
-1992 Kruse Conversion of Human Interleukin-4 Into a High Affinity Antagonist by a Single Amino Acid...
8
The EMBO Journal vol. 11 no.9 pp.3237 - 3244, 1992 Conversion of human interleukin-4 into a high affinity antagonist by a single amino acid replacement N.Kruse, H.-P.Tony' and W.Sebald2 Theodor-Boveri-Institut fUir Biowissenschaften (Biozentrum) der Universitat Wurzburg, Physiologische Chemie II, Am Hubland, D-8700 Wurzburg and 'Medizinische Poliklinik der UniversitSt, Klinikstr. 6-8, 8700 Wurzburg, Germany 2Corresponding author Communicated by H.Michel Interleukin-4 (IL14) represents a prototypic lymphokine (for a recent review see Paul, 1991). It promotes differentiation of B-cells and the proliferation of T- and B-cell, and other cell types of the lymphoid system. An antagonist of human IL-4 was discovered during the studies presented here after Tyrl24 of the recombinant protein had been substituted by an aspartic acid residue. This IL-4 variant, Y124D, bound with high affinity to the IL-4 receptor (KD = 310 pM), but retained no detectable proliferative activity for T-cells and inhibited IL-4-dependent T-cell proliferation competitively (K; = 620 pM). The loss of efficacy in variant Y124D was estimated to be > 100-fold on the basis of a weak partial agonist activity for the very sensitive induction of CD23 positive B-cells. The subsitution of Tyrl24 by either phenylalanine, histidine, asparagine, lysine or glycine resulted in partial agonist variants with unaltered receptor binding affinity and relatively small deficiencies in efficacy. These results demonstrate that high affinity binding and signal generation can be uncoupled efficiently in a ligand of a receptor belonging to the recently identified hematopoietin receptor family. In addition we show for the first time, that a powerful antagonist acting on the IL4 receptor system can be derived from the IL4 protein. Key words: drug design/partial agonists/receptor signalling Introduction Interleukin-4 (IL-4) represents a typical immunoregulatory lymphokine (for review see Paul and Ohara, 1987; Finkelman et al., 1990; Paul, 1991). It is produced mainly by activated T-cells and mast cells and has a wide range of biological activities on various cell types of the lymphoid system. B-cells are stimulated in vitro to express class II major histocompatibility complex (MHC H) molecules, the IgE low affinity receptor (FceRII, CD23) and immuno- globulins class E and GI. DNA synthesis is induced in activated B-cells, as well as in mature T-cells, foetal thymocytes, mast cells and others. Mice treated with neutralizing antibodies directed against IL-4 or against the IL-4 receptor (Finkelman et al., 1990; Urban,Jr et al., 1991), as well as mice bearing an inactivated IL-4 gene (Kuhn et al., 1991), are unable to produce IgE and show reduced serum levels of IgG 1 after challenge with () Oxford University Press the relevant stimuli. This indicates a physiological role of IL-4 at least in the regulation of IgE levels, most likely by acting as an isotype switching factor. Consequently, IL-4 may exert a pathophysiological role in the generation of disease states, as for example hyper-IgE syndrome or IgE- mediated allergic conditions (see Finkelman et al., 1990; Tepper et al., 1990). IL-4 function is mediated by its binding to plasma membrane receptors occurring at the relatively small numbers of 150-2500 molecules per target cell (Cabrillat et al., 1987; Park et al., 1987). A single dissociation constant of - 100 pM has been determined for this inter- action. The IL-4 receptor probably comprises only a single polypeptide chain of - 800 amino acid residues as suggested by chemical crosslinking experiments using 125I labelled IL-4 (Mosley et al., 1989; Galizzi et al., 1990; Harada et al., 1990; Idzerda et al., 1990). Binding to IL-4 in vitro is not influenced by structural elements apart from the extracellular portion of the receptor, since similar dissociation constants were found for association with IL-4 with the entire receptor and only the ligand binding domain in the case of both the mouse and the human protein (Maliszewski et al., 1990; Garrone et al., 1991). At pre- sent it is unclear if a second protein aggregates with the receptor after IL-4-induced activation. The IL-4 receptor is evolutionarily related to the large family of hematopoietin receptors, including receptors (or receptor subunits) for interleukins-2, 3, 4, 5, 6 and 7, for GM- and G-CSF and also for erythropoietin, growth hormone and prolactine (Bazan, 1990a,b; Cosman et al., 1990). All these proteins show particular homologies in the amino acid sequence of their extracellular ligand binding domains. NMR studies revealed a three-dimensional structure of human IL-4 comprising a four-helix bundle motif with an up-up-down-down connectivity (Redfield et al., 1991) similar to both human and porcine growth hormone (Abdel- Meguid et al., 1987; De Vos et al., 1992). Other ligands binding to members of the haematopoietin receptor family also appear to be helically organized proteins (see Bazan, 1990a,b). The crystal structure of human IL-2 shows a four- helix bundle (Brandhuber et al., 1987) albeit with a connectivity different from IL-4. The amino acid sequences of these ligands display no clear homologies. A puzzling similarity, however, is apparent in the pattern of amino acid residues forming one surface of a putative amphipathic helix at the C-terminus of these proteins (Sanderson et al., 1988; see Bazan, 1990b). Sequence positions involved in receptor binding have been identified in several ligands of the haematopoietin receptor family, e.g. in IL-2 (Collins et al., 1988; Weigel et al., 1989; Zurawski and Zurawski, 1989), growth hormone (Cunningham and Wells, 1991; Cunningham et al., 1991), GM-CSF and IL-5 (Shanafelt et al., 1991). Amino acid residues involved in signal generation, however, have been more difficult to identify. In order to address this issue, contributions of particular 3237
-
Upload
nilabh-ranjan -
Category
Documents
-
view
219 -
download
0
Transcript of -1992 Kruse Conversion of Human Interleukin-4 Into a High Affinity Antagonist by a Single Amino Acid...

The EMBO Journal vol. 1 1 no.9 pp.3237 - 3244, 1992
Conversion of human interleukin-4 into a high affinityantagonist by a single amino acid replacement
N.Kruse, H.-P.Tony' and W.Sebald2Theodor-Boveri-Institut fUir Biowissenschaften (Biozentrum) derUniversitat Wurzburg, Physiologische Chemie II, Am Hubland,D-8700 Wurzburg and 'Medizinische Poliklinik der UniversitSt,Klinikstr. 6-8, 8700 Wurzburg, Germany
2Corresponding author
Communicated by H.Michel
Interleukin-4 (IL14) represents a prototypic lymphokine(for a recent review see Paul, 1991). It promotesdifferentiation of B-cells and the proliferation of T- andB-cell, and other cell types of the lymphoid system. Anantagonist of human IL-4 was discovered during thestudies presented here after Tyrl24 of the recombinantprotein had been substituted by an aspartic acid residue.This IL-4 variant, Y124D, bound with high affinity tothe IL-4 receptor (KD = 310 pM), but retained nodetectable proliferative activity for T-cells and inhibitedIL-4-dependent T-cell proliferation competitively(K; = 620 pM). The loss of efficacy in variant Y124Dwas estimated to be > 100-fold on the basis of a weakpartial agonist activity for the very sensitive inductionof CD23 positive B-cells. The subsitution of Tyrl24 byeither phenylalanine, histidine, asparagine, lysine orglycine resulted in partial agonist variants with unalteredreceptor binding affinity and relatively small deficienciesin efficacy. These results demonstrate that high affinitybinding and signal generation can be uncoupled efficientlyin a ligand of a receptor belonging to the recentlyidentified hematopoietin receptor family. In addition weshow for the first time, that a powerful antagonist actingon the IL4 receptor system can be derived from the IL4protein.Key words: drug design/partial agonists/receptor signalling
IntroductionInterleukin-4 (IL-4) represents a typical immunoregulatorylymphokine (for review see Paul and Ohara, 1987;Finkelman et al., 1990; Paul, 1991). It is produced mainlyby activated T-cells and mast cells and has a wide range ofbiological activities on various cell types of the lymphoidsystem. B-cells are stimulated in vitro to express class IImajor histocompatibility complex (MHC H) molecules, theIgE low affinity receptor (FceRII, CD23) and immuno-globulins class E and GI. DNA synthesis is induced inactivated B-cells, as well as in mature T-cells, foetalthymocytes, mast cells and others.Mice treated with neutralizing antibodies directed against
IL-4 or against the IL-4 receptor (Finkelman et al., 1990;Urban,Jr et al., 1991), as well as mice bearing an inactivatedIL-4 gene (Kuhn et al., 1991), are unable to produce IgEand show reduced serum levels of IgG1 after challenge with
() Oxford University Press
the relevant stimuli. This indicates a physiological role ofIL-4 at least in the regulation of IgE levels, most likely byacting as an isotype switching factor. Consequently, IL-4may exert a pathophysiological role in the generation ofdisease states, as for example hyper-IgE syndrome or IgE-mediated allergic conditions (see Finkelman et al., 1990;Tepper et al., 1990).
IL-4 function is mediated by its binding to plasmamembrane receptors occurring at the relatively smallnumbers of 150-2500 molecules per target cell (Cabrillatet al., 1987; Park et al., 1987). A single dissociationconstant of - 100 pM has been determined for this inter-action. The IL-4 receptor probably comprises only a singlepolypeptide chain of - 800 amino acid residues as suggestedby chemical crosslinking experiments using 125I labelledIL-4 (Mosley et al., 1989; Galizzi et al., 1990; Haradaet al., 1990; Idzerda et al., 1990). Binding to IL-4 in vitrois not influenced by structural elements apart from theextracellular portion of the receptor, since similardissociation constants were found for association with IL-4with the entire receptor and only the ligand binding domainin the case of both the mouse and the human protein(Maliszewski et al., 1990; Garrone et al., 1991). At pre-sent it is unclear if a second protein aggregates with thereceptor after IL-4-induced activation. The IL-4 receptor isevolutionarily related to the large family of hematopoietinreceptors, including receptors (or receptor subunits) forinterleukins-2, 3, 4, 5, 6 and 7, for GM- and G-CSF andalso for erythropoietin, growth hormone and prolactine(Bazan, 1990a,b; Cosman et al., 1990). All these proteinsshow particular homologies in the amino acid sequence oftheir extracellular ligand binding domains.NMR studies revealed a three-dimensional structure of
human IL-4 comprising a four-helix bundle motif with anup-up-down-down connectivity (Redfield et al., 1991)similar to both human and porcine growth hormone (Abdel-Meguid et al., 1987; De Vos et al., 1992). Other ligandsbinding to members of the haematopoietin receptor familyalso appear to be helically organized proteins (see Bazan,1990a,b). The crystal structure of human IL-2 shows a four-helix bundle (Brandhuber et al., 1987) albeit with aconnectivity different from IL-4. The amino acid sequencesof these ligands display no clear homologies. A puzzlingsimilarity, however, is apparent in the pattern of amino acidresidues forming one surface of a putative amphipathic helixat the C-terminus of these proteins (Sanderson et al., 1988;see Bazan, 1990b). Sequence positions involved in receptorbinding have been identified in several ligands of thehaematopoietin receptor family, e.g. in IL-2 (Collins et al.,1988; Weigel et al., 1989; Zurawski and Zurawski,1989), growth hormone (Cunningham and Wells, 1991;Cunningham et al., 1991), GM-CSF and IL-5 (Shanafeltet al., 1991). Amino acid residues involved in signalgeneration, however, have been more difficult to identify.In order to address this issue, contributions of particular
3237
N.Kruse, H.-P.Tony and W.Sebald
amino acids to receptor affinity and biological activity,respectively, have to be distinguishable. As yet, uncouplingof receptor binding from biological effects has been observedto some extent only in the case of certain mutant derivativesof both human and murine IL-2 (Liang et al., 1988;Zurawski et al., 1990).
Recently we have generated a set of mutant variants ofhuman IL-4 with single substitutions at the positions of allcysteinyl and aromatic residues (Kruse et al., 1991). Oneof these variants, Y124D, was completely inactive even atmicromolar concentrations during a T-cell proliferationassay. Subsequently, we realized that variant Y124Defficiently inhibits IL4 induced T-cell proliferation. Thispromoted the generation of IL-4 variants in which Tyrl24was substituted by residues differing in size, polarity orcharge, respectively. The receptor binding affinity of theseisolated mutant proteins was determined, as well as theirbiological activities in two in vitro assays differing insensitivity by nearly two orders of magnitude. The resultsof these experiments indicated that the nature of the sidechain at position 124 of human IL-4 dramatically affectssignal generation and biological activity, while the influenceon receptor binding is marginal.The demonstration of the feasibility to obtain an efficient
antagonist of the IL4 receptor system has several importantimplications. First, inhibitory IL-4 variants representpotential drugs helpful for example in the treatment of IgE-mediated diseases (see e.g. Finkelman et al., 1990; Paul,1991) similar to therapeutic monoclonal antibodies againstIL-4 or to the soluble extracellular IL-4 receptor domain(Maliszewski et al., 1990; Garrone et al., 1991). Secondly,antagonistic ligands may prove valuable tools for theinvestigation of IL-4 receptor-mediated signal transducingmechanisms. Thirdly, considering the homologies andsimilarities mentioned above, it is an intriguing possibilitythat similar antagonistic variants will be found for otherligands of this receptor family (see also Zurawski et al.,1990).
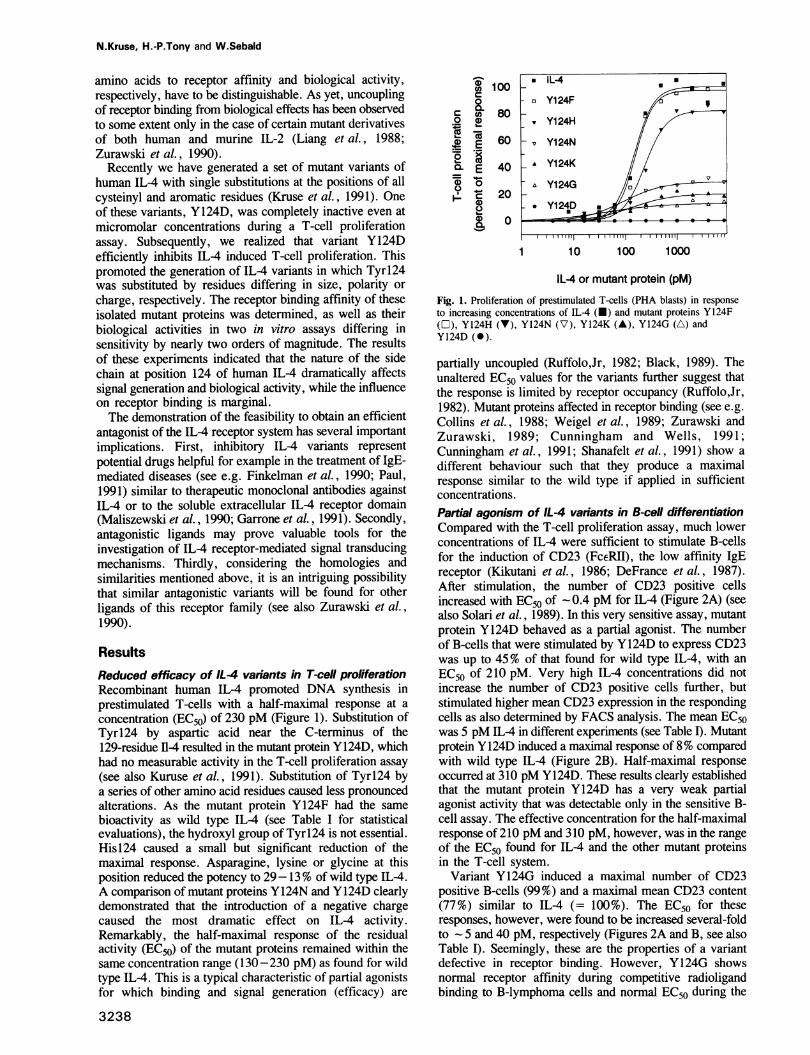
ResultsReduced efficacy of IL4 variants in T-cell proliferationRecombinant human IL-4 promoted DNA synthesis inprestimulated T-cells with a half-maximal response at aconcentration (EC50) of 230 pM (Figure 1). Substitution ofTyrl24 by aspartic acid near the C-terminus of the129-residue I14 resulted in the mutant protein Y124D, whichhad no measurable activity in the T-cell proliferation assay(see also Kuruse et al., 1991). Substitution of Tyrl24 bya series of other amino acid residues caused less pronouncedalterations. As the mutant protein Y124F had the samebioactivity as wild type IL4 (see Table I for statisticalevaluations), the hydroxyl group of Tyrl24 is not essential.His124 caused a small but significant reduction of themaximal response. Asparagine, lysine or glycine at thisposition reduced the potency to 29-13% of wild type IL-4.A comparison of mutant proteins Y124N and Y124D clearlydemonstrated that the introduction of a negative chargecaused the most dramatic effect on IL-4 activity.Remarkably, the half-maximal response of the residualactivity (EC50) of the mutant proteins remained within thesame concentration range (130-230 pM) as found for wildtype IL-4. This is a typical characteristic of partial agonistsfor which binding and signal generation (efficacy) are
3238
ID 100a IL-4 .0Yl124F
80.0 E Y124H(D 60 v,Y1 24N
QL E 40 _ Y124K
~~2~20 AYY24GA
0220 X
11Y12 1D IAitliii
1 10 100 1000
IL4 or mutant protein (pM)
Fig. 1. Proliferation of prestimulated T-cells (PHA blasts) in responseto increasing concentrations of IL-4 (M) and mutant proteins Y124F(O), Y124H (V), Y124N (V), Y124K (A), Y124G (A) andY124D (0).
partially uncoupled (Ruffolo,Jr, 1982; Black, 1989). Theunaltered EC50 values for the variants further suggest thatthe response is limited by receptor occupancy (Ruffolo,Jr,1982). Mutant proteins affected in receptor binding (see e.g.Collins et al., 1988; Weigel et al., 1989; Zurawski andZurawski, 1989; Cunningham and Wells, 1991;Cunningham et al., 1991; Shanafelt et al., 1991) show adifferent behaviour such that they produce a maximalresponse similar to the wild type if applied in sufficientconcentrations.Partial agonism of IL-4 variants in B-cell differentiationCompared with the T-cell proliferation assay, much lowerconcentrations of IL-4 were sufficient to stimulate B-cellsfor the induction of CD23 (FceRII), the low affinity IgEreceptor (Kikutani et al., 1986; DeFrance et al., 1987).After stimulation, the number of CD23 positive cellsincreased with EC50 of -0.4 pM for IL4 (Figure 2A) (seealso Solari et al., 1989). In this very sensitive assay, mutantprotein Y124D behaved as a partial agonist. The numberof B-cells that were stimulated by Y124D to express CD23was up to 45% of that found for wild type IL-4, with anEC50 of 210 pM. Very high IL-4 concentrations did notincrease the number of CD23 positive cells further, butstimulated higher mean CD23 expression in the respondingcells as also determined by FACS analysis. The mean EC50was 5 pM IL-4 in different experiments (see Table I). Mutantprotein Y124D induced a maximal response of 8% comparedwith wild type IL-4 (Figure 2B). Half-maximal responseoccurred at 310 pM Y124D. These results clearly establishedthat the mutant protein Y124D has a very weak partialagonist activity that was detectable only in the sensitive B-cell assay. The effective concentration for the half-maximalresponse of 210 pM and 310 pM, however, was in the rangeof the EC50 found for IL-4 and the other mutant proteinsin the T-cell system.
Variant Y124G induced a maximal number of CD23positive B-cells (99%) and a maximal mean CD23 content(77%) similar to IL-4 (= 100%). The EC50 for theseresponses, however, were found to be increased several-foldto 5S and 40 pM, respectively (Figures 2A and B, see alsoTable I). Seemingly, these are the properties of a variantdefective in receptor binding. However, Y124G showsnormal receptor affinity during competitive radioligandbinding to B-lymphoma cells and normal EC50 during the

Human IL-4 antagonist mutant proteins
Table I. Effective concentration (EC50) and maximal response (Rmax) of IL-4 and I1-4 variants evaluated from T- and B-cell assays
Protein T-cell proliferation B-cell (spleen)EC50 (pM) Rmax (%) Mean CD23 content Number of CD23 positive cells
EC50 (pM) Rmax (%) EC50 (pM) Rmax (%)
IL4 230 (140-390) 100 + 7 5 (2.5-10) 100 + 11 0.4 (0.1-1.6) 100 6Variant Y124F 150 (110-210) 97 ± 6Variant Y 124H 190 (130-290) 82 ± 5Variant Y124N 220 (170-290) 29 ± 7Variant Y124K 230 (210-250) 20 ± 4Variant Y124G 130 (120-140) 13 ± 4 -40 -77 -5 -99Variant Y124D <0.5 ± 0.7 310 (160-620) 8 ± 3 -210 (120-360) 45 ± 6
EC50 values were distributed log normal. Numbers in brackets were calculated from log EC50 ± log SD.Rmax values + SD were related to Rmax of IL-4 as 100%.Approximate values (-) were obtained from a single experiment.
T-cell proliferation assay (see Tables I and II). Thus, theproperties of partial agonist Y124G are consistent with anIL-4 receptor system in B-cells operating at low occupancyduring CD23 induction. In this case, the EC50 values fora partial agonist are expected to increase proportionally tothe loss of efficacy (Ruffolo,Jr, 1982).The CD23 positive cells represented 67% of the total cells
analysed by FACS. This percentage remained constant atall saturation levels. Thus, the induction of the number ofCD23 positive cells and the induction of mean CD23 contentmost probably reflected two aspects of the same process inthe same cell population. Of two alternatives-either eachinduced cell immediately develops a full CD23 content orthe whole population is first induced to develop a low CD23content which upon higher stimulation is increased-thesecond is consistent with the observed data.
Competitive receptor bindingThe dose-response curves suggest that the amino acidexchanges at position 124 did not dramatically alter thereceptor affinity of the mutant proteins compared with wildtype IL-4. This has been corroborated by radioligandreceptor binding experiments (Figure 3). IL-4, as well asthe mutant proteins studied here, competed efficiently withiodinated IL-4 for the IL-4 receptors present on the B-lymphoma cell line Raji (Cabrillat et al., 1987; Park et al.,1987) as shown in Figure 3 for Y124G and Y124D. Thedisplacement curves of all variants, with the exception ofY124D, were not significantly different from that of IL-4(see also Table II). Half-maximal displacement of iodinatedIL-4 required a 3.4-fold higher concentration of mutantprotein Y124D compared with that of IL-4. Correspondingcompetitive binding studies performed with both PHA-activated T-cells and activated tonsillar B-cells yielded similarrelative IC50 values (Table II). (Unfortunately, the largenumbers of activated B-cells necessary for radioligandbinding could not routinely be obtained from human spleen).The concentration of 1 nM ['251]IL-4 during the
competitive binding experiments was - 10-fold higher thanthe dissociation constant. Thus, the KD of the competingligands could not be determined from the IC50 (Munson,1983). The KD Of [251I]IL4 determined independently (seeMaterials and methods) was 91 pM for the B-lymphoma cellsand 81 pM for the PHA-activated T-cells. Assuming areceptor dissociation constant KD of - 100 pM for IL-4(see also Cabrillat et al., 1987; Park et al., 1987), mutantprotein Y124D accordingly had a KD of 310 pM.
m
Cl
0Q.4)
Cr-F00CLco
0
C.
0)
0C 10000.02D 80a.E 60
E 400Co 200)8 00
0
00@ 100 - . IL-40 A-*0
m 2 80 *_̂ Y124D6060
0 ~~~~~~~b0 E 40-CIOcmJ
0.01 0.1 1 10 100 1000
ILA or mutant protein (pM)
Fig. 2. Induction of the low affinity Fcc receptor (CD23) on B-cells inresponse to increasing concentrations of ILL4 (U) or mutant proteinsY124G (A) and Y124D (0). (a) Number of CD23 positive B-cells aspercent of maximal response. (b) Mean CD23 content of B-cells.
Activated human B- and T-cells both expose 500-1000molecules of IL-4 receptor at their surface showing a KDof 100 pM in terms of IL4 binding (see above). Thereforethe proliferative response ofT-cells appears to closely followthe receptor occupancy. Assuming that in the B-cell assaysthe biological response is solely mediated by the receptorwith a KD of 100 pM, the induction of CD23 requires theoccupancy of only a few percent of the receptors by IL-4.We suggest that only in case of the 'emasculated' (Black,1989) Y124D mutant protein, the receptor had to bemaximally saturated in order to generate a small partialresponse. It remains unclear, however, if the assay periods
3239
N.Kruse, H.-P.Tony and W.Sebald
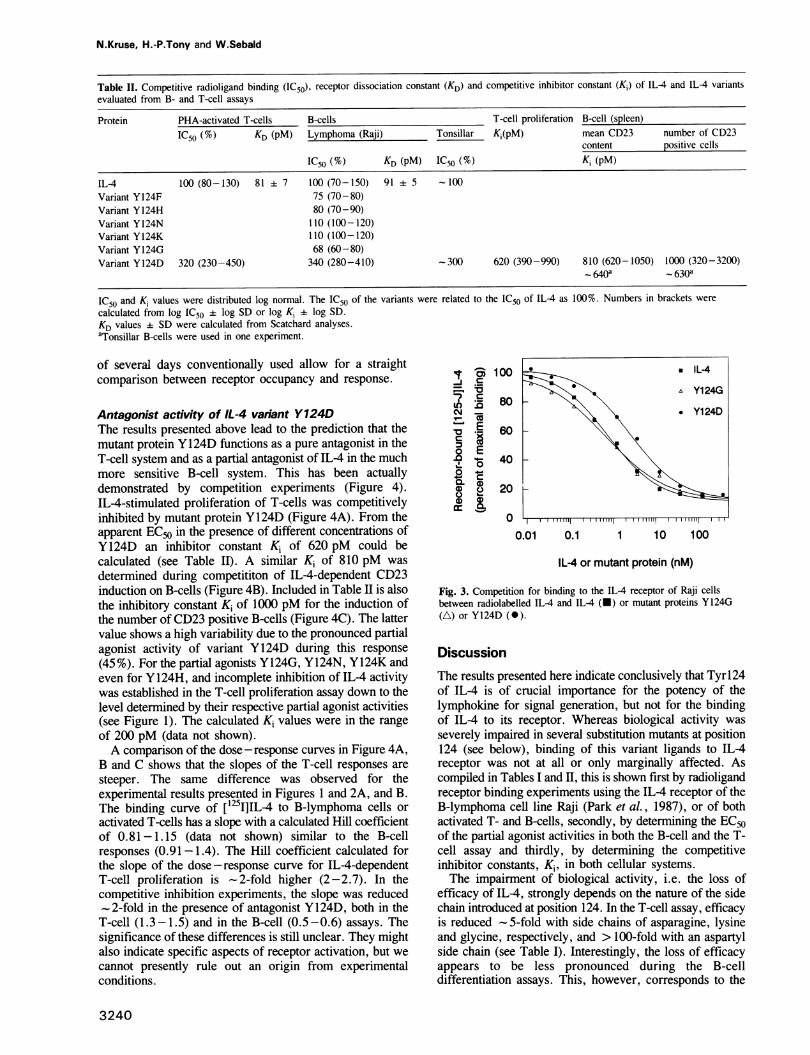
Table II. Competitive radioligand binding (IC50), receptor dissociation constant (KD) and competitive inhibitor constant (Ki) of IL-4 and IL-4 variantsevaluated from B- and T-cell assays
Protein PHA-activated T-cells B-cells T-cell proliferation B-cell (spleen)IC50 (%) KD (pM) Lymphoma (Raji) Tonsillar Ki(pM) mean CD23 number of CD23
content positive cells
IC50 (%) KD (pM) IC50 (%) Ki (pM)
IL-4 100 (80-130) 81 7 100 (70-150) 91 5 - 100Variant Y 124F 75 (70-80)Variant Y124H 80 (70-90)Variant Y124N 110 (100-120)Variant Y124K 110 (100-120)Variant Y 124G 68 (60-80)Variant Y124D 320 (230-450) 340 (280-410) -300 620 (390-990) 810 (620-1050) 1000 (320-3200)
0640a - 630a
IC50 and Ki values were distributed log normal. The IC50 of the variants were related to the IC50 of IL-4 as 100%. Numbers in brackets werecalculated from log IC50 + log SD or log Ki ± log SD.KD values + SD were calculated from Scatchard analyses.aTonsillar B-cells were used in one experiment.
of several days conventionally used allow for a straightcomparison between receptor occupancy and response.
Antagonist activity of IL-4 variant Y124DThe results presented above lead to the prediction that themutant protein Y124D functions as a pure antagonist in theT-cell system and as a partial antagonist of IL-4 in the muchmore sensitive B-cell system. This has been actuallydemonstrated by competition experiments (Figure 4).IL-4-stimulated proliferation of T-cells was competitivelyinhibited by mutant protein Y124D (Figure 4A). From theapparent EC50 in the presence of different concentrations ofY124D an inhibitor constant Ki of 620 pM could becalculated (see Table I1). A similar Ki of 810 pM wasdetermined during competititon of IL-4-dependent CD23induction on B-cells (Figure 4B). Included in Table II is alsothe inhibitory constant Ki of 1000 pM for the induction ofthe number of CD23 positive B-cells (Figure 4C). The lattervalue shows a high variability due to the pronounced partialagonist activity of variant Y124D during this response(45 %). For the partial agonists Y 124G, Y 124N, Y124K andeven for Y124H, and incomplete inhibition of IL-4 activitywas established in the T-cell proliferation assay down to thelevel determined by their respective partial agonist activities(see Figure 1). The calculated K, values were in the rangeof 200 pM (data not shown).A comparison of the dose -response curves in Figure 4A,
B and C shows that the slopes of the T-cell responses aresteeper. The same difference was observed for theexperimental results presented in Figures 1 and 2A, and B.The binding curve of [1251]IL-4 to B-lymphoma cells oractivated T-cells has a slope with a calculated Hill coefficientof 0.81-1.15 (data not shown) similar to the B-cellresponses (0.91-1.4). The Hill coefficient calculated forthe slope of the dose-response curve for IL-4-dependentT-cell proliferation is -2-fold higher (2-2.7). In thecompetitive inhibition experiments, the slope was reduced-2-fold in the presence of antagonist Y124D, both in the
T-cell (1.3-1.5) and in the B-cell (0.5-0.6) assays. Thesignificance of these differences is still unclear. They mightalso indicate specific aspects of receptor activation, but wecannot presently rule out an origin from experimentalconditions.
; ' 80̂ Y124GCM8r Yl 24D
;E 60
4-0
0.0o 20H
0.01 0.1 1 10 100
IL-4 or mutant protein (nM)
Fig. 3. Competition for binding to the IL-4 receptor of Raji cellsbetween radiolabelled IL-4 and IL-4 (U) or mutant proteins Y124G(A) or Y124D ()
DiscussionThe results presented here indicate conclusively that Tyrl24of IL4 is of crucial importance for the potency of thelymphokine for signal generation, but not for the bindingof IL-4 to its receptor. Whereas biological activity wasseverely impaired in several substitution mutants at position124 (see below), binding of this variant ligands to IL4receptor was not at all or only marginally affected. Ascompiled in Tables I and II, this is shown first by radioligandreceptor binding experiments using the IL-4 receptor of theB-lymphoma cell line Raji (Park et al., 1987), or of bothactivated T- and B-cells, secondly, by determining the EC50of the partial agonist activities in both the B-cell and the T-cell assay and thirdly, by determining the competitiveinhibitor constants, Ki, in both cellular systems.The impairment of biological activity, i.e. the loss of
efficacy of IL4, strongly depends on the nature of the sidechain introduced at position 124. In the T-cell assay, efficacyis reduced - 5-fold with side chains of asparagine, lysineand glycine, respectively, and > 100-fold with an aspartylside chain (see Table I). Interestingly, the loss of efficacyappears to be less pronounced during the B-celldifferentiation assays. This, however, corresponds to the
3240

Human IL-4 antagonist mutant proteins
D120
X 100o
80
Ec- 0
60
Q ECL E 40
-6
0, 20
0
0 I
10
=0 o 120
9 °10000
8
0CO
0
cvw 40
o 4, 200 E
.U
0
.0E
0
0
U1)
CO
Ec
C
(I)
2'a
c
CD
r
120
100
80
60
40
20
0
10c
1
0.1
Fig. 4. Competitive inhibition of IL
(a) IL-4-induced CD23 content of B
of CD23 positive B-cells (c) by mut
concentrations of 0 nm (U), 0.67 n
(A). The results in (B) and (C) are
agonist activities of variant Y124D.
- 40-fold lower EC50 of IL-4
or the 500-fold lower EC50facells when compared with th
These apparent discrepancexplained in terms of recept
- 200 pM for T-cell prolifeireceptor dissociation constant
may be limited by the numbe
B-cell response appears to be
following receptor activation.
the EC,O of 5 pM (0.4 pM'
correspond to a receptor occu
content) or 0.4% (number
calculation and its underlying assumptions are detailed inMaterials and methods. The lower efficacy of the IL-4
M) variants can thus be partially compensated for by higherXj*/ receptor saturation. In case of mutant protein Y124D
maximal response is 8% (45%) with an EC50 or 310 pM1) / / / / (210 pM) for mean CD23 induction (CD23 positive cells).
According to these data, the loss of efficacy is 125- toa°/^/ 300-fold. During the T-cell proliferation assays, the mean
error of the control was 0.5% of the maximal response of
IL-4. The responses of variant Y124D were within the meanerror of the control. Thus, the efficacy of Y124D is at least
l11111 l l 200-fold lower than that of IL4 in this system. Although1000 10000 these calculations are rough estimates derived from long term
assays, measuring complex proliferation and differentiationprocesses, at least they should provide information concern-ing the upper limit. (An alternative explanation of the resultsinvolving a very small subpopulation of receptors, e.g.20-50 molecules/cell, with a very low dissociation constant
in the range of 1 pM in the B-cells appears rather unlikely,o/ < since the antagonist Y124D inhibits with the same inhibitorybz/ constant of 620-1000 pM both B- and T-cells.)b The identification of a whole panel of partial agonists
suggests that the residue at position 124 is directly involvedin signal generating interactions. The amino acid exchangein mutant protein Y124D, however, produces an
unexpectedly large effect. Thus, additional conformation
10 100 1000 effects appear to be possible in this case. It may be relevantin this respect that variant Y124D does not easily crystallize(T.Muller and W.Sebald, unpublished observation) incontrast to IL-4 (Cook et al., 1991). It remains to beestablished if further residues in the IL-4 molecule, togetherwith Tyrl24, form a site distinct from a IL-4 receptorbinding site, which is involved in signal generation.According to the available structural information (Redfield
cC et al., 1991; see also Diederichs et al., 1991) residues at
the C-terminal end of helix D are close to Tyrl24. Cys127forms a disulfide bond to Cys3. Thus, side chains of helixA may also be in proximity to Tyrl24. It is interesting tonote that for human GM-CSF, which has the same fold as
1 10 100 IL-4, a receptor binding site was postulated consisting of
two peptide stretches centered around Arg24 (helix A) and
IL-4 (pM) Met79 (helix C) (Diederichs et al., 1991). According to the
GM-CSF crystal structure, this binding site would be
4-dependent T-cell proliferation. spatially far away from the C-terminus of helix D.
-cells (b) andIL-4-induced number Tyrl24 of human IL-4 is found at the correspondingtant protein Y124D at position of murine IL-4 (Sanderson et al., 1988). It occursM(O), 2 nM (A), and 6.7 nM at the end of helix D (Redfield et al., 1991), only six residuescorrected by subtracting partial from the C-terminus. It has been noted (Bazan, 1990b;
Sanderson, 1988) that this C-terminal segment has featuresin common with other known ligands of the haematopoietin
for the mean CD23 induction receptor family. Amino acid substitutions of Glu141 ofr induction of CD23 positive murineIL-2 (Zurawski et al., 1990), whose position appearsie T-cell response. to be equivalent to Tyrl24 of human IL-4, yielded a series
-ies in IL-4 efficacy can be of partial agonists. In particular, murine IL-2 variant Q141Dtor occupancy. The EC50of was inactive or only weakly active in proliferation assaysration is similar to the IL-4 using various cell lines. However, high affinity binding wast. Accordingly, this response lost and no competitive inhibition of IL-2-dependent:r of activated receptors. The proliferation of HT2 cells could be found even at a
limited by a signalling step 5 x104-fold molar excess of variant Q141D. Variant
his step then would determine Q141D of murine IL-2 is a capable antagonist, however,in these processes, which against mutant proteins of IL-2 with receptor-binding defects.
pancy of -5% (mean CD23 mIL-2 variants with substitutions of Glu141 by asparaginef CD23 positive cells). The (class I), lysine (class II), aspartic acid (class HI) and glycine
3241
- * Y124D (OnM)- o Y124D (0.67nl
_ A Y124D (2nM)_ & Y124D (6.7nMJ
II 1 11 I1

N.Kruse, H.-P.Tony and W.Sebald
(class IV), could be grouped into four classes according totheir differing partial agonist activities. This is unlike thevariants of human IL-4, where substitutions of Tyrl24 byasparagine, lysine and glycine produced very similar agonistactivities (see Figure 1, Table I). Thus, it remains unclearin how far the generation of partial agonism by an aspartylside chain at the human IL-4 Y124 and at the murine IL-2Q141 positions indicates comparable structural requirements.Two non-overlapping binding sites were identified in
human growth hormone (Cunningham and Wells, 1991;Cunningham et al., 1991; De Vos et al., 1992) that areoccupied in a sequential manner by two growth hormonereceptor molecules. Site 1 comprises the C-terminus of helixD (Cunningham et al., 1991). The equivalent of IL-4 Tyrl24might be human growth hormone Val 185, which is locatedat the end of helix D and which is part of site 1. Thebiological activity of a growth hormone variant V185D hasnot yet been assessed. It was postulated (Cunningham et al.,1991) that non-dimerizing variants of growth hormone,which have lost the ability to bind two receptor molecules,should show antagonistic properties.A natural receptor antagonist was detected in the
interleukin 1 system. The human protein (IL-iRa) shares19% identical amino acid positions with IL-la and 26%identical positions with IL-1: and is probably composed of12 f-strands. It behaves as a pure antagonist in binding withhigh affinity to the IL-1 receptor (type I) without evokingany detectable response (Eisenberg et al., 1990; Hannumet al., 1990). This indicates that in the IL-I system receptorbinding and receptor activation are completely separable.An efficient uncoupling of both steps also was obtained bysingle amino acid replacements in IL-I molecules. IL-lI$variants R127G (Gehrke et al., 1990) and D145K (Ju et al.,1991) as well as an IL-la variant D15 1Y (Yamayoshi et al.,1990) exhibited little biological activity but bound with alargely unaltered affinity to the receptor. The discontinuousreceptor binding site identified in human IL-1,B (Labriola-Tompkins et al., 1991) clusters around Arg4 and does notcomprise the positions determining receptor activation. Thiscould argue for a structural separation of two independentfunctional sites in the IL-1 protein. Interestingly, however,significant differences exist among those amino acid residuesof IL-lae, IL-lI1 and ll-lRa that probably provide the contactpoints in each receptor-ligand complex. Accordingly, anapparently unaltered receptor affinity of an engineeredantagonist might also result from multiple compensatingeffects and it does not necessarily indicate a structuralindependence of the two ligand domains that bind or activatethe receptor, respectively.
Especially for IL-la variant D141G pure agonist or partialagonist activities were established in various cellularresponses (Yamayoshi et al., 1990), which resemble ourresults with human IL-4 variant Y124D. Considering thefundamental structural differences between IL-1 and IL-4,as well as between their respective receptors, it is interestingto speculate that the occurrence and the straight forwardidentification of high affinity antagonists in both systems iscorrelated with a single chain composition of both thereceptors.Other more complex receptor systems may put more
stringent constraints on structural modifications of the ligand.In the multichain IL-2 receptor system the intrinsically highKD of 70 nM of the ,B subunit (the homologue of the IL-4
receptor) was found to decrease to 5 pM in the presence ofthe a subunit (see Ringheim et al., 1991). A decreased KDof 1.2 nM was observed in YT cells due to the presenceof a still poorly defined 'y subunit. Interestingly, IL-2 variantsaffected in a subunit binding showed an apparently unalteredEC50 in the biological assays (Weigel et al., 1989;Zurawski and Zurawski, 1989; Ju et al., 1991). For theexplanation of this puzzling behaviour a model was recentlyproposed (Grant et al., 1992) according to which the asubunit contributes several functions to IL-2-mediatedsignalling through the high affinity IL-2 receptor system.Interestingly, the partial agonist and antagonist variantsaffected at position Glnl41 of murine IL-2 (Zurawski et al.,1990) were described to be deficient in -y subunit interactions.They inhibited efficiently, however, the IL-2 variantsdefective in a subunit binding. Possibly, the interaction ofthe 7y subunit and the fi subunit is critical for receptoractivation and any disturbance of this interaction will reduceboth receptor binding and receptor activation. This complexsubunit interplay in the IL-2 receptor system thereforecomplicates its analysis, but may also provide specialadvantages for the engineering of selective agonists (seeZurawski, 1991).
Increasing evidence supports the view that the formationof receptor homodimers or the forming of heterooligomersrepresents an important step in receptor transmembranesignalling (see Schlessinger, 1988; Williams, 1989; Ullrichand Schlessinger, 1990). This leads to the question how theligand triggers this aggregation. 'Allosteric receptoroligomerization' (Schlessinger, 1988), proposed as a modelfor tyrosine kinase receptors, involves two conformationsof the extracellular domain. Only the high affinity ligandbinding conformation forms oligomers. Bridging of tworeceptor proteins by a single ligand molecule representsanother possible mechanism for oligomerization (Ullrich andSchlessinger, 1990; Cunningham et al., 1991) supported bythe recent finding that growth hormone forms a 1:2 complexwith the isolated extracellular receptor domain. For the IL-4receptor, no evidence for the occurrence of dimers oroligomers has as yet been provided. A single class of IL-4receptor or IL4 binding protein has been identified so far,exhibiting one homogeneous dissociation constant of-100 pM. It remains to be established whether IL-4
receptor signalling involves the formation of a homodimeror whether the aggregation with other hitherto unknownproteins is necessary. The availability of antagonist IL-4variants will open up new ways to address the question ofwhich mechanisms operate during IL-4 receptor signalling.
Materials and methodsProduction of IL-4 mutant proteinsHuman IL-4 and mutant proteins were produced in Escherichia coli,subjected to a renaturation step and highly purified by CM-Sepharose 6Bchromatography followed by HPLC (Weigel et al., 1989; Kruse et al.,1991). By means of in vitro mutagenesis Tyrl24 (Kruse et al., 1991) hadbeen replaced by phenylalanine, histidine, asparagine, lysine, glycine oraspartic acid, respectively, to generate mutant proteins Y124F, Y124H,Y124N, Y124K, Y124G and Y124D. The mutated genes recloned into theexpression plasmid were sequenced in both directions (373A DNASequencer, Applied Biosystems) to confirm the mutation. Proteinconcentration was determined by measuring absorbance at 280 nm. It wasassumed that 1 mg IL-4/ml (50% Acetonitril and 0.1% trifluoracetic acid)yields an absorbance of 1 at 280 nm (-0'% = 1). For proteinmeasurements of variants deficient in Try 124, the absorbance at 280 nmwas multiplied by a factor of 1.15.
3242

Human IL-4 antagonist mutant proteins
T-cell proliferation assay(see Yokota et al., 1986; Solari et al., 1989) Peripheral blood mononuclearcells obtained from healthy donors were purified by Ficoll-Hypaquecentrifugation (Pharmacia) and stored in aliquots at -80°C. The thawedcells were cultured for 7 days with 9 Ag/ml phytohemagglutinine (PHA;HA-15, Wellcome) and consisted at this stage to a high percentage ofactivated T-cells (PHA-blasts). Cells (5 x 105/mIl) were incubated in0.2 ml aliquots with log 2 dilutions of the IL-4 protein for 3 days, beforethe amount of [3H]thymidine incorporated during the final 4 h wasdetermined. Maximal response was 6-23 x 103 c.p.m. incorporated[3H]thymidine over a background of 55-1400 c.p.m. The EC50 valueswere calculated from the known IL-4 concentration of a stock solution andthe dilution factor yielding half-maximal response.
Induction of the low affinity Fcc receptor (CD23) on B-cells(see Kikutani et al., 1986; DeFrance et al., 1987; Solari et al., 1989)Mononuclear cells were purified from the spleen of a donor that had nohaematological disease by centrifugation over Ficoll-Hypaque. B-cells wereisolated by rosetting of contaminating cells with ox erythrocytes coupledwith monoclonal antibodies anti-CD2, anti-CD4 and anti-CD8 PurifiedB-cells were positive by >95% for CD19. Tonsillar B-cells were preparedin the same way. B-cells were cultured at 106 cells/mil for 40 h with10 itg/ml soluble F(ab')2 fragments of affinity purified goat anti-human IgM(Jackson Laboratories-Dianova, Hamburg, Germany) plus log 3 dilutionsof IL-4 or mutant proteins. FACS analysis (FACScan, Becton-Dickinson)was done after double-staining with monoclonal antibodies against CDl9(Leul2-PE, Becton-Dickinson) and CD23 (lOB8-FITC, Dianova). Thenumber of CD23 positive cells and the relative mean fluorescence intensitywas calculated from a histogram obtained from the single cell analysis of5000 cells evaluated over a 4 log range. B-cells without stimulation wereCD23 positive by < 1% with a relative mean fluorescence intensity of < 10.After maximal stimulation 67 (62-72)% of the cells were CD23 positiveexhibiting a relative mean fluorescence intensity of 795 11. EC50 valueswere calculated from the known concentrations of the IL-4 stock solutionand the dilution factor leading to half-maximal response.
Receptor binding assays[125I]IL-4 labelled to a specific radioactivity of 0.22 ACi/pmol (Cabrillatet al., 1987) was incubated at a concentration of 1 nM with 107/ml RajiB-lymphoma cells or activated T-cells or activated tonsillar B-cells,respectively, in 0.2 ml plus the indicated concentrations of the competingIL-4 proteins. The [125I]IL-4 radioactivity bound during a 2 h incubationperiod at 4°C was determined. Dissociation constants, KD, were calculatedfrom relative ICse according to Ruffolo (1982). Measurements of [1251]IL-4binding to receptors on PHA-activated T-cells and activated tonsillar B-cells was performed as described above but using varying concentrations(10-1000 pM) of [125I]IL-4 in the absence and presence of 10 AMunlabelled IL-4, respectively.
Competitive inhibition assaysActivated T cells (PHA-blasts) were incubated with log 2 dilutions of IL-4(see above) plus a constant amount of mutant protein Y124D. B-cells (CD19positive cells) were incubated with log 3 dilutions of IL-4 (see above) ata constant concentration of mutant protein Y124D. The CD23 content ofthe cells was analysed by FACS and evaluated as described above. TheKi of variant Y124D was calculated from the apparent EC50 valuesmeasured in the presence of two different inhibitor concentrations [12] and[I'] according to the relation:
[121 - EC502/EC50' x [Ii]K = EC502/EC50' -
1 (Ruffolo, 1982)
Calculations of receptor occupancy and relative efficacyThe receptor occupancy y is the ratio between the concentrations of ligandedreceptor [RL] and total receptor [Re]. Making the most simple assumptionthat one ligand binds to one receptor with one homogeneous dissociationconstant KD then
% [RL] x 100 _ [L] x 100
[Rt] KD + [L]
It is further assumed that the dissociation constant KD of 10- 10 determinesthe IL4-dependent CD23 induction in B-cells. Then the receptor occupancyy for IL4 is 5% (0.4%) at the EC50 of 5 pM (0.4 pM) for the inductionof mean CD23 content (number of CD23 positive cells).The partial agonist Y124D probably saturates the receptor (y = 100%)
in B-cells to produce the maximal partial response, i.e. the induction of8% (45%) of the full mean CD23 content (number of CD23 positive cells).The relative efficacy (er) of two ligands is the ratio of the responses R'
and R2 that would be produced by these ligands at the same receptoroccupancy.
R' y2er= R2 Xy
Accordingly, for IL4 and variant Y124D the relative efficacy amountsto 50/8 x 100/5 = 125(50/45 x 100/0.4 = 275) during induction of meanCD23 content (number of CD23 positive cells).
AcknowledgementsWe thank H.U.Schairer, U.Schwulera and A.Duschl for help and discussion.This work was supported by the Deutsche Forschungsgemeinschaft and bythe Fonds der Chemie.
References
Abdel-Meguid,S.S., Shieh,H.-S., Smith,W.W., Dayringer,H.E.,Violand,B.N. and Bentle,L.A. (1987) Proc. Natl. Acad. Sci. USA, 84,6434-6437.
Bazan,J.F. (1990a) Proc. Natl. Acad. Sci. USA, 87, 6934-6938.Bazan,J.F. (1990b) Immunology Today, 11, 350-354.Black,J. (1989) Science, 245, 486-493.Brandhuber,B.J., Boone,T., Kenney,W.C. and McKay,D.B. (1987) Science,
238, 1707-1709.Cabrillat,J., Galizzi,J.-P., Djossou,O., Arai,N., Yokota,T., Arai,K. and
Banchereau,J. (1987) Biochern Biophys. Res. Commun., 149, 995-1001.Collins,L. et al. (1988) Proc. Natl. Acad. Sci. USA, 85, 7709-7713.Cook,W.J., Ealick,S.E., Reichert,P., Hammond,G.S., Le,H.V.,
Nagabhushan,T.L., Trotta,P.P. and Bugg,C.E. (1991) J. Mol. Biol., 218,675-678.
Cosman,D., Lyman,S.D., Idzerda,R.L., Beckman,M.P., Park,L.S.,Goodwin,R.G. and March,C.J. (1990) Trends Biochem. Sci., 15,265-270.
Cunningham,B.C. and Wells,J.A. (1991) Proc. Natl. Acad. Sci. USA, 88,3407-3411.
Cunningham,B.C., Ultsch,M., De Vos,A.M., Mulkerrin,M.G.,Clauser,K.R. and Wells,J.A. (1991) Science, 254, 821-825.
DeFrance,T. et al. (1987) J. Exp. Med., 165, 1459-1467.De Vos,A.M., Ultsch,M. and Kossiakoff,A.A. (1992) Science, 255,
306-312.Diederichs,K., Boone,T. and Karplus,P.A. (1991) Science, 254,
1779-1782.Eisenberg,S.P., Evans,R.J., Arend,W.P., Verderber,E., Brewer,M.T.,Hannum,C.H. and Thompson,R.C. (1990) Nature, 343, 341-346.
Finkelman,F.D. et al. (1990) Annu. Rev. Immunol., 8, 303-333.Galizzi,J.-P., Zuber,C.E., Harada,N., Gorman,D.M., Djossou,O.,
Kastelein,R., Banchereau,J., Howard,M. and Miyajima,A. (1990) Int.Immunol., 2, 669-675.
Garrone,P., Djossou,O., Galizzi,J.-P. and Banchereau,J. (1991) Eur. J.Immunol., 21, 1365-1369.
Gehrke,L., Jobling,S.A., Paik,L.S.K., McDonald,B., Rosenwasser,L.J.and Auron,P.E. (1990) J. Biol. Chem., 265, 5922-5925.
Grant,A.J., Roessler,E., Ju,G., Tsudo,M., Sugamura,K. andWaldmann,T.A. (1992) Proc. Natl. Acad. Sci. USA, 89, 2165-2169.
Hannum,C.H., Wilcox,C.J., Arend,W.P., Joslin,F.G., Dripps,D.J.,Heimdal,P.L., Armes,L.G., Sommer,A., Eisenberg,S.P. andThompson,R.C. (1990) Nature, 343, 336-340.
Harada,N., Castle,B.E., Gorman,D.M., Itoh,N., Schreurs,J., Barrett,R.L.,Howard,M. and Miyajima,A. (1990) Proc. Natl. Acad. Sci. USA, 87,857-861.
Idzerda,R.L. et al. (1990) J. Exp. Med., 171, 861-873.Ju,G. et al. (1991) Proc. Natl. Acad. Sci. USA, 88, 2658-2662.Kikutani,H. et al. (1986) Cell, 47, 657-665.Kruse,N., Lehmbecher,T. and Sebald,W. (1991) FEBS Letn., 286, 58-60.Kuhn,R., Rajewsky,K. and Muller,W. (1991) Science, 254, 707-710.Labriola-Tompkins,E. et al. (1991) Proc. Natl. Acad. Sci. USA, 88,
11182-11186.Liang,S.-M., Lee,N., Zoon,K.C., Manischewitz,J.F., Chollet,A., Liang,
C.-M. and Quinnan,G.V. (1988) J. Biol. Chem., 263, 4768-4772.Maliszewski,C.R., Sato,T.A., Vanden Bos,T., Waugh,S., Dower,S.K.,
3243

N.Kruse, H.-P.Tony and W.Sebald
Slack,J., Beckmann,M.P. and Grabstein,K.H. (1990) J. Immunol., 144,3028-3033.
Mosley,B. et al. (1989) Cell, 59, 335-348.Munson,P.J. (1983) Methods Enzymol., 92, 543-576.Park,L.S., Friend,D., Sassenfeld,H.M. and Urdal,D.L. (1987) J. Exp.
Med., 166, 476-488.Paul,W.E. and Ohara,J. (1987) Annu. Rev. Immunol., 5, 429-459.Paul,W.E. (1991) Blood, 77, 1859-1870.Redfield,C., Smith,L.J., Boyd,J., Lawrence,G.M.P., Edwards,R.G.,
Smith,R.A.G. and Dobson,C.M. (1991) Biochemistry, 30, 11029-11035.Ringheim,G.E., Freimark,B.D. and Robb,R.J. (1991) Lymnphokine Cytokine
Res., 10, 219-224.Ruffolo,R.R.Jr (1982) J. Auton. Pharniac., 2, 277-295.Sanderson,C.J., Campbell,H.D. and Young,I.G. (1988) Immunol. Rev.,
102, 29-50.Shanafelt,A.B., Miyajima,A., Kitamura,T. and Kastelein,R.A. (1991)EMBO J., 10, 4105-4112.
Solari,R. et al. (1989) Biochem. J., 262, 897-908.Schlessinger,J. (1988) Trends Biochem. Sci., 13, 443-447.Tepper,R.I., Levinson,D.A., Stanger,B.Z., Campos-Torres,J., Abbas,A.K.
and Leder,P. (1990) Cell, 62, 457-467.Ullrich,A. and Schlessinger,J. (1990) Cell, 61, 203-212.Urban,J.F.Jr, Katona,I.M., Paul,W.E. and Finkelman,F.D. (1991) Proc.
Natl. Acad. Sci. USA, 88, 5513-5517.Weigel,U., Meyer,M. and Sebald,W. (1989) Eur. J. Biochem., 180,295-300.
Williams,L.T. (1989) Science, 243, 1564-1570.Yamayoshi,M., Ohue,M., Kawashima,H., Kotani,H., lida,M., Kawata,S.
and Yamada,M. (1990) Lymphokine Res., 9, 405-413.Yokota,T., Otsuka,T., Mosmann,T., Banchereau,J., DeFrance,T.,
Blanchard,D., De Vries,J.E., Lee,F. and Arai,K.-I. (1986) Proc. Natl.Acad. Sci. USA, 83, 5894-5898.
Zurawski,S.M. and Zurawski,G. (1989) EMBO J., 8, 2583-2590.Zurawski,S.M., Imler,J.-L. and Zurawski,G. (1990) EMBO J., 9,
3899-3905.Zurawski,G. (1991) Trends Biotechnol., 9, 250-257.
Received on February 4, 1992; revised on May 19, 1992
3244


















