прикрепят върху плазмената мембрана, като по този начин моделират последващото поведение на сперматозоидите.
Фигура 25. На фигурата е показана снимка на сперматозоиди от проба 3 инкубирани със СПП от фракция 3, ясно се вижда аглутинацията (бялата стрелка) и слепването на клетките (обектив 10х, SCA, Microptic, Spain).
4.4 Изследване на целостта на ПМ (виталитета) на сперматозоидите чрез флоуцитометър.
За прецизиране на изследванията са направени анализи върху интегритета на ПМ чрез флоуцитометър. Резултатите показаха, че след 24 часа инкубиране на 50С, във всички изследвани проби се наблюдава значително нарастване на двойно оцветените сперматозоиди. Това са сперматозоиди с промени в ПМ в различна степен. Най-висок е процента на CFDA+/PI+ сперматозоиди в проба 3, а най-нисък в К- и проба 15, съответно 58.66±0.33 срещу 50.39±2.17 и 52.77±1.12 - Таблица 23. Тези данни потвърждават нашите изследванията по подзадача 4.1, 4.2 и 4.3.
Таблица 23. На таблицата са представени процентите на CFDA/PI оцветените сперматозоиди от коч след 24 часа инкубиране на 5oС
проби(n=3)
CFDA+/PI- CFDA-/PI+ CFDA+/PI+
K+ 15.73±1.37 7.42±0.38 54.20±2.31K- 11.90±1.18 16.59±2.46 50.39±2.17проба 3 11.47±1.48b 17.20±0.21c 58.66±0.33проба 15 17.27±0.55a 7.25±0.25a 52.77±1.12
Данните са x ± SD; разликите между а и b p<0.05; a и c p<0.001, във вертикалните колони
Установява се, че при продължително съхраняване на сперматозоидите в охладено състояние се наблюдава нарушаване на целостта на ПМ, която става пропусклива за пропидивия йодид. Броят на PI+, мъртви полови клетки е най-висок в К- и проба 3 съответно 16.59±0.46 и 17.20±0.21. За разлика от тях в К+ и проба 15 бяха отчетени най-ниските проценти на PI позитивните клетки. Като разликата между проби 3 (17.20±0.21) и 15 (7.25±0.25) е статистически значима (р<0.001). Все пак изследванията показаха, че процентът на CFDA+, живи сперматозоиди е най-висок в проба 15, а най-нисък в проба 3, съответно

17.27±0.55 и 11.47±1.48, като разликата е статистически достоверна (p<0.05) – Таблица 23. Това означава, че в проба 15 има най-голям брой сперматозоиди с добре запазена интактна плазмена мембрана, в сравнение с другите проби. Може да се твърди, че в проба 15 и К+ се запазват най-много жизнени сперматозоиди след 24 часа съхраняване на 50С.
Задача 5. Анализ върху поведението на вътреклетъчни структури – митохондрии и ДНК при нискотемпературно съхранение на 50С на сперматозоиди от коч, в присъствие на селектирани спермално плазмени протеини. Определянето функционалността на сперматозоидите е свързано с
поведението на вътреклетъчните структури – митохондрии и ДНК. Съществуват данни в литературата, които показват, че митохондриите и ядрото са изключително важни клетъчни органели за живота на сперматозоида. Също така проучването на митохондриите и ядрото може да се използва като показател показващ биологичната пълноценност на сперматозоида.
5.1. Влияние на спермално плазмени протеини върху трансмембрания митохондриален потенциал на половите клетки при нискотемпературно съхранение на 50С.Анализът е проведен чрез определяне на функционалната активност на
митохондриите с R123 тест. За провеждането на експериментите сме инкубирали сперматозоидите със селектирани СПП от проби 3, 6 и 11. Опитът има две контроли, К- (без спермална плазма) и К+ (с цяла спермална плазма). Резултатите от проведените изследвания показаха, че митохондриите на сперматозоидите от коч са много чувствителни към влиянието на ниските температури. Прави впечатление, че веднага след ТШ, процентът на R123++ сперматозоиди се понижава във всички изследвани проби –Фигура 28.
0
10
20
30
40
50
60
70
К+ К- проба 3 6 11
%
R123++
R123+
R123-
Фигура 28. Състояние на митохондриите в проби К+, К-, 3, 6 и 11, след ТШ, изследване с R123. R123+, сперматозоиди, в които има известни нарушения в митохондриите; R123++ нормални сперматозоиди с функционално съхранени митохондрии; R123- сперматозоиди с нефункциониращи митохондрии.
От анализа на данните се установи, че най-висок брой сперматозоиди с нормално функциониращи митохондрии (R123++) има в проба 11, а най-нисък в проба 3, съответно 35.72±8.56 и 13.39±6.12%, разликата е статистически
R123-

достоверна (р<0.05). Прави впечатление, че след охлаждане процентът на сперматозоидите с интактни митохондрии е силно занижен като най-многобройни и преобладаващи в почти всички проби са R123+ оцветените сперматозоиди. От друга страна, в сравнение със свежите проби, където процента на R123 негативните сперматозоиди е 9.63±3.73, след ТШ се наблюдава повишаване на броя R123 негативни клетки в пробите, достигащ до 30% при проба К+. Може да се каже, че в пробите след температурен шок се наблюдава драстично намаляване на броя на R123++ сперматозоиди, т.ест. намаляват клетките с нормално функциониращи митохондрии, а значително се увеличава процента на сперматозоидите с начални изменения в митохондриите (R123+).
5.2 Влияние на спермално плазмени протеини върху интегритета на ДНК на половите клетки при нискотемпературно съхранение на 50С.Анализът на ДНК интегритета е направен при сперматозоиди от коч,
съхранявани на 50С за 24 часа в присъствието на СПП чрез АО тест (ТАблица 25). Преди провеждането на опитите инкубирахме сперматозоидите за минимум 24 часа на 50С. Това се направи с цел да се предостави достатъчно време на факторите от средата и клетъчните процеси за действие и проява на деструктивните изменения. Изследването се проведе върху еякулати от коч след отстраняване на СП, към които са добавяни СПП от фракции 3 и 11. Избраните проби показаха най-големи различия относно биологичните параметри на сперматозоидите, подвижност и преживяемост. Данните показват, че след охлаждане се наблюдава леко повишаване на процента сперматозоиди с увредена ДНК в сравнение с контролата (Фигура 31, приложение 4).
Таблица 25. Изследване на ДНК интегритета, чрез акридин оранж тест при сперматозоиди от коч съхранявани 24 часа на 50С (n=3).
проби зелена флуоресценция
оранжева флуоресценция
контрола 99.34±0.47a 0.66±0.3a
проба 3 95.85±1.9b 4.15±0.87b
проба 11 96.4±1.00 3.6±0.99b
Данните са x ± SD; разликите между а и b p<0.05, във вертикалните колони

А) Б)
Фигура 31. Изследване на ДНК интегритета чрез акридин оранж тест при: А) свежи сперматозоиди от коч; Б) сперматозоиди след нискотемпературно съхранение. Зелено оцветените ядра представляват клетките с нормално ДНК съдържимо, а оранжево оцветените ядра – увредена ДНК. (Olympus, филтър FITC. х200)
Задача 6. Изкуствено осеменяване на овце с поликомпонентна среда обогатена със селектирани спермално плазмени протеини с доказан протективен ефект.От проведените опити по съответните задачи на настоящата дисертация
до тук видяхме какво е влиянието на отделните СПП при инкубиране на сперматозоиди от коч на ниска температура. Резултатите показаха, че при пробите съдържащи ниско молекулни СПП се отчитат по-добра подвижност, преживяемост и състояние на митохондриите в сравнение с другите проби.
Таблица 26. Използване на селектирани СПП в протективните среди при ИО на овце.
фракции брой овце
повторки(брой)
повторки(%)
ранни аборти(брой)
ранни аборти
(%)
заплодяемост(%)
проба 4 20 6 30 5 25 45проба 6 20 16 80 3 15 5проба 15 20 4 20 1 5 75контрола 20 5 25 1 5 70
Получените данни ни дадоха основание да проведем биологичен опит, с който да проверим оплодителната способност на охладените сперматозоиди от коч съхранявани в присъствието на ниско молекулни СПП. Получените резултати след провеждането на опитите показаха, че при използване на ниско молекулните спермално плазмени протеини се постига по-добра заплодяемост в сравнение с пробите съдържащи високомолекулни СПП (Таблица 26).
В настоящата дисертация си поставихме за цел да проучим биологичните ефекти на ниските и ултраниските температури върху сперматозоиди от коч, протективното действие на някои спермално плазмени фактори, с оглед подобряване на настоящите и създаване на нови репродуктивни биотехнологии, необходими за контролираното размножаване в овцевъдството.
green
orange

В първата задача от изследванията беше проучен ефекта на ултраниските температури върху мотилитета и преживяемостта на сперматозоиди от коч след криоконсервация. Замразяването на половите клетки причинява ултраструктурни, биохимични и функционални промени като най-често се засягат мотилитетът и ПМ на сперматозоидите (Bailey JL. 2000). Също така, преживяемостта, способността на сперматозоидите да преминат цервикса и оплодителния потенциал на размразените полови клетки биват променени (Holt WV. 2000a; Holt WV. 2000b; Dominguez M.P. 2008). Такива изменения в гаметите обикновено водят до ниски резултати при ИО на овцете. За изясняване на причините, обуславящи ниската криоустойчивост на сперматозоидите от коч е необходимо да се проучи влиянието на ниските и ултраниските температури върху отделните компоненти, лабилни към тяхното въздействие. Установява се, че след криоконсервация значително се редуцира броят на подвижните сперматозоиди. Това означава, че ниските и ултраниските температури по време на процеса на замразяване са довели до необратими промени в половите клетки, които в крайна сметка водят до понижаване на мотилитета на сперматозоидите. Процесът криоконсервация уврежда митохондриите, акрозомата и опашката на половите клетки (Wooley and Richardson. 1978). Мотилитетът на сперматозоидите е особено чувствителен към такива увреждания (Holt WV. 2000a). Нашето предположение е, че вероятно съвкупността от въздействията на всички тези фактори води до увреждания в ключови клетъчни структури отговорни за движението на сперматозоидите. В резултат на това след криоконсревация се наблюдава драстично редуциране на броя на подвижните сперматозоид. Това води до намаляване на оплодителния потенциал на размразената сперма. Сперматозоидите от коч са особено чувствителни на замразяване, като оплодителната им способност след размразяване е минимална. Използването на такива сперматозоиди в практиката е все още ограничено. Именно това налага търсенето на нови подходи за замразяване и съхраняване на половите клетки от коч. Съществуващите данни в литературата за възможен протективен ефект на някои спермално плазмени фактори ни накара да проверим тази теория, като изследваме влиянието на отделни фракции СПП върху сперматозоидите. В настоящата дисертация се разглежда влиянието на СПП върху половите клетки съхранявани на ниска температура (50С).
Все още не е изяснено как точно се осъществява протекцията на половите клетки чрез използването на спермално плазмени фактори. Може да се предполага, че някои от ниско молекулните спермално плазмени протеини принадлежат към т. нар. спермадхезини, които могат да се свързват с глюкозоаминогликани и фосфолипиди по повърхността на сперматозоидите. Установено е, че това тяхно взаимодействие стабилизира ПМ и предпазва половите клетки от преждевременна капацитация и акрозомна реакция. Други изследвания показват, че някои от ниско молекулните СПП с протективен ефект инхибират протеинтирозин фосфорилирането на белтъци от ПМ на сперматозоидите. Това фосфорилиране е характерно за капацитацията, а при охлаждане се наблюдава капацитация-подобно явление с характерно протеинтирозин фосфорилиране на определени белтъци. Предполага се, че инхибирането на фосфорилирането от СПП би довело да стабилизиране на ПМ по време на охлаждането. Именно стабилизирането на ПМ е от изключително важно значение за жизнеността на сперматозоидите съхранявани на ниски температури. Установяването на нови субстанции,

които да протектират ПМ на сперматозоидите от коч може да има голямо приложение в практиката. Получените от нас резултати показват, че ниско молекулните спермално плазмени протеини протектират достоверно по-добре подвижността и жизнеността на сперматозоидите от коч при съхраняване на 50С. Селектираните спермално плазмени белтъци може да се използват като компоненти в средите за съхранение на сперматозоиди от коч при ИО на овце.

VII. ИЗВОДИ И ПРИНОСИИЗВОДИ1. Плазмената мембрана на сперматозоидите от коч е слабо
криоустойчива и процесът криоконсервация предизвиква: - в голям процент от сперматозоидите нарушения в региона на акрозомата (до 33.6±3.90 набъбнали акрозоми в еякулатите със снижена криотолерантност);- повишена транслокация на фосфатидилсерина от плазмената мембрана в региона на mid-piece до 31.854.53.
2. След криоконсервация значително намалява процента на клетки с нормално функциониращи митохондрии (35.06%, R123++) и се увеличава броят на сперматозоидите с неактивни митохондрии (39.00%, R123+) в сравнение с контролите (83.07%, R123++ и 7.30%, R123+ клетки). Ядрото на сперматозоидите от коч е устойчива структура на ниски и ултраниски температури.
3. Чрез хроматографско сепариране са получени 15 фракции, съдържащи спермално плазмени протеини. В три от фракциите присъстват ниско молекулни протеини с протективен ефект при in vitrо съхранение на сперматозоиди от коч в охладено състояние.
4. Сперматозоидите от коч, инкубирани в среда със селектирани ниско молекулни спермално плазмени протеини (под 30 кДа), запазват достоверно по-добра прогресивна подвижност, скоростни параметри (VCL, VSL и VAP) след температурен шок и след съхраняване на 50С за 24 часа.
5. Инкубирането на сперматозоиди от коч с поликомпонентна среда с ниско молекулни спермално плазмени протеини, съхранява по-голям брой клетки с интактна плазмената мембрана и нормално функциониращи митохондрии (разликата с пробите с високомолекулни протеини е статистически достоверна, р<0.05).
6. Адсорбцията на високомолекулни протеини (фракции 3, 4 и 6) върху определени региони на плазмената мембрана индуцира аглутинация на сперматозоидите и формиране на неправилни образувания, както и набъбване на акрозомите (19.13±5.56 - фракция 6 и 1.95±0.42, за фракция 15, р<0.001).
7. Протекцията на плазмената мембрана, вътреклетъчните структури (митохондрии и ДНК) и съхраняването на виталитета на сперматозоидите се осъществява чрез екстрацелуларно въздействие на биокомпонентите - селектирани спермално плазмените протеини.
8. Изкуствено осеменяване на овце с дълбоко аплициране на сперма, разредена със среда, съдържаща новият биокомпонент (сепарирани ниско молекулни спермално плазмени протеини (до 30 кДа), води до добра заплодяемост (до 75%). При използване на високо молекулни спермално плазмени протеини заплодяемостта на овцете е незадоволителна (до 5% за фракция 6 и до 45% – за фракция 4).
ПРИНОСИ1. Разработена е модифицирана методика на Comet assay за
изследване интегритета на ДНК .2. Разработен е оптимизиран кристалоиден разредител, на базата на
среда 6А, съдържащ нов биокомпонент.

3. Разработен е подход за успешно нискотемпературно съхранение на сперматозоиди от коч, основан на нов механизъм на протекция с биокомпоненти – сепарирани и селектирани нискомолекулни спермално плазмени протеини.
ПРЕДЛОЖЕНИЯ ЗА ВНЕДРЯВАНЕ В ПРАКТИКАТА
Предлагаме нов кристалоиден разредител съдържащ нов биокомпонент - селектирани ниско молекулни спермално плазмени протеини, за съхраняване на сперматозоиди от коч в охладено състояние.
Практическо използване за ИО на овце.

ПУБЛИКАЦИИ ВЪВ ВРЪЗКА С ДИСЕРТАЦИОННИЯ ТРУД
1. Kukov A, Daskalova D, Ivanova-Kicheva M, Stefanov R, Neicheva A. Immunocytochemical localization of separated seminal plasma proteins on the ram’s sperm plasma membrane during in vitro preservation at 40C. Biotechnology in Animal husbandry, 25 (5-6):925-934 (2009). ISSN: 1450-9156.
2. Dimitrov A, Ivanova-Kicheva M, Daskalova D, Kacheva D and Solenkova E. Protective role of separated seminal plasma proteins in in vitro storage of ram spermatozoa. Journal of Mountain Agriculture on the Balkans, 10, 5:922-932 (2007). ISSN: 1311-0489.
3. Solenkova E, Daskalova D, Kukov A, Ivanova-Kicheva M, Georgiev V. The influence of cryopreservation on cell morphology and phospholipids asymmetry of plasma membrane in ram spermatozoa. Journal of Mountain Agriculture on the Balkans, 11, 6: 982-999 (2008). ISSN: 1311-0489.
4. Куков А, Запрянова С, Даскалова Д, Минчева Ж, Герзилов В, Нейчева А, Иванова-Кичева М, Димов Г. Компютърен спермоанализ на протективната роля на семинално плазмени протеини при кочове от породата черноглава плевенска. Аграрни науки 2010, in press. ISSN: 1313-6577.
УЧАСТИЕ В НАУЧНИ ФОРУМИ
• Denica Daskalova, Alexander Kukov, Maria Ivanova – Kicheva. Chromatography separated proteins of seminal plasma provide protective effects to ram spermatozoa to maintain motility and survivor rate. 12 th
International Symposium of Immunology of Reproduction, 25-27 June, Varna, 2009.
• Kukov А, D. Daskalova, M.Ivanova-Kicheva, R.Stefanov, A. Neicheva. Immunocytochemical localization of separated seminal plasma proteins on the ram’s sperm plasma membrane during in vitro preservation at 40C. 9th
International symposium on animal husbandry. Modern trends in livestock production. 7-9 october. 2009
• Ivanova –Kicheva M, Kukov A, Daskalova D. Immunocytochemical localization of seminal plasma proteins on ram spermatozoa plasma membrane during the low temperature anabiosis. Conference of the Balkan network for biotechnology in animal reproduction. 18-19.02.2009.
• Даскалова Д, Куков А, Кичева. М., Петров М. Влияние на сепарирани спермално плазмени протеини върху преживяемостта на сперматозоиди от коч при съхранение ма 40С. Научна Сесия Ветеринарен Факултет, ЛУ София 2007
• Denica Daskalova, Alexander Kukov, Maria Ivanova - Kicheva. Chromatography separation of seminal plasma proteins and their effect on the viability of ram spermatozoa. XI Anniversary Scientific Conference, 120 Years of Academic Education in Biology, 45 Years Faculty of biolgy, 27 – 29 may, Sofia, 2009.


















